
基于九个微卫星标记的核桃举肢蛾地理种群的遗传多样性分析
2024-02-26 05:49:14王琦琦唐光辉
昆虫学报 2024年1期
王琦琦, 孙 艳, 唐光辉,*
(1. 西北农林科技大学林学院, 西部森林生物灾害治理国家林业和草原局重点实验室, 杨凌 712100;2. 中国科学院动物研究所, 动物进化与系统学院重点实验室, 北京 100101)
核桃举肢蛾Atrijuglanshetaohei又名核桃展足蛾、核桃黑,属鳞翅目(Lepidoptera)织蛾科(Oecophoridae)黑展足蛾属Atrijuglans,是对核桃果实危害最大的害虫(Wangetal., 2016; 王琦琦等, 2016)。其幼虫蛀食核桃青果,导致提前落果,核桃产量下降40%以上,严重影响核桃的品质和经济价值(田敏爵等, 2010)。每年,核桃举肢蛾危害在我国河北、河南、山东、山西、陕西、四川、贵州等核桃产区普遍发生,严重影响核桃果实的品质、产量和商业价值,给当地经济带来重大损失(田敏爵等, 2010; 武海斌等, 2020; 孟树标, 2022; 王永平等, 2022)。目前,有关核桃举肢蛾的研究主要集中于生物学特性(南小宁等, 2017; 刘桂湘等, 2018)、田间发生规律(武海斌等, 2020; 孟树标, 2022)及防治技术(王相宏等, 2015; 陈邦清等, 2021)等方面,对核桃举肢蛾种群遗传结构和种群扩张历史的研究却未见报道。
微卫星标记(microsatellite marker)又称简单序列重复(simple sequence repeat, SSR),因其具有高多态性、共显性、丰富性等特点,可作为DNA分子标记广泛应用于动植物的系统发生、遗传多样性和地理谱系等方面的研究。微卫星标记技术在农林业害虫的种群遗传结构和种群遗传研究中应用广泛,如瓜实蝇Bactroceracucuribitae、枣食芽象Scythropusyasumatsui、灰飞虱Laodelphaxstriatellus、天鹅绒豆毛虫Vespulapensylvanica和寄生蜂Anticarsiagemmatalis等(张亚楠等, 2018; 洪波等, 2019; 姜姗等, 2020; Huetal., 2023; Neivaetal., 2023; Rustetal., 2023)。
核桃种植已成为核桃产区的经济支柱产业,然而,许多产区皆有核桃举肢蛾危害,使当地经济蒙受巨大损失,为深入了解核桃举肢蛾的分布特点和扩散途径,本研究根据核桃举肢蛾转录组测序数据获取的大量微卫星位点,并对所选取的微卫星位点进行筛选,获得可用于核桃举肢蛾地理种群遗传多样性分析的9个微卫星分子标记,对8个省/市16个核桃举肢蛾地理种群的遗传多样性和遗传结构进行研究,分析其种群间的遗传分化和基因流,并对核桃举肢蛾种群扩散途径进行综合分析和探讨,为提高核桃举肢蛾的防控效率提供理论依据和参考资料,降低因核桃举肢蛾危害而造成的经济损失。
1 材料与方法
1.1 供试虫源和总DNA提取
于2016年8月上、中旬分别在我国北京、河北、河南、山东、陕西、甘肃、四川等8个省/市核桃举肢蛾的主要发生区采集核桃举肢蛾危害虫果,虫果采集距离保证50 m以上,且每个虫果中取出1头核桃举肢蛾幼虫作为供试样本,以保证其为来自不同父母本的后代个体,然后浸泡于无水乙醇中,带回西北农林科技大学林学院,-20 ℃保存备用,共计16个地理种群,319头个体,详细采集信息见表1。
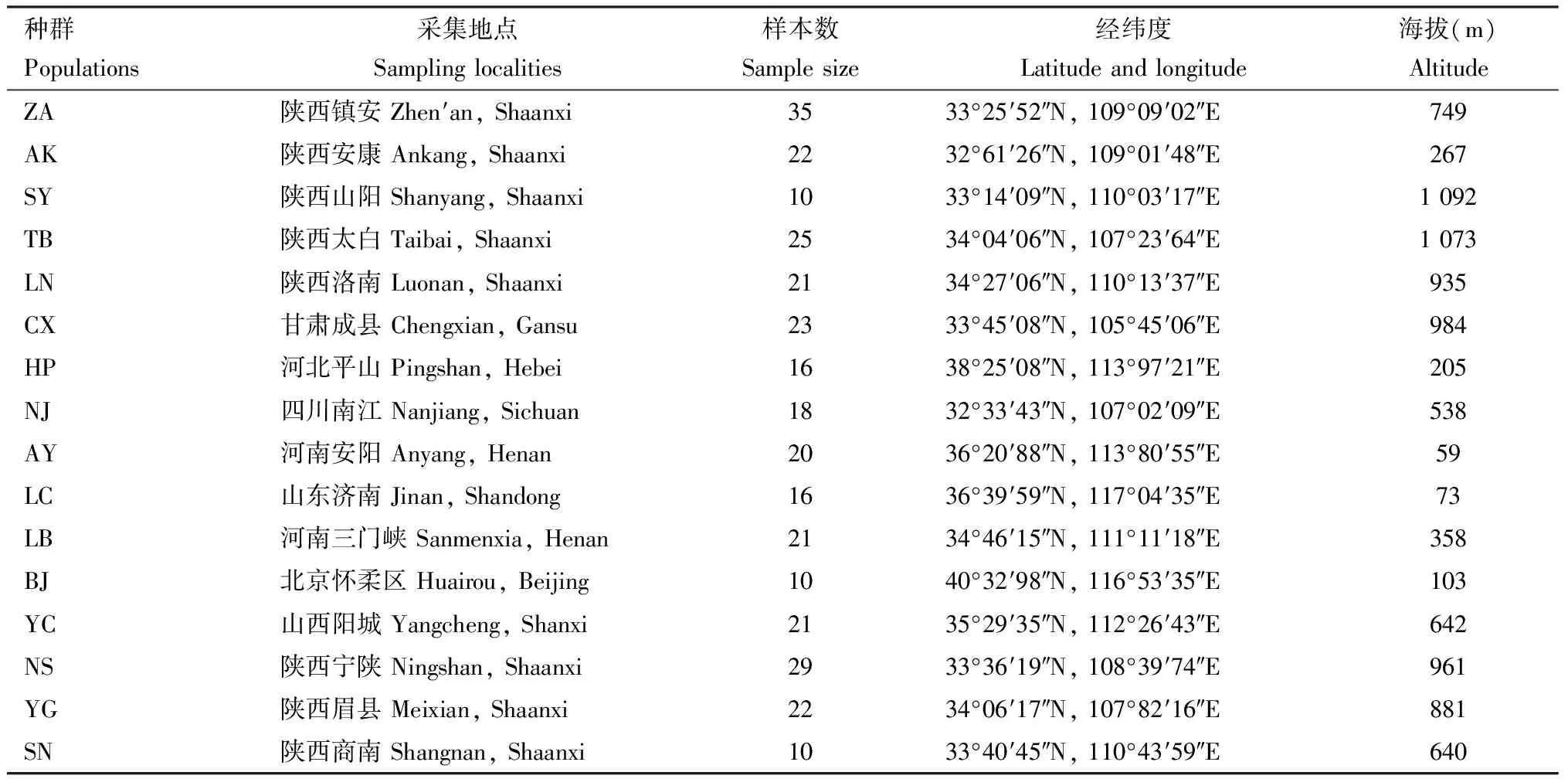
表1 核桃举肢蛾幼虫采集信息
将每个供试样本分别置于1.5 mL灭菌离心管中,使用血液/细胞/组织基因组DNA提取试剂盒[天根生化(北京)有限公司]进行总DNA提取。使用Q3000核酸测定仪(Quawell,美国)检测所提取的核桃举肢蛾基因组DNA的浓度和吸光值,检测合格的DNA样品保存于-20 ℃冰箱中。
1.2 RNA提取、cDNA文库的制备和测序
利用从陕西镇安采集的核桃举肢蛾危害虫果中获得的新鲜核桃举肢蛾幼虫,将15头幼虫的胸部组织置于1个1.5 mL离心管中,采用RNAisoPlus总RNA提取试剂盒(TaKaRa宝生物工程(大连)有限公司)提取总RNA并构建cDNA文库,由陕西博瑞德生物科技有限公司采用Illumina高通量测序平台(Hiseq 2000)进行测序(Lietal., 2020)。
1.3 微卫星位点的筛选和扩增
使用SSR软件MISA(MicroSAtellite)对核桃举肢蛾转录组45 696条unigene进行分析查找,共获得SSR位点2 586个。根据SSR重复单元长度和重复次数的不同(罗纯等, 2015),选出40个SSR位点,并使用Primer Premier 5.0软件设计引物并由上海生工生物工程技术服务有限公司合成。以1.1节提取的16个地理种群的核桃举肢蛾幼虫DNA为模板进行PCR扩增,PCR扩增反应体系(20 μL): 2×Taq PCR MasterMix 10 μL, 正反向引物(10 μmol/L)各1 μL, 模板DNA(50~100 ng/μL)1 μL, ddH2O补足至20 μL。PCR反应程序: 94 ℃ 5 min; 94 ℃ 30 s, 退火 30 s, 72 ℃ 30 s, 共35个循环;72 ℃ 5 min。PCR产物用1%凝胶电泳检测以判断引物的稳定性和特异性。然后经6%的非变性聚丙烯酰胺凝胶电泳(polyacrylamide gel electrophoresis, PAGE)对扩增片段的大小和多态性进行检测,最终筛选出具有高多态性且易于扩增的微卫星位点并对所有样本进行扩增,送上海生工生物工程技术服务有限公司经ABI3700自动测序仪(赛默飞, 美国)进行毛细管电泳(capillary electrophoresis, STR)检测,检测结果使用GeneMarker软件进行基因分型,并对位点数据进行人工校对。
1.4 数据分析
1.4.1数据整理与遗传参数统计:(1)数据整理:将基因分型数据按照种群和位点的顺序进行整理并储存在Excel中;(2)遗传参数统计:使用GenAlex version 6.5(Peakall and Smouse, 2005, 2012)和Popgene version 1.32(Yehetal., 1999)软件统计各种群和各位点的遗传参数,包括等位基因数(number of alleles,Na)、有效等位基因数(effective number of alleles,Ne)、观测杂合度(observed heterozygosity,Ho)、期望杂合度(expected heterozygosity,He)、非偏差期望杂合度(unbiased expected heterozygosity,uHe)、Shannon’s指数(Shannon’s index,I)、近交系数(inbreeding coefficient,FIS)和基因流(gene flow,Nm)等。使用PIC_CALC version 0.6软件(Nagyetal., 2012)计算多态信息含量(polymorphism information content,PIC)。
1.4.2遗传平衡检验和遗传距离-地理距离相关性分析:(1)使用GenePop version 3.4软件(Rousset, 2008)对核桃举肢蛾各种群和各位点进行哈迪-温伯格平衡(Hardy-Weinbergequilibrium, HWE)和连锁不平衡(linkage disequilibrium)的显著性分析,结果经过Bonferroni校正;(2)用Google Earth version 7.1.8软件测算各种群间的地理距离,并使用Excel分析种群间的遗传分化系数[FST/(1-FST)]与地理距离(km)自然对数间的相关性。
1.4.3种群遗传结构分析:(1)使用STRUCTURE version 2.3.4软件(Pritchardetal., 2000; Hubiszetal., 2009)根据贝叶斯聚类方法基于蒙特卡罗马尔可夫算法分析核桃举肢蛾的种群遗传结构。K值范围1-16,每个取值重复计算10次,利用Evanno的ΔK法确定最佳K值(Evannoetal., 2005; Earl and Vonholdt, 2011)。使用CLUMPP version 1.1.2软件(Jakobsson and Rosernberg, 2007)进行比对整合并使用Distruct软件(Rosenberg, 2004)绘图;(2)基于非空间模型和贝叶斯方法,使用BAPS(Bayesian analysis of population structure)软件(Coranderetal., 2008)将每个种群视为1个遗传整体进行种群遗传结构分析,设定K值取值范围1-16,各取值重复计算10次;(3)基于种群间的Nei氏遗传距离(Nei, 1978)构建遗传距离矩阵进行主坐标分析(principal coordinates analysis, PCoA)。
1.4.4分子变异分析(analysis of molecular variance, AMOVA):使用Arlequin version 3.52软件包(Excoffieretal., 2007)计算种群内和种群间的分子遗传变异以及各种群间的遗传分化系数(FST)和基因流(Nm), 1 000次随机重复抽样。其中,以‘Sum of squared size difference (RST)’代替‘Number of different alleles(FST)’以获取更多的位点信息。
2 结果
2.1 核桃举肢蛾SSR位点的筛选
根据转录组测序结果选取的40对微卫星引物中,通过1%凝胶电泳检测出可稳定扩增的特异性引物后,经6%的非变性聚丙烯酰胺凝胶电泳(PAGE)进行检测,共筛选出9个高多态性的SSR位点用于核桃举肢蛾遗传多样性分析,其位点编号分别为SSR05, SSR08, SSR12, SSR21, SSR23, SSR28, SSR32, SSR33和SSR38(表2)。
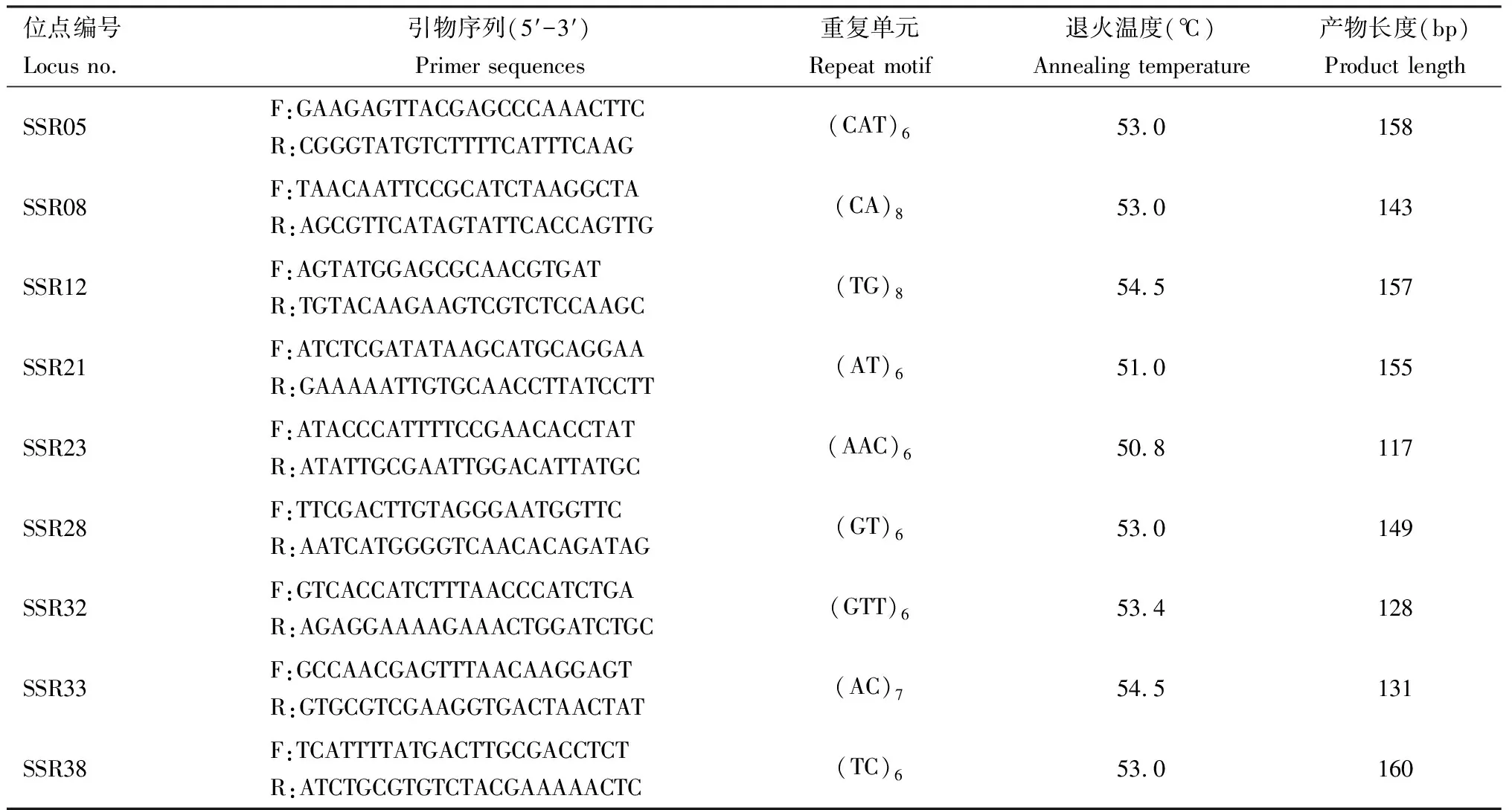
表2 核桃举肢蛾地理种群9个微卫星位点引物序列及位点特征
2.2 基于9个微卫星位点的核桃举肢蛾地理种群遗传多样性
在16个地理种群中共检测到46个等位基因(表3),各位点等位基因数(Na)在3~7之间,有效等位基因数(Ne)变化范围是1.019~2.286。其中,SSR05的Na和Ne值分别为4和1.019,在所有位点中均为最低,SSR28的Na最高为7,而Ne值最高的位点是SSR08(2.286)。Shannon’s指数(I)为0.063~0.931,平均值为0.557;观测杂合度(Ho)为0.019~0.627,平均值为0.268;期望杂合度(He)为0.018~0.509,除SSR05, SSR23和SSR32外其他位点的观测杂合度均低于期望杂合度,即杂合缺失。多态信息含量(PIC)为0.019~0.469。总体上,核桃举肢蛾各位点等位基因的多态性相对中等,可提供一定的遗传信息。
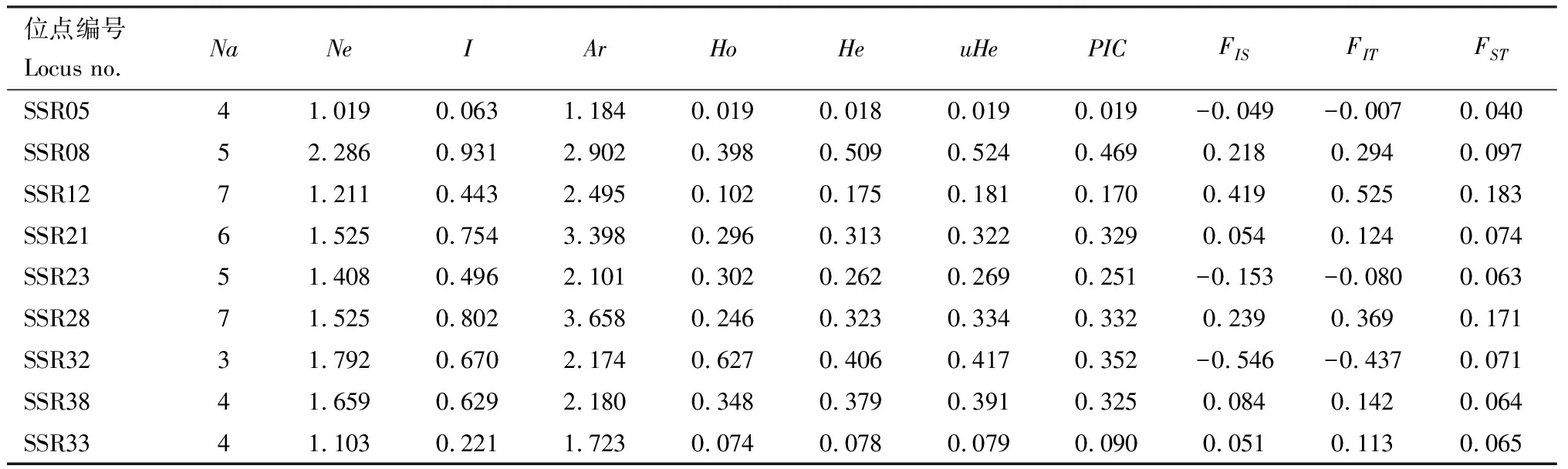
表3 核桃举肢蛾地理种群9个微卫星位点的遗传多样性
由表4可以看到,核桃举肢蛾16个地理种群的微卫星位点特征较为一致,各地理种群均存在一定程度的多态性。其中,山西阳城种群(YC)的等位基因数最高为3.000,其I(0.594)和北京怀柔区种群(BJ)的uHe(0.360)均高于其他种群的,表现出相对较丰富的种群多样性,而陕西山阳种群(SY)的多样性水平则较低,各个参数(Na,Ne,I,Ho,He和uHe)值都是最低的。此外,北京怀柔区种群(BJ)、河北平山种群(HP)、河南安阳种群(AY)、河南三门峡种群(LB)、山东济南种群(LC)、山西阳城种群(YC)、陕西洛南种群(LN)、陕西商南种群(SN)和陕西山阳种群(SY)的观测杂合度Ho皆小于期望杂合度He,且各种群的FIS范围在-0.153(NJ)~0.202(LB),说明群体内的杂合子不足,种群内近交机会更多,这可能与核桃举肢蛾本身的飞行能力不足所导致的活动范围受限有关。
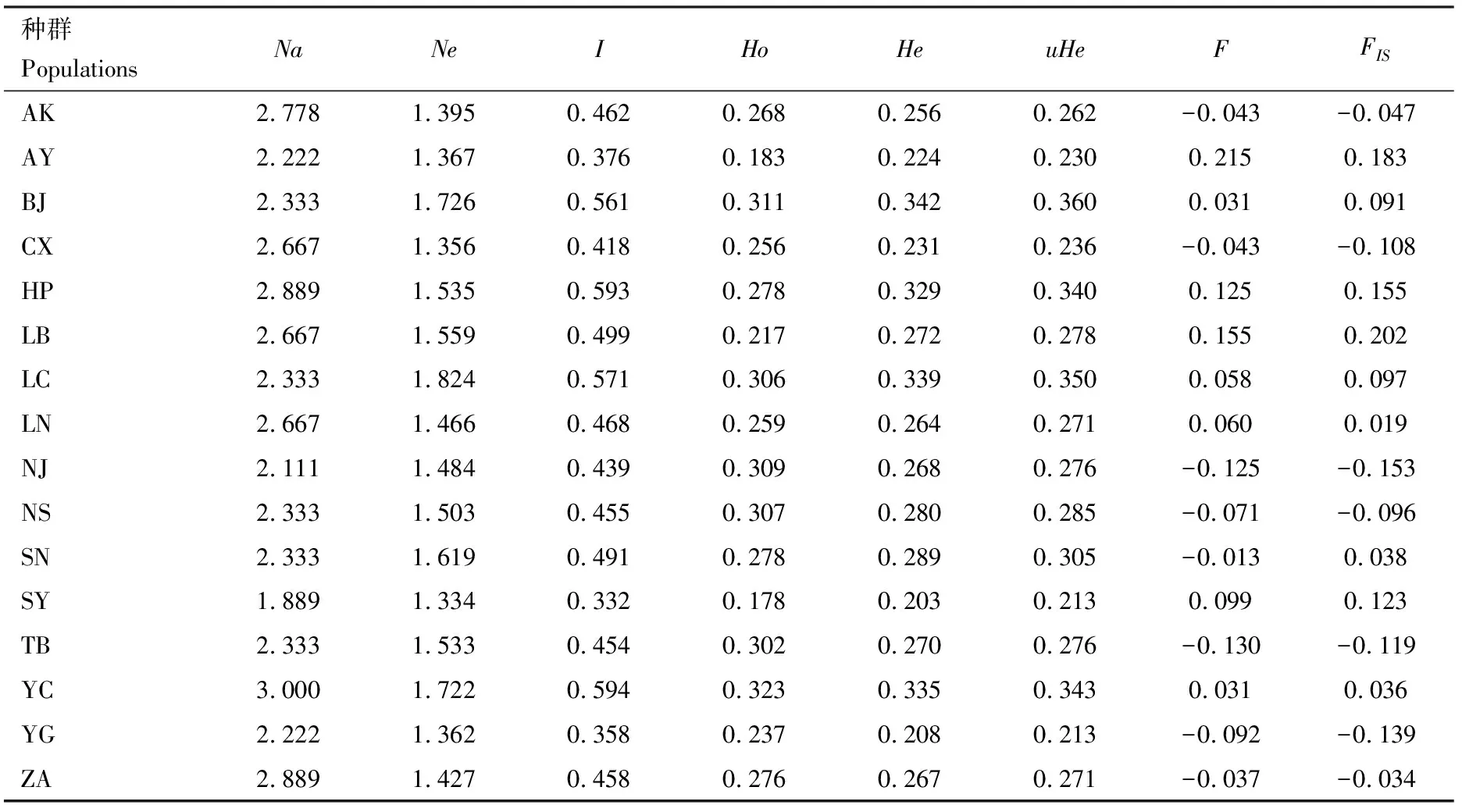
表4 基于9个微卫星位点的核桃举肢蛾16个地理种群的遗传多样性
2.3 核桃举肢蛾地理种群微卫星位点的哈迪-温伯格平衡和连锁不平衡
如表5所示,各地理种群在大多数位点均未偏离哈迪-温伯格平衡(P>0.05)。在位点SSR08上显著偏离哈迪-温伯格平衡(P<0.05)的种群有8个,其中,河南三门峡种群(LB)、山西阳城种群(YC)、陕西商南种群(SN)和陕西宁陕种群(NS)在该位点呈极显著偏离哈迪-温伯格平衡(P<0.01)。此外,河南安阳种群(AY)在位点SSR21也表现为极显著偏离哈迪-温伯格平衡(P<0.001)。
表5 核桃举肢蛾16个地理种群9个微卫星位点的哈迪-温柏格平衡
9个微卫星位点的基因连锁不平衡检测结果(表6)显示,除位点对SSR08和SSR12(P=0.022), SSR12和SSR21(P=0.039)以及SSR38和SSR33(P=0.003)外,其他成对位点间不存在连锁不平衡的情况,为完全独立遗传。

表6 核桃举肢蛾16个地理种群在各成对微卫星位点间的基因连锁不平衡
2.4 核桃举肢蛾地理种群遗传分化和基因流
16个地理种群的遗传分化(FST)及其基因流(Nm)计算结果(表7)表明,各地理种群间的遗传分化指数最高为0.142,表明各种群间的遗传分化处于中等或较低水平。山东济南种群(LC)与其他种群间遗传分化系数皆大于0.05,表明存在中等水平的遗传分化。陕西镇安种群(ZA)和陕西安康种群(AK)的种群间基因流最大为23.800,山东济南种群(LC)与陕西太白种群(TB)的种群间基因流最小,为1.514,但与陕西眉县种群(YG)、陕西商南种群(SN)、陕西宁陕种群(NS)和甘肃成县种群(CX)的种群间的基因流皆小于2。因此,LC与上述5个种群间存在较大的遗传分化,而其他种群间的遗传分化并不明显。

表7 基于9个微卫星位点的核桃举肢蛾16个地理种群间成对固定系数FST (下三角)和基因流Nm (上三角)
2.5 核桃举肢蛾地理种群遗传距离与地理距离间的相关性
以16个地区的核桃举肢蛾种群间的遗传距离和地理距离进行相关性分析,如图1所示,回归方程y=0.0209x-0.0707,R2=0.226,P<0.01,表明核桃举肢蛾种群遗传距离与地理距离间存在显著的相关性。
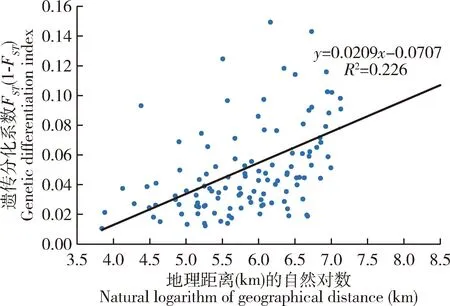
图1 基于9个微卫星位点的核桃举肢蛾不同地理种群间遗传分化系数与地理距离(km)的自然对数间的相关性
2.6 核桃举肢蛾地理种群遗传结构
使用STRUCTURE软件对16个核桃举肢蛾地理种群进行聚类分析(图2),当K为2时分组最佳(图2: A),即16个核桃举肢蛾地理种群分为2个分支:分支1由北京怀柔区种群(BJ)、河北平山种群(HP)、河南三门峡种群(LB)、山东济南种群(LC)、山西阳城种群(YC)、陕西商南种群(SN)和陕西山阳种群(SY)组成;分支2由河南安阳种群(AY)、陕西洛南种群(LN)、陕西眉县种群(YG)、陕西太白种群(TB)、陕西宁陕种群(NS)、陕西镇安种群(ZA)、陕西安康种群(AK)、甘肃成县种群(CX)和四川南江种群(NJ)组成(图2: B)。

图2 基于9个微卫星位点的核桃举肢蛾地理种群遗传结构
同时,使用BAPS软件分析核桃举肢蛾种群的遗传结构,结果如图2(C),山东济南种群(LC)个体分歧过大,不参与分组,其他15个地理种群分为两个分支,分支1由北京怀柔区种群(BJ)、河北平山种群(HP)、河南三门峡种群(LB)、山西阳城种群(YC)和陕西商南种群(SN)种群组成,分支2由河南安阳种群(AY)、陕西洛南种群(LN)、陕西眉县种群(YG)、陕西太白种群(TB)、陕西宁陕种群(NS)、陕西镇安种群(ZA)、陕西山阳种群(SY)、陕西安康种群(AK)、甘肃成县种群(CX)和四川南江种群(NJ)组成。
主坐标分析结果如图3所示,第1和第2坐标因子的贡献率分别为108.7%和19.6%;北京怀柔区种群(BJ)、河北平山种群(HP)、河南三门峡种群(LB)、山西阳城种群(YC)和陕西商南种群(SN)的核桃举肢蛾相距较近,占据第3和第4象限,说明其具有相似的主要成分;河南安阳种群(AY)、陕西洛南种群(LN)、陕西眉县种群(YG)、陕西太白种群(TB)、陕西宁陕种群(NS)、陕西镇安种群(ZA)、陕西安康种群(AK)、甘肃成县种群(CX)和四川南江种群(NJ)集中在第二象限;而山东济南种群(LC)和陕西山阳种群(SY)位于第一象限并与前两组相距较远,表明这两个种群与前两组差异较大,尤其是山东济南种群(LC)。

图3 基于9个微卫星位点的核桃举肢蛾16个地理种群基于Nei氏遗传距离矩阵的主坐标分析(PCoA)
2.7 AMOVA分子方差分析
基于STRUCTURE和BAPS分析结果,根据山东济南种群(LC)和陕西山阳种群(SY)分组的不同分别将16个地区的核桃举肢蛾种群分为2组(表8)或3组,并对种群的分子遗传变异进行比较分析。在不同分组条件下,各参数结果呈现一定程度的一致性,FST值都小于0.1,说明群体内个体间存在中等甚至很小的遗传分化。同时,种群内部的遗传变异占比都在90%以上(STRUCTURE中为95.42%,BAPS中分别为95.33%和92.71%),而种群间遗传变异则较小。此外,考虑到其他可能影响种群分布的因素,包括地理阻隔和气候差异等,进一步的分子变异分层分析结果可以看到,不同分组方式下的遗传分化FCT值在0.03941~0.06449之间,说明地理阻隔和气候差异不是影响核桃举肢种群遗传结构和地理分布格局的主要因素。

表8 基于9个微卫星位点的核桃举肢蛾16个地理种群的AMOVA分析
3 讨论
本研究根据核桃举肢蛾转录组数据进行微卫星引物的设计,使用非变性聚丙烯酰胺凝胶电泳和毛细管电泳筛选出9个多态性较高的微卫星位点,并对核桃举肢蛾的种群遗传多样性进行分析,同时对核桃举肢蛾的种群分布和种群扩散途径进行探讨,为鳞翅目昆虫的微卫星位点开发提供了参考资料,也为核桃举肢蛾的防治提供理论参考。
3.1 核桃举肢蛾的种群遗传多样性
在种群遗传多样性研究中,等位基因数、观测杂合度和期望杂合度等参数是衡量种群遗传多样性水平的重要指标,其值的大小可反映基因的丰富程度和研究对象对环境的适应能力(谢丽等, 2009)。利用9个微卫星标记对核桃举肢蛾16个地理种群的遗传多样性分析发现,平均每个标记的等位基因数在3~7个,所有位点的平均Ho为0.268,平均He为0.274(表3),表明各种群的遗传多样性较低。所有位点的平均PIC为0.260,除SSR05, SSR33和SSR12外,其余6个位点的PIC均大于0.25,为中度多态性位点,可以提供一定的多态信息对核桃举肢蛾的遗传多样性进行分析,这与我们前期基于线粒体基因和核基因的核桃举肢蛾种群遗传多样性分析结果(Wangetal., 2022)是一致的。16个种群的平均Ho和平均He分别为0.268和0.274,其中,北京怀柔区种群(BJ)、河北平山种群(HP)、河南安阳种群(AY)、河南三门峡种群(LB)、山东济南种群(LC)、山西阳城种群(YC)、陕西洛南种群(LN)、陕西商南种群(SN)和陕西山阳种群(SY)的平均期望杂合度高于平均观测杂合度(表4),且近交系数FIS为正,皆表明这些种群内有不同程度的近交。然而,陕西眉县种群(YG)、陕西太白种群(TB)、陕西宁陕种群(NS)、陕西镇安种群(ZA)、陕西安康种群(AK)、甘肃成县种群(CX)和四川南江种群(NJ)的种群平均Ho高于平均He且各种群的FIS为负,表明其近交程度很低,可能有大量外来个体补充到当地种群中,或种群经历了瓶颈效应或奠基者效应(Stoneetal., 2002),这可能与核桃举肢蛾起源单一,种群扩张历史较短,尚未积累到足够的遗传变异有关,另一个可能的原因则是经济林木害虫的种群扩散受人为因素影响更大。
3.2 核桃举肢蛾的遗传分化、基因流和遗传结构
群体间的遗传分化指数FST又称为固定系数,反映了种群间的遗传分化程度。FST越大,表明两种群间的差异越大,分化程度越高。本研究中,不同地区的核桃举肢蛾都具有中等程度的遗传分化,其遗传分化系数在0.010~0.142之间(表7)。遗传分化程度与种群间的基因流呈负相关,陕西镇安种群(ZA)和陕西安康种群(AK)之间的遗传分化最低,基因流最大。而山东济南种群(LC)与大多数群体间的遗传分化系数大于0.1,表明该种群较其他种群有一定程度的遗传分化。
综合STRUCTURE和BAPS聚类分析以及主坐标分析的结果发现,虽然不同分析结果存在一定的差异,但不同方法获得的核桃举肢蛾的种群遗传结构具有一定的共性,即将所有种群大致分为两组:分组1包括北京怀柔区种群(BJ)、河北平山种群(HP)、河南三门峡种群(LB)、山西阳城种群(YC)和陕西商南种群(SN),分组2包括河南安阳种群(AY)、陕西洛南种群(LN)、陕西眉县种群(YG)、陕西太白种群(TB)、陕西宁陕种群(NS)、陕西镇安种群(ZA)、陕西安康种群(AK)、甘肃成县种群(CX)和四川南江种群(NJ),但在山东济南种群(LC)和陕西山阳种群(SY)的归属问题上出现分歧(图2)。从主坐标分析图(图3)可以看到,相对于陕西山阳种群(SY),山东济南种群(LC)与前述两组间距离都是最远的,而在BAPS分析中,当将所有种群分为两组时,该种群被排除在外,表明其不属于任何一组,而若将所有种群分为3组时,山东济南种群则单独为一组。与此相异的是,在STRUCTURE分析中,最佳分组K取值为2时,山东济南种群(LC)归属于分组1。另一方面,BAPS和STRUCTURE分析在陕西山阳种群(SY)的分组上也存在分歧:在BAPS分析中,山阳种群属于分组2,而在STRUCTURE中K=2时又属于分组1,但在K=3时,陕西山阳种群(SY)又和分组1的大部分种群形成一个分组。因此,陕西山阳种群(SY)可能与分组2更相似,这与PCoA分析结果是一致的。综上所述,排除山东济南种群(LC),其他15个地区的核桃举肢蛾种群可分为两组,组1包括北京怀柔区种群(BJ)、河北平山种群(HP)、河南三门峡种群(LB)、山西阳城种群(YC)和陕西商南种群(SN)共5个种群,分组2则由河南安阳种群(AY)、陕西洛南种群(LN)、陕西眉县种群(YG)、陕西太白种群(TB)、陕西宁陕种群(NS)、陕西镇安种群(ZA)、陕西山阳种群(SY)、陕西安康种群(AK)、甘肃成县种群(CX)和四川南江种群(NJ)共10个种群组成。
3.3 影响核桃举肢蛾种群遗传结构的因素
已有许多研究报道,季风、气流和扩散能力等其他因素也可促进种群扩散并形成特定的种群遗传结构(Weietal., 2013; Xunetal., 2016)。例如夏季亚热带季风有助于许多小型有翅昆虫通过气流由东南向西北扩散。然而,核桃举肢蛾一年1代以幼虫结茧越冬,次年春季孵化并蛀食核桃果实,羽化后成虫寿命较短,仅约1周左右(田敏爵等, 2010)。核桃举肢蛾成虫过短的寿命和特殊的生活史使其无法借助上述力量进行传播。此外,核桃举肢蛾也不具备远距离迁飞的能力。因此,地理阻隔(河流和山川)尤其是大型河流,可能对其种群扩散的阻碍作用则更为明显(Wangetal., 2022)。AMOVA分析的群体变异分析和分层变异分析中较高的种群内变异(92.71%和90.05%)也证实了这一点。
结合种群遗传结构及地理分布来看,核桃举肢蛾可以分为西部种群和东部种群两大分支。西部种群主要分布于秦岭山脉和大巴山一带,包括河南安阳种群(AY)、陕西洛南种群(LN)、陕西眉县种群(YG)、陕西太白种群(TB)、陕西宁陕种群(NS)、陕西镇安种群(ZA)、陕西山阳种群(SY)、陕西安康种群(AK)、甘肃成县种群(CX)和四川南江种群(NJ);东部种群则主要覆盖太行山及其东部和南部平原地区,由北京怀柔区种群(BJ)、河北平山种群(HP)、河南三门峡种群(LB)、山西阳城种群(YC)和陕西商南种群(SN)组成。核桃栽培历史悠久,核桃举肢蛾作为核桃的专食性害虫,其种群分布与宿主植物具有一定程度的一致性。胡昳恒等(2014)利用核基因ITS对秦岭地区和四川的核桃种群遗传结构分析发现,秦岭南北坡核桃群体与东北和四川地区核桃群体存在明显的差异,并进一步确定秦岭核桃自然种群在历史上并未经历种群扩张,表明山系对核桃自然种群的分布影响不大。
此外,陕西山阳种群(SY)与西部秦岭地区种群处于同一分组内,而山东济南种群(LC)又与其他种群差异较大。可能原因是近几十年来,核桃果仁较高的营养价值被广泛认可,经济利益驱使我国甚至世界核桃种植面积快速扩张,促进核桃种苗的远距离运输和移植,进一步促进了核桃举肢蛾的长距离传播和扩散,即核桃举肢蛾种群的地理分布可能受到了人为因素等的干扰,这也是农林害虫种群遗传结构表现异常的主要原因。
猜你喜欢
特产研究(2022年6期)2023-01-17 05:05:06
今日农业(2021年19期)2021-11-27 00:45:49
当代陕西(2021年16期)2021-11-02 06:45:42
疯狂英语·新悦读(2021年5期)2021-06-08 01:54:12
当代陕西(2020年22期)2021-01-18 06:20:06
基层中医药(2018年2期)2018-05-31 08:45:17
——喜迎十九大 追赶超越在陕西
当代陕西(2017年10期)2017-10-16 01:23:39
四川动物(2017年4期)2017-07-31 23:54:19
集美大学学报(自然科学版)(2015年4期)2015-02-28 01:13:37
河北遥感(2014年3期)2014-07-10 13:16:48
