
伊犁河谷山地草甸群落特征和植物多样性对封育年限的响应
2024-02-28 03:29:10马晓林陈路瑶巴雅尔塔
新疆师范大学学报(自然科学版) 2024年1期
陈 茂,马晓林,陈路瑶,巴雅尔塔
(伊犁师范大学 资源与生态研究所 生物科学与技术学院,新疆 伊宁 835000)
围栏封育能最大限度降低人类活动和家畜活动对草地生态系统的破坏,使草地生态系统在无外界干扰的情况下依靠自身缓和能力进行修复,达到可持续发展[1]。封育改变群落物种组成结构,随着封育年限延长,禾本科植物代替杂草植物成为优势种,这种变化与植物生理生态特征的适应性有关[2]。由于各地区的环境、气候和植被群落结构有很大不同,将对封育年限有不同的响应情况。对南非、英国、美国的山地草甸进行调查可知,随着封育年限延长,草地群落特征逐渐趋向稳定,长期封育的效果相比4 年以内的短期封育来说无太大的提高[3]。对青藏高原东北边缘地区高寒草甸封育响应的研究表明,封育使草地植被盖度、高度、出现的频度得到增加,随着封育时间的增加,多样性指数不升反降,3~4年为较佳封育年限[4-5]。内蒙古锡林郭勒盟南部半干旱草原在封育第14年植被盖度、密度和多样性最大[6]。这些研究在不同季节、不同地区、不同气候、不同地形、不同植被群落结构上进行,反映出草地群落对封育年限有不同的响应。
伊犁河谷天然草地总面积约3.42 万km2,占伊犁河谷总面积的61.42%,草本植物的种类多达235 种,是天山山脉地区物种最丰富的地区,伊犁河谷草地在经济功能、农业功能和生态功能上是新疆各草地类型中最高水平代表[7]。作为新疆最为重要的陆地生态系统之一,近年来,由于过度放牧,破坏草地群落生态系统结构与功能的稳定性,伊犁河谷草地发展不可持续[8]。那拉提山地草甸作为伊犁州退化草原的典型代表之一,主要以两种表现为主:一是有毒植物大量繁殖,家畜误食中毒;二是过度放牧导致高大优良牧草减少,小型密丛形植物增加,占据优质牧草的生长空间,造成牧草产草量下降[9],对伊犁草地资源的保护刻不容缓。本研究为了更好研究封育对伊犁河谷山地草甸群落的影响,收集不同封育年限下的群落特征参数,研究在自然放牧、封育4年、封育10年的条件下山地草甸群落特征和多样性的变化对封育年限的反应,为伊犁河谷草地保护的最佳封育年限的确定提供科学依据。
1 材料与方法
1.1 样地自然概况
本实验选择新疆维吾尔自治区伊犁哈萨克自治州新源县那拉提退化草地围栏封育站山地草甸类草原自然保护区(43°16'57″N,83°58'47″E,海拔1800~2100 m)作为样地。该区气候属温凉湿润、半湿润山地气候,年降水量600~800 mm,无霜期为80~110 d,年日照时数2400~2700 h,≥10℃积温低于1000 h.土壤类型为山地黑钙土、灰褐色森林土等,植被类型为禾草和杂类草,主要原生植被有鸭茅(Dactylisglomerata)、细叶早熟禾(Poapratensis)、巨序剪股颖(Agrostisgigantea)、牛至(Origanumvulgare)等。新源县那拉提镇自1989 年起建立山地草甸自然保护封育区,封育群草地至今尚未受到除草、灌溉、施肥等其他人为修复措施的干扰[10]。
1.2 实验设计
在实验区退化山地草甸封育群落中,按照封育时间将其划分为夏季放牧地群落、4 年封育群落、10 年封育群落,即3个区组。在各区组设置间隔≥30 m的3个20 m×20 m采样单元(cell),以钢钉固定采样区边界,每个采样单元设置6个1 m×1m 样方,即一个区组18个样方,共设置54个1 m×1 m 样方。在样方内选取相同4株植物并用直尺测量株高,随后计算其平均值作为样方的高度,计算样方内所有相同植物类型总数量的密度,盖度采取目测估计法,按照功能群分为禾草、杂草、豆科,并收集地上生物量的样品一并置入65℃恒温烘干箱烘干12 h后称重。
1.3 数据分析
按物种调查的原始数据统计各个样地相对密度、相对盖度、相对频度,对每个样方计算各个物种的重要值。
选取以下指数进行多样性分析
Shannon-Wiener多样性指数
Simpson多样性指数
Margalef丰富度指数
Pielou均匀度指数
式中,S为物种数,N为全部种个数,Ni为种i的个体数,,使用Excel2016 和SPSS25 运用单因素方差分析和最小显著差法对不同封育年限下生物群落各指标进行比较,使用Origin2021 进行图行绘制,运用Python3.8的seaborn包绘制显著性热力图。
2 结果分析
2.1 封育年限对群落特征的影响
2.1.1 高度、盖度、密度分析
图1给出那拉提山地草甸自然放牧条件和不同封育年限下植被密度、盖度和高度。由图1可知,植被的密度呈现先升高后降低的趋势,从自然放牧的257株/m²增加到封育4年290株/m²,封育10年降低到184株/m².单因素方差分析与最小显著差法分析检验表明自然放牧和封育10 年植物的密度存在显著差异(P<0.05),自然放牧和封育4 年差异不显著(P>0.05),封育4 年与封育10 年存在显著差异(P<0.05)。整体上看,随着封育年限的增加,植物盖度呈现先增加后降低的变化趋势,在自然放牧、封育4年、封育10年分别达到41%、174%、158%,分析检验表明自然放牧与封育4年存在显著差异(P<0.05),封育4年和封育10年差异不显著(P>0.05)。高度随着封育时间的增加而增加,自然放牧到封育4 年和10 年,从5.7cm 增加到37.4 cm 和58 cm,自然放牧和封育4年存在显著差异(P<0.05),封育4年和10年存在显著差异(P<0.05)。
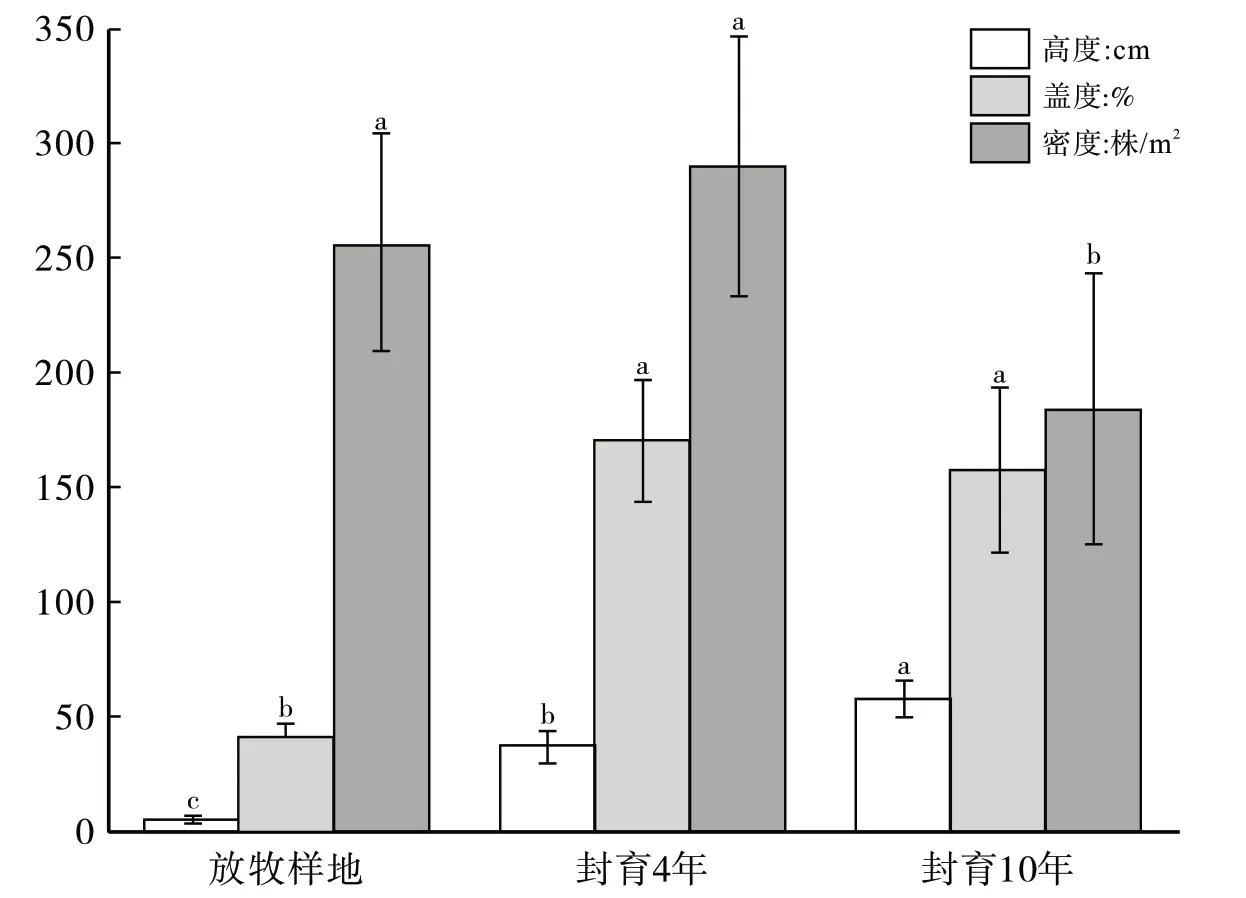
图1 不同封育年限植被密度和盖度分布状况
2.1.2 植物群落频度分析
频度作为植物群落的重要特征之一,能表示植物群种类的均匀程度(图2)。根据Raunkiaer制定的频度分析表,将出现的植物种类频度划分为5个等级,频度1%~20%为A 级,21%~40%为B级,41%~60%为C级,61%~80%为D 级,81%~100%为E 级。Raunkiaer 频度定律理想情况下物种数为A>B>C>D<E,因为罕见种在群落中总是较多,E级是群落中的优势种和建群种,一般情况下,E级物种越多群落越均匀[11]。由图2可知,封育4年后E级物种数大于自然放牧和封育10年物种数,说明封育4年群落的均匀度大于封育10年。
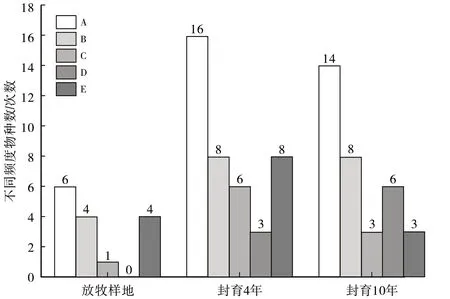
图2 不同封育年限频度分析图
2.1.3 封育对植物功能群生物量的影响
山地草甸植被群落生物量随着封育年限的延长而增加,植被群落地上生物量呈现增加的趋势(表1),依次为90.3 g/m²、270.6 g/m²、347.8 g/m².其中禾草的增加最为明显,从自然放牧的58.5 g/m²增加到封育10年的218.0 g/m²,杂草先从自然放牧的26.1 g/m²增加到175.0 g/m²后降低到118.5 g/m².通过方差分析检验发现,总生物量自然放牧与封育4 年存在显著差异(P<0.05),封育4 年与封育10 年差异显著(P<0.05),禾草和杂草比例的变化说明禾草类在随着封育年限的延长有较强的竞争力及恢复能力,而杂草类相反。
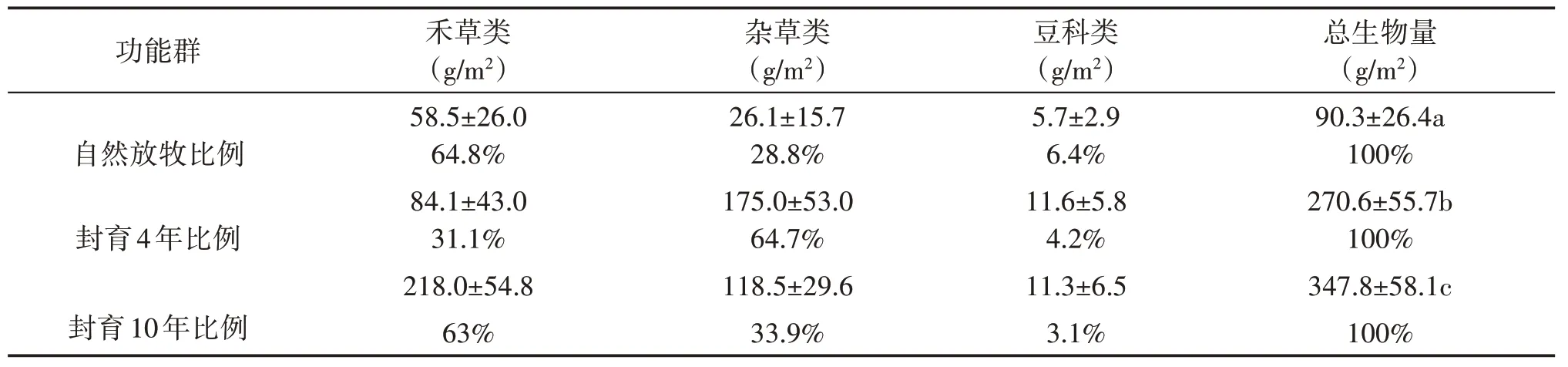
表1 不同封育年限生物量
2.1.4 群落种类组成分析
调查植物群落种类组成(表2),自然放牧条件下山地草甸植物群落种有15 种,分别属于7 个科,其中禾本科和毛茛科各4种,蔷薇科3种,牻牛儿苗科、菊科、车前科、卫矛科均为1种。封育4年后植物群落由41种植物组成,分别属于20个科,其中禾本科和菊科6种,蔷薇科5种,毛茛科4种,唇形科、豆科、伞形科、车前科各2种、牻牛儿苗科、茜草科、列当科、大戟科、杜鹃花科、百合科、紫草科、蓼科、卫矛科、十字花科、沙草科、忍冬科各1种。封育10年后植物群落由34种植物组成,分别属于15个科,其中禾本科7种,毛茛科和菊科各4种,豆科3种,蔷薇科5种,唇形科、伞形科2种,牻牛儿苗科、茜草科、列当科、大戟科、杜鹃花科、百合科、紫草科、蓼科各1种。从表2的重要值和物种名得到自然放牧和不同封育年限物种组成和优势种发生明显变化。自然放牧下建群种为细叶早熟禾(Poaangustifolia),优势种为白三叶(Trifoliumrepens)、天山羽衣草(Alchemilla tianschanica)、山地蒲公英(Taraxacumpseudoalpinum),封育4年下建群种为细叶早熟禾(Poaangustifolia),优势种为蓍(Achilleamillifolium)、天山羽衣草(Alchemillatianschanica)、无芒雀麦(Bromusjaponicus);封育10年下建群种鸭茅(Dactylisglomerata),优势种为箭头唐松草(Thalictrumsimplex)、虉草(Phalarisarundinaceae)、细叶早熟禾(Poaangustifolia)。封育4 年后物种比自然放牧增加26 种,封育10 年后物种比自然放牧增加19种,封育10年比封育4年物种数减少7种。自然放牧与封育4年下建群种均为细叶早熟禾(Poaangustifolia),自然放牧与封育4年相比,优势种只有天山羽衣草(Alchemillatianschanica),其余发生改变;自然放牧与封育10 年相比建群种和优势种都发生改变。优势种从自然放牧下的一种禾本科增加到封育4 年和10 年下两种禾本科。随着封育年限增加禾本科和豆科优良牧草的物种数明显增加。封育可能影响到植物物种重要值变化,有些物种的重要值明显增加,如箭头唐松草(Thalictrumsimplex)、虉草(Phalarisarundinaceae);有些物种重要值明显减少甚至消失,如白三叶(Trifoliumrepens)。虉草(Phalarisarundinaceae)生长需要占据大量水肥,竞争能力强,虉草(Phalarisarundinaceae)自然放牧下重要值为0,封育4 年、10 年下重要值分别为1.54、10.35,由群落不存在物种到伴生种、亚优势种和一些小型杂草的消失以及重要值降低,表明随着封育年限的延长重要值发生变化,草地向以高大植物为主的生态系统发展。
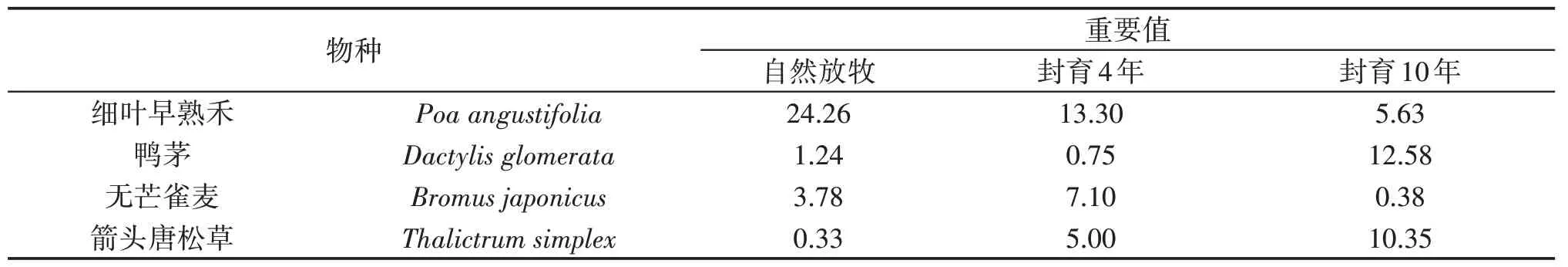
表2 不同封育年限群落物种组成及重要值
2.1.5 群落特征与封育年限的相关性
由热力分析图(图3)中的相关性可知植物群落特征与封育年限的相关性显著,高度和生物量为0.93 和0.86,与封育年限呈显著高度正相关;盖度为0.6,与封育年限呈显著中度正相关;物种数和密度为0.42 和-0.48,为显著弱相关并且密度为负相关。群落特征与封育年限的相关性:高度>生物量>盖度>物种数>密度,其中盖度与高度、生物量、物种数的相关性强,可能会导致封育时间超过10 年后高度和生物量的变化趋势与盖度相同。
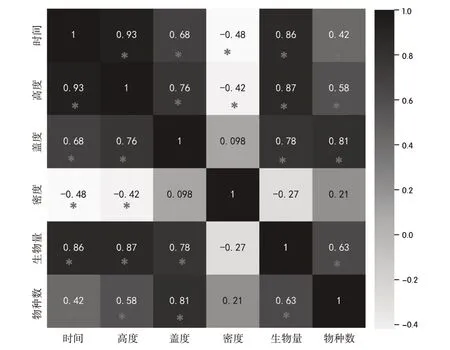
图3 封育年限与群落特征相关性热力分析图
2.1.6 山地草甸群落特征与封育年限的回归分析
封育能使退化的草地生态系统得到恢复,有效改善过度放牧导致的群落特征降低,提高群落特征具体数值。但是封育到一定时间后群落特征逐渐稳定,有些群落特征还出现下降的趋势,因此封育时间不是越长越好,针对植被的高度、密度、盖度、物种组成数、生物量这些群落特征的统计分析,使群落的不同特征在合适的封育年限达到最大值,根据各类群落特征参数构建回归方程。对封育年限与植被平均高度、盖度、物种数和生物量进行回归分析发现,植被平均高度随封育年限延长而增加,存在线性回归关系(图4),生物量(yb=24.68x+120.89,R²=0.837)表明随封育年限延长,至少在封育10 年内平均高度随着时间增加呈直线关系。但从群落盖度、生物量、密度和物种种数组成来看,随着封育年限延长,这些特征参数呈一元二次曲线关系,对这些特征参数构建函数,计算出最大值就可以得到最合适的封育年限,对植物群落的高度(yh=-0.47x²+9.16x+5.62,R²=0.92)、密度(yd=-2.58x2+18.8x+257.556,R2=0.411)、盖度(yc=-3.59x²+47.67x+41,R2=0.85)和物种种类组成数(ys=-0.347x2+4.068x+6.389,R2=0.86)进行函数分析发现,植物群落的高度、密度、盖度,物种种类组成数符合二次方程。通过对二次方程求最大值得到x 为9.7、3.9、6.2、5.9 时为函数的最大值,综上所述通过群落特征构建得到那拉提山地草甸最佳封育年限为6~8年最为合适。

图4 封育年限与高度、密度、盖度、物种数之间的关系
2.2 封育对群落多样性的影响
从图5可以看出多样性指数和均匀度指数随着封育年限增加呈增加趋势,丰富度指数先增加后降低,均匀度指数在封育第10 年达到最大,Pielou 均匀指数为0.88,封育第4 年Shannon-Wiener 多样性指数、Simpson多样性指数和Margalef丰富度指数最大,分别为2.26、0.85、2.8.方差分析发现Shannon-Wiener多样性指数和Simpson 多样性指数在自然放牧与封育4 年下存在显著差异(P<0.05),封育4 年与封育10 年下差异不显著(P>0.05)。Margalef 丰富度指数表明在自然放牧与封育4 年下存在显著差异(P<0.05),在封育4 年与封育10年下存在显著差异(P<0.05)。Pielou均匀度指数在自然放牧与封育4年下存在显著差异(P<0.05),在封育4年与封育10年下差异不显著(P>0.05)。
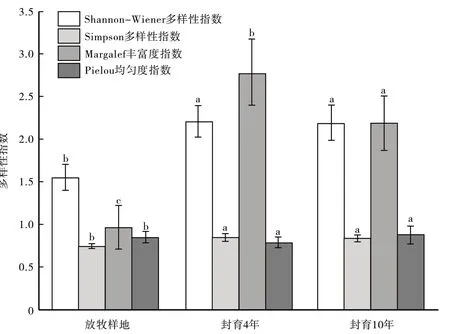
图5 自然放牧与不同封育年限物种多样性指数对比
3 讨论
新源县那拉提山地草甸保护区由于特殊的地理位置和气候,以及近年来旅游开发和传统畜牧业发展导致大部分草场出现不同程度的退化,生态环境破坏日趋严重。通过研究围栏封育年限对山地草甸草地植被群落结构的影响发现,不论是地上生物量、植被密度、盖度,还是物种组成及群落物种多样性,围栏封育均能够起到恢复作用,但对封育年限的讨论尤为激烈。
3.1 不同封育年限植物群落特征变化因素
不同的封育时间对植被群落特征的影响不同,在4年封育下物种数和盖度优于10年封育。黄东月等人研究发现短期封育能有效减少人为和家畜干扰,提高土壤的水分、有机质含量,环境差异性能为不同生态习性的物种提供生存空间,使物种数不断增加,封育也为植物的种子提供保护,使其免受家畜的采食,使植物能够顺利完成种子繁衍[12]。随着封育时间的增加,在西北地区干燥低温的环境下,植物的枯落物和腐质大量堆积,分解缓慢,不利于植物利用,导致土壤含水量降低,从而导致植物生存空间缩小,更加重视对光能的利用,而低矮植物对草地空间资源及光能的利用能力低,不利于低矮植物的生长[13]。随着封育时间的延长,竞争能力更强的高大禾草成为群落的主要物种,植物群落的高度随着时间延长而增加,这与本研究中物种高度封育10 年优于4 年的结论一致。一般情况下杂草类植被具有较宽的叶片,大多处在第二生长层,随封育时间延长,杂草类植物会因具有较厚的覆盖物而不能得到充足的光照,进而失去竞争力。因为叶片较窄,高度高,获得光照大于杂草类,有利于禾草生长,导致群落以细长叶片的禾草为主要外观,群落的盖度出现下降[14-15]。封育10年总生物量大于封育4年,这与张义等人的研究是一致的[16],余潇等人的研究表示由于该地处于半干旱气候和山地气候,掉落物的堆积,降水难以进入土壤,随封育年限延长,土壤含水量下降,禾草种子萌发形成幼苗很少,禾草主要依靠分蘖繁殖,导致禾草株丛大型化,禾草生物量显著提高[17]。该研究与本研究同处西北山地,气候较为一致,这可能是导致禾草在封育第10年大量增加的原因。封育10年后群落以禾草为优势种,在测量禾草密度时以丛为密度单位,而不是按株计算,禾草分蘖生长占据有限生存空间,导致杂草物种减少或消失,密度必然出现下降。
3.2 不同封育年限植物多样性变化因素
作为维持草地生态系统稳定和生产力基础之一的物种多样性[18],有关封育与物种多样性关系的研究较多,由于影响多样性的因子繁多并且复杂,研究结果也有较大差异。国内学者认为对退化山地草甸长期封育会减少物种多样性[19-20]。而国外对南澳大利亚低矮灌丛植被的研究表明长期封育对植物多样性有良好的影响[21]。Cardinale 认为不仅封育时间能影响植物多样性,植物组成,生长的环境以及草地退化程度都能影响植物多样性[22]。在封育措施保护下导致不同的封育时间土壤的水分、板结程度、微量元素的含量有所不同,罗亚勇等人的研究表明在土壤含水量、全N、全P 呈与多样性显著正相关[23]。刘晓琴的研究中封育时间延长后土壤的含水量下降[24]。闫成才等人的研究中封育13 年后土壤含水量、全N、速效P 都出现了下降[25]。可见封育时间与土壤理化性质并不呈正相关。本研究中封育后的多样性指数、丰富度指数并没有随着封育时间延长而增加,封育5年条件下的指数大于封育10年下,除了封育导致的土壤变化情况外,自然放牧状态下,由于家畜的采食,抑制早熟禾(Poaangustifolia)、鸭茅(Dactylisglomerata)、野豌豆(Viciacracca)等高营养适口的禾草和豆科的生长,不适口杂草类植物和毒类杂草在自然放牧状态下受到的抑制比较小,封育后消除了家畜的干扰,有利于植物的生长,体型较小的杂草在封育开始时竞争能力强,因此小型杂草大量繁殖,而且对土壤种子库起到保护作用,所以多样性指数和丰富度指数在封育早期达到最大[26-27]。随着群落内物种达到一定数量后,对资源的利用达到平衡,种间资源竞争平稳,群落趋于稳定[28],群落中的优势种的优势地位提升,优势种竞争力继续增强,群落中的劣势种或偶见种逐渐减少甚至消失,物种丰富度和多样性降低。可见植物多样性变化是多种因素导致的,其中既有土壤等外因也有植物生理特性,封育年限只是其中一项。
3.3 长期封育下有害草的繁殖
白喉乌头和唐松草作为有害毒草,封育10年相比,自然放牧和封育4年条件下不仅没有消失,而且在群落中优势度还出现上升,有毒杂草的繁殖也是当下伊犁河谷地区面临的严重问题之一[29-30]。唐松草(Thalictrum simplex)和白喉乌头(Aconitumleucostomum)在封育10年后重要值提高,白喉乌头(Aconitumleucostomum)等有毒草的繁殖出现在封育4年是因为长期的超载放牧导致草原毒害草增生,但封育10年后重要值下降后一度增加。一些学者认为封育能有效防止毒草的蔓延[31-32],从其他学者对白喉乌头(Aconitumleucostomum)和唐松草(Thalictrumsimplex)生长和繁殖特征的研究可以得出出现此类情况的原因:白喉乌头和唐松草的生活史类似于禾草,并且返青早、生长快。在土壤20~30cm 地方的种子库最大,能避免家畜的践踏,抗化学与生物除草剂的能力强,适合生长在干旱少雨地区,竞争能力强于蒲公英(Taraxacumpseudoalpinum)和羽衣草(Alchemillatianschanica)等小型草类[33-35],唐松草和白喉头高度和叶面积优于其他杂草类,获得的光能强于其他杂草,在长期封育后成为优势种杂草。加上封育10 年土壤的变化可能适合白喉乌头和唐松草的生长,并且文章研究样地未进行除草、喷洒农药等人为活动,封育防止了家畜对幼苗的践踏,将这几点结合起来,可能是导致长期封育下毒害杂草优势度不降反增的原因。
4 结论
文章通过对植被群落的特征和多样性具体值和方差分析植物群落的密度、盖度、物种数、Shannon-Wiener多样性指数、Simpson多样性指数和Margalef丰富度指数得到封育4年优于封育10年的效果,通过高度、生物量、Pielou均匀指数的分析得出封育10年优于封育4年。通过相关性热力图的分析发现,盖度与其他群落特征有中强度正相关并且根据群落特征构建的曲线方程中盖度在超过6年后出现下降,其他群落特征超过10年后可能也会出现相似情况。Pielou均匀指数和体现群落均匀性的植物种出现频度、Simpson多样性指数出现矛盾,这需要对更长的封育时间植物群落进行研究。通过重要值的变化,草地植物群落随着封育时间的延长从未封育的杂草为主要物种逐步向以禾草为主要物种的方向演进后趋于稳定,群落的多样性和丰富度出现下降。综上,长期封育不适合对那拉提山地草甸的保护,封育时间控制在6~9年最佳。
猜你喜欢
天津农林科技(2022年2期)2022-04-19 10:48:14
河南畜牧兽医(2022年3期)2022-04-13 09:12:58
草地学报(2021年7期)2021-08-03 13:37:10
绿色科技(2021年10期)2021-06-23 03:13:06
黑龙江工程学院学报(2020年5期)2020-10-21 05:37:10
水土保持研究(2019年6期)2019-10-19 03:33:32
草业学报(2019年2期)2019-02-25 01:57:16
南方农业·下旬(2017年11期)2018-01-09 14:37:50
中国饲料(2017年15期)2017-08-22 04:38:29
杂草学报(2015年2期)2016-01-04 14:57:54
