
二甲双胍对卵巢癌细胞的抗肿瘤作用
2024-02-29 01:47:06余慧李臻王佳
肿瘤防治研究 2024年1期
余慧,李臻,王佳
0 引言
卵巢癌是常见的妇科恶性肿瘤之一,其发病率排妇科恶性肿瘤第2位,死亡率居妇科恶性肿瘤首位[1]。临床上一线治疗方式是肿瘤细胞减灭术联合化疗,新兴治疗方式还有免疫治疗和靶向治疗[2]。大多数患者确诊时已达肿瘤晚期,发生远处转移并易复发,预后极差[2]。因此,亟需探索新的治疗方案以改善卵巢癌患者的预后。二甲双胍是一种常用的降糖药,基于人群的回顾性研究发现,二甲双胍在糖尿病患者的治疗中对癌症发生发展有一定程度的保护作用,可降低癌症发病风险和癌症相关死亡率[3-5]。有研究发现接受过二甲双胍/胰岛素治疗的患者卵巢癌发病率低于未接受过二甲双胍及胰岛素治疗的女性[6]。一项Meta分析显示使用二甲双胍的糖尿病患者其卵巢癌患病风险降低,因此推测二甲双胍具有潜在预防卵巢癌的作用[7]。但是二甲双胍的抗癌机制尚不明确。有研究表明,二甲双胍主要通过激活腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)通路而改变肿瘤细胞的代谢进而抑制卵巢癌细胞的增殖与转移[8]。CXCR4为趋化因子家族成员中的主要研究靶点,在多种恶性肿瘤的转移中也起重要作用。我们前期研究发现CXCR4在卵巢癌中高表达,参与卵巢癌的侵袭转移,并调控Wnt/β-catenin通路[9]。因此本研究拟探讨二甲双胍对卵巢癌细胞的体外抗肿瘤作用,及其是否对AMPK、CXCR4、Wnt/β-catenin通路产生影响,从而为二甲双胍应用于卵巢癌的临床治疗提供基础理论依据。
1 材料与方法
1.1 实验材料
人卵巢癌细胞株SKOV3、A2780(中国科学院细胞库,上海),CAOV3(吉凯基因,上海),DMEM高糖培养基、RPMI1640培养基(美国Gibco公司),胎牛血清(Gemini,澳达利亚)。二甲双胍购自美国MCE公司,溶于无菌PBS中,储存液浓度为500 mmol/L。MTT购自美国Sigma公司,Transwell小室购自美国Millipore公司,BCA蛋白定量试剂盒购自美国赛默飞公司。兔抗mTOR(#2983)、p-mTOR(#5536)、AMPK(#5831)、p-AMPK(#2535)均购自美国Cell Signaling Technology公司,兔抗CXCR4(ab181020)、Dvl3(ab76081)、β-catenin(ab32572)均购自美国Abcam公司。
1.2 细胞培养
SKOV3和CAOV3使用含10%胎牛血清的DMEM培养基,A2780细胞使用含10%胎牛血清的RPMI1640培养基,置于37℃、5%CO2培养箱,2~3天换液,待细胞融合至80%~90%,用0.25%胰酶消化传代。
1.3 MTT实验
将200 μl细胞悬液种于96孔板(5×103个/孔),过夜细胞贴壁后,将不同浓度的二甲双胍溶液(5、10、15、20、25、35、40、50和60 mmol/L)加入每孔中,同时正常培养基设为对照组,每组5个复孔。药物孵育24、48和72 h后取出96孔板,加入MTT溶液(终浓度0.5 mg/ml)后置于培养箱孵育3 h,吸净孔中溶液,加入200 μl DMSO,振荡器振荡10 min。酶标仪490 nm波长下测吸光度。细胞相对存活率=(OD实验组-OD空白组)/(OD对照组-OD空白组)。
1.4 平板克隆实验
将细胞悬液以1×103个/孔接种至6孔板,过夜细胞贴壁后,加入不同浓度二甲双胍(5、10 mmol/L),每3天换液,继续培养1~2周,待肉眼可见明显细胞集落后,PBS清洗3次,4%多聚甲醛固定15 min,0.1%结晶紫染色15 min,PBS洗3次,室温干燥后拍照,统计集落形成个数。实验重复3次。
1.5 流式细胞仪检测细胞周期
药物处理24 h后,用无EDTA胰酶消化细胞,收集细胞,1 500 r/min离心5 min,预冷PBS洗3次,70%乙醇4℃固定细胞过夜,PBS洗2次,加入RnaseA(50 μg/ml)孵育15 min,加入PI(50 μg/ml)避光孵育15 min,PBS重悬上流式细胞仪(美国BD公司)。
1.6 流式细胞仪检测细胞凋亡
药物处理24 h后,收集细胞中的培养基,用不含EDTA的胰酶将细胞消化下来,中止消化,收集细胞,1 500 r/min离心5 min,PBS洗3次,加入FITC Annexin V和PI,轻轻混匀,避光孵育15 min,同时设置空白对照组。再加入1×缓冲液,重悬后上流式细胞仪。
1.7 划痕实验
将细胞悬液种于6孔板,待细胞融合至100%,用200 μl枪头在单层细胞中垂直划过,形成划痕,PBS洗去漂浮细胞,加入含有不同浓度二甲双胍(5、10 mmol/L)的无血清培养基,间隔12 h拍照观察划痕愈合情况,直至划痕完全愈合。用Image J计算划痕愈合面积进行对比。
1.8 Transwell实验
下室加入500 μl含10%FBS的培养基,轻轻放入小室,将200 μl无血清培养基重悬的细胞悬液(5×104个)加入小室内,培养箱培养24 h后取出,4%多聚甲醛固定15 min,0.1%结晶紫染色,PBS清洗,用棉签小心擦拭小室内表面的细胞,室温晾干后于显微镜下拍照,计数。
1.9 Western blot实验
药物处理48 h后,冰化PBS清洗3次,冰上用含蛋白酶和磷酸酶抑制剂的裂解液裂解细胞15 min,4℃、12 000 r/min离心15 min,收集上清液,用BCA试剂盒进行蛋白定量,其余上清中加入上样缓冲液沸水煮10 min使蛋白变性。将20 μg蛋白在凝胶电泳系统中分离,再转移到NC膜上,牛奶室温封闭4 h,根据分子marker裁剪膜,分别置于一抗中孵育过夜。第二天用TBST洗膜3次后,室温孵育二抗1 h,再次洗膜3次,用ECL化学免疫发光液浸染条带置于Bio-Rad成像系统显影。
1.10 qRT-PCR实验
药物处理24 h后,按照FAST 200试剂盒(上海飞捷生物技术有限公司)的说明书提取细胞RNA,并用酶标仪在260 nm下定量RNA浓度。用Prime Script RT-PCR试剂盒(日本TaKaRa公司)将RNA反转录为cDNA后,与CXCR4引物(Forward Primer: ACTACACCGAGGAAATGGGCT,Reverse Primer: CCCACAATGCCAGTTAAGAAGA)和SYBR-Green PCR Master Mix(日本TaKaRa公司)进行扩增,利用CFX96 Real-time PCR系统(美国Bio-Rad公司)对CXCR4的表达进行检测。基因的表达量用2-ΔΔCT法计算,并以对照组进行标化。
1.11 统计学方法
组间比较使用t检验和One-way ANOVA,使用GraphPad Prism 8进行作图及统计学分析。P<0.05为差异有统计学意义。
2 结果
2.1 二甲双胍抑制卵巢癌细胞的增殖
M T T 实验评估二甲双胍对卵巢癌细胞(A2780、CAOV3、SKOV3)的细胞毒性,结果显示,二甲双胍对卵巢癌细胞的增殖具有抑制作用,且呈剂量及时间相关性。A2780细胞24小时IC50为91.5 mmol/L,48小时IC50为16.36 mmol/L,见图1A;CAOV3细胞24小时IC50为78.2 mmol/L,48小时IC50为36.65 mmol/L,见图1B;SKOV3细胞24小时IC50为78.25 mmol/L,48小时IC50为43.44 mmol/L,见图1C。同时,二甲双胍对单个卵巢癌细胞的克隆形成能力具有明显抑制作用,且呈剂量依赖关系,见图1D、1E。

图1 二甲双胍抑制卵巢癌细胞的增殖(A, B, C)及克隆形成能力(D, E)Figure 1 Metformin inhibited proliferation(A, B, C) and cloning ability(D, E) of ovarian cancer cells
2.2 二甲双胍导致卵巢癌细胞周期阻滞
二甲双胍导致卵巢癌细胞周期阻滞,细胞阻滞于G0/G1期,见图2A。同时通过Western blot检测周期蛋白Cyclin D1和CDK1的表达,与对照组相比,10、20和40 mmol/L的二甲双胍处理CAOV3和SKOV3细胞48小时后,Cyclin D1和CDK1表达水平降低,见图2B、C。

图2 二甲双胍导致卵巢癌细胞周期G0/G1期阻滞(A)并抑制周期蛋白(CyclinD1, CDK1)的表达(B, C)Figure 2 Metformin induced G0/G1 phase arrest of ovarian cancer cell cycle(A) and inhibited expression of cyclin (cyclin D1 and CDK1) (B, C)
2.3 二甲双胍诱导卵巢癌细胞凋亡
用Annexin V-FITC/PI对卵巢癌细胞进行染色后,通过流式细胞仪检测凋亡细胞。不同浓度的二甲双胍处理细胞24小时后,与对照组相比均可明显促进细胞凋亡,见图3。

图3 二甲双胍诱导卵巢癌细胞凋亡Figure 3 Metformin induced apoptosis of ovarian cancer cells
2.4 二甲双胍抑制卵巢癌细胞的迁移
选取增殖抑制率不超过10%的药物浓度进行划痕实验,观察二甲双胍对卵巢癌细胞迁移能力的影响。划痕处理24小时后,对照组的划痕区域被细胞覆盖(99.44%),空白划痕面积减小,二甲双胍组的划痕区域面积仍较大,说明经二甲双胍处理后的A2780细胞迁移能力减弱,且10 mmol/L的迁移抑制(24.26%,P=0.0051)较5 mmol/L(30.47%,P=0.0067)明显;划痕处理48小时后,对照组的划痕区域被细胞覆盖(69.59%),空白划痕区域面积减小,而二甲双胍处理的细胞划痕创面仍较大,说明经二甲双胍处理后的SKOV3细胞迁移能力减弱(P=0.0172),见图4A。

图4 二甲双胍抑制卵巢癌细胞迁移Figure 4 Metformin inhibited migration of ovarian cancer cells
通过Transwell实验也观察到同样的现象:与对照组相比,二甲双胍处理24小时后,穿过Transwell小室的卵巢癌细胞较少,说明二甲双胍明显抑制卵巢癌细胞的迁移能力(P<0.01),见图4B、4C。
2.5 二甲双胍抑制卵巢癌细胞增殖、迁移的机制
Western blot结果证明二甲双胍可以激活AMPK的磷酸化,抑制mTOR的磷酸化,抑制Dvl3和β-catenin及CXCR4的表达,见图5。
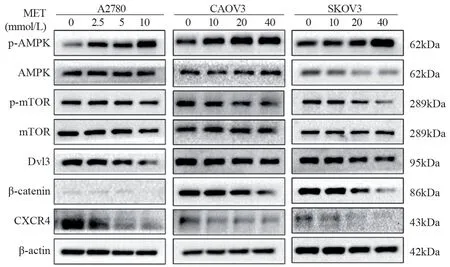
图5 二甲双胍对卵巢癌细胞p-AMPK/AMPK、p-mTOR/ mTOR、Dvl3、β-catenin及CXCR4蛋白表达水平的影响Figure 5 Effect of metformin on expression levels of p-AMPK/AMPK,p-mTOR/mTOR, Dvl3, β-catenin, and CXCR4 in ovarian cancer cells
二甲双胍处理CAOV3细胞24小时后,提取RNA进行RT-qPCR检测CXCR4 mRNA的变化,发现在转录水平二甲双胍抑制CXCR4的表达(P=0.0296),见图6。

图6 二甲双胍对CAOV3细胞CXCR4 mRNA表达水平的影响Figure 6 Effect of metformin on expression level of CXCR4 mRNA in CAOV3 cells
3 讨论
在体外细胞培养模型中,二甲双胍可以抑制多种肿瘤细胞的增殖,包括乳腺癌、前列腺癌、子宫内膜癌、宫颈癌[10-13]。既往研究表明,二甲双胍通过直接作用和间接作用发挥抗癌潜能:通过抑制线粒体呼吸链复合物Ⅰ降低肿瘤细胞的能量消耗直接抑制肿瘤生长,也可以通过降低空腹血浆胰岛素水平并增加骨骼肌葡萄糖摄取来减弱胰岛素抵抗,抑制肿瘤细胞的有丝分裂和生长而发挥间接抗癌作用[14]。这些抗癌作用都是通过细胞能量监测器AMPK介导的,AMPK的活化是二甲双胍发挥抗癌作用的主要效应分子[15]。AMPK依赖性抑制mTOR信号对肿瘤细胞的生长和增殖至关重要,是目前双胍类药物研究最多的抗肿瘤机制[15]。有研究发现,二甲双胍通过抑制CyclinD1而抑制前列腺癌的细胞增殖[11]。二甲双胍还可以通过卵巢癌细胞的AMPK/GSK3β轴诱导CyclinD1降解而导致细胞周期发生G1期阻滞[16],通过促进Parkin诱导的P53泛素化而抑制卵巢癌细胞生长[17]。
本研究发现二甲双胍可以有效抑制卵巢癌A2780、CAOV3、SKOV3的增殖、迁移,诱导细胞周期阻滞,促进细胞凋亡,在细胞水平发挥抑制肿瘤恶性行为的作用,还说明了二甲双胍可降低卵巢癌细胞中CXCR4蛋白及转录水平的表达。CXCR4属于趋化因子受体的CXC家族,可以被单一配体CXCL12特异性激活[18]。低氧条件下,癌细胞产生的CXCL12与内皮细胞表面的CXCR4相互作用可能会增强卵巢癌的血管生成[18]。Scotton等在卵巢癌细胞中检测了14种趋化因子受体,其中CXCR4是唯一表达于卵巢癌细胞的趋化因子受体[19]。我们的前期研究结果显示卵巢癌组织中CXCR4 mRNA的表达显著高于正常卵巢组织[9,20]。多项研究表明CXCR4在卵巢癌组织中高表达,且是上皮性卵巢癌患者的独立预后因素,CXCR4表达水平高预示着卵巢癌的预后较差[9,21-23]。CXCR4在卵巢癌的发生发展中具有促进肿瘤细胞迁移、侵袭及EMT转化的作用,有望成为治疗靶标[24-26]。因此,二甲双胍降低卵巢癌细胞中CXCR4的水平,可能是其发挥抗肿瘤作用的原因。有研究报道二甲双胍在杜氏肌营养不良症的动物模型中可以降低CXCL12和CXCR4的水平[27],还可以显著降低成纤维细胞中CXCR4的表达[28]。Dirat等[29]提出二甲双胍通过降低CXCR4的表达而抑制前列腺癌细胞的迁移。但二甲双胍单独作用时并不影响乳腺癌细胞中CXCR4蛋白的变化,仅在与熊果酸协同作用时才下调CXCR4的表达从而抑制乳腺癌细胞的侵袭和迁移[30]。本研究进一步拓展了二甲双胍在不同疾病模型中对CXCR4的影响形式。
总之,本研究提示二甲双胍可以抑制卵巢癌细胞的增殖,且呈时间-浓度依赖性。二甲双胍可以诱导细胞发生周期阻滞和细胞凋亡,并抑制细胞的迁移能力。本研究进行了二甲双胍发挥作用的初步机制探讨,这些作用可能是通过激活AMPK磷酸化、抑制mTOR磷酸化、抑制CXCR4的表达及Wnt/β-catenin通路来实现的,具体作用机制还需要进一步研究。
利益冲突声明:
所有作者均声明不存在利益冲突。
猜你喜欢
皮肤病与性病(2021年3期)2021-07-30 08:08:48
家庭医学(下半月)(2020年2期)2020-05-11 02:07:26
学生天地·小学低年级版(2017年12期)2018-04-16 03:24:06
中成药(2017年9期)2017-12-19 13:34:27
CHINESE JOURNAL OF AERONAUTICS(2017年1期)2017-11-21 12:54:14
中成药(2017年5期)2017-06-13 13:01:12
安徽医科大学学报(2016年12期)2017-01-15 14:21:54
初中生学习·高(2016年10期)2016-05-30 22:58:34
医学研究杂志(2015年4期)2015-06-10 06:42:43
物理实验(2015年10期)2015-02-28 17:36:58
