
缺氧诱导因子-1α调控骨形态发生蛋白9对间充质干细胞成骨分化的影响
2024-02-22 06:54丘志河谢卫勇闵水平王安森
临床骨科杂志 2024年1期
李 祥,丘志河,谢卫勇,黄 刚,闵水平,王安森
间充质干细胞(MSCs)在骨髓中含量丰富,具有向骨、肌肉等组织多向分化的潜能,对目前骨组织工程研究具有重要价值[1-2]。MSCs成骨分化受多基因表达调控,骨形态发生蛋白(BMP)是目前唯一可诱导异位成骨的细胞因子,其家族众多成员中BMP9诱导能力较强[3],主要通过经典BMPs-Smads信号通路及非经典丝裂原活化蛋白激酶(MAPK)信号通路调控MSCs成骨分化[4-5]。另有研究[6]报道,BMP9诱导的MSCs成骨分化可能存在微小RNA(miRNA)或其他基因共同参与影响,但具体调控机制尚未完全明确。缺氧诱导因子-1α(HIF-1α)可促进低氧状态下细胞持续分化,促进血管新生[7]。过表达miR-155可能通过调控Smad/BMP信号通路或靶向抑制HIF-1α的表达来减弱BMP9诱导的小鼠MSCs C3H10T1/2成骨分化[8]。然而关于HIF-1α单独对BMP9诱导小鼠MSCs C3H10T1/2成骨分化的影响机制尚未阐明。本研究旨在采用小鼠MSCs C3H10T1/2为细胞模型,探讨HIF-1α对BMP9诱导小鼠MSCs C3H10T1/2成骨分化的影响机制,以期为全面阐明BMP9诱导MSCs成骨分化具体机制提供新的参考。
1 材料与方法
1.1 细胞和重组腺病毒来源小鼠MSCs C3H10T1/2细胞系购自美国菌种保藏中心,BMP9腺病毒(Ad-BMP9)、腺病毒对照液(Ad-GFP,GFP为绿色荧光标记)及重组HIF-1α过表达质粒(pcDNA3.1-HIF-1α)、空质粒(pcDNA3.1-mRFP,mRFP为红色荧光标记)均由上海柯雷生物科技有限公司设计并构建。
1.2 主要试剂及仪器DMEM培养基、胎牛血清(FBS)均购自Gibco公司;茜素红S染料购自Sigma公司;Lipofectamine®2000转染试剂盒购自Invitrogen公司;碱性磷酸酶(ALP)显色试剂盒、ALP活性检测试剂盒、蛋白抽提试剂盒、BCA蛋白定量试剂盒均购自上海碧云天有限公司;BMP9、HIF-1α、骨钙素(OC)、骨桥蛋白(OPN)、Runt相关转录因子2(Runx2)、GAPDH单克隆抗体、羊抗鼠二抗均购自Abcam公司。Forma Steri-Cycle i160型二氧化碳(CO2)培养箱购自Thermo Fisher公司;MODEL550型酶标仪购自Bio-Rad公司;IX75荧光显微镜购自奥林巴斯公司;JS-780型凝胶成像仪购自上海迪奥生物科技有限公司。
1.3 方法
1.3.1细胞培养 C3H10T1/2细胞常规复苏后置于提前配置的含10% FBS、1%青链霉素DMEM培养基中,于37℃、5% 饱和湿度的CO2培养箱中培养。细胞传代用0.25%胰蛋白酶消化。
1.3.2细胞转染及分组 取对数期生长C3H10T1/2细胞接种于6孔细胞培养板,待细胞密度生长至70%左右时,分别加入Ad-BMP9、Ad-GFP腺病毒液、Ad-BMP9+pcDNA3.1-mRFP、Ad-BMP9+pcDNA3.1-HIF-1α,分别设为Ad-BMP9组(转染BMP9过表达的腺病毒液)、Ad-GFP组(转染腺病毒液)、pcDNA3.1-mRFP组(转染Ad-BMP9+pcDNA3.1空载质粒)和pcDNA3.1-HIF-1α组(转染Ad-BMP9+HIF-1α过表达质粒),另选取NC组(正常培养C3H10T1/2细胞),每组设置6个平行样。pcDNA3.1-mRFP组及pcDNA3.1-HIF-1α组转染采用Lipofectamine®2000转染法,具体操作严格按试剂盒说明书进行。48 h后在Ad-BMP9组、Ad-GFP组观察到绿色荧光,在pcDNA3.1-mRFP组和pcDNA3.1-HIF-1α组观察到绿色和红色荧光,进行后续实验操作。
1.3.3ALP活性检测 取1.3.2中各组C3H10T1/2细胞,培养7 d后用磷酸缓冲盐溶液(PBS)冲洗2次,4 ℃无水乙醇固定1 h,ALP染色液染色,避光孵育30 min后观察结果。参照ALP活性测定试剂盒测定ALP活性。
1.3.4茜素红S染色 取1.3.2中各组C3H10T1/2细胞,转染6 h后换液同时添加50 mg/L维生素C、10 mmol/L β-磷酸甘油,继续培养14 d后弃去培养基,PBS洗3次,4 ℃ 0.05%戊二醛固定10 min,去离子水(ddH2O)洗3次,加入0.4%茜素红S染色5 min,ddH2O终止染色并洗涤,显微镜下观察结果。为了进一步定量分析钙化结节,显微镜下拍照后,用PBS洗涤各组细胞2次,加入10%氯化十六烷基吡啶(采用10 mmol/L pH为7.0的磷酸盐配制而成),室温下孵育15 min,将溶解下的染料使用酶标仪在570 nm处测量其吸光度(A)值。
1.3.5各组C3H10T1/2细胞中BMP9、HIF-1α、OC、OPN及Runx2蛋白表达检测 取1.3.2中各组C3H10T1/2细胞,采用蛋白抽提试剂盒提取总蛋白,BCA法测定蛋白浓度,置于-80 ℃冰箱保存待用。采用蛋白免疫印迹(WB)法检测蛋白表达。取50 mg蛋白样品经十二烷基硫酸钠-聚丙烯酰氨凝胶电泳(SDS-PAGE)、转膜、封闭、加入BMP9(1 ∶200)、HIF-1α(1 ∶500)、OC(1 ∶200)、OPN(1 ∶500)、Runx2(1 ∶500)、GAPDH(1 ∶500)一抗孵育,4 ℃过夜,加入羊抗鼠二抗孵育(1 ∶5 000),37 ℃ 1 h,显像、曝光、拍照并分析结果。以靶蛋白灰度值与内参GAPDH灰度值比值作为靶蛋白相对表达量。
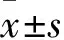
2 结果
2.1 5组C3H10T1/2细胞转染情况及ALP活性检测鉴定转染48 h后,与NC组比较,荧光显微镜下观察到Ad-BMP9组、Ad-GFP组有大量绿色荧光出现,说明Ad-BMP9和Ad-GFP转染成功;pcDNA3.1-mRFP组和pcDNA3.1-HIF-1α组不仅有大量绿色荧光,还有大量红色荧光出现,提示Ad-BMP9与pcDNA3.1-HIF-1α或pcDNA3.1-mRFP均转染成功;见图1。ALP活性:NC组与Ad-GFP组比较差异无统计学意义(P>0.05);Ad-BMP9组、pcDNA3.1-mRFP组、pcDNA3.1-HIF-1α组均较NC组、Ad-GFP组增加(P<0.05);Ad-BMP9组与pcDNA3.1-mRFP组比较差异无统计学意义(P>0.05);pcDNA3.1-HIF-1α组较Ad-BMP9组、pcDNA3.1-mRFP组均增加(P<0.05);见表1。
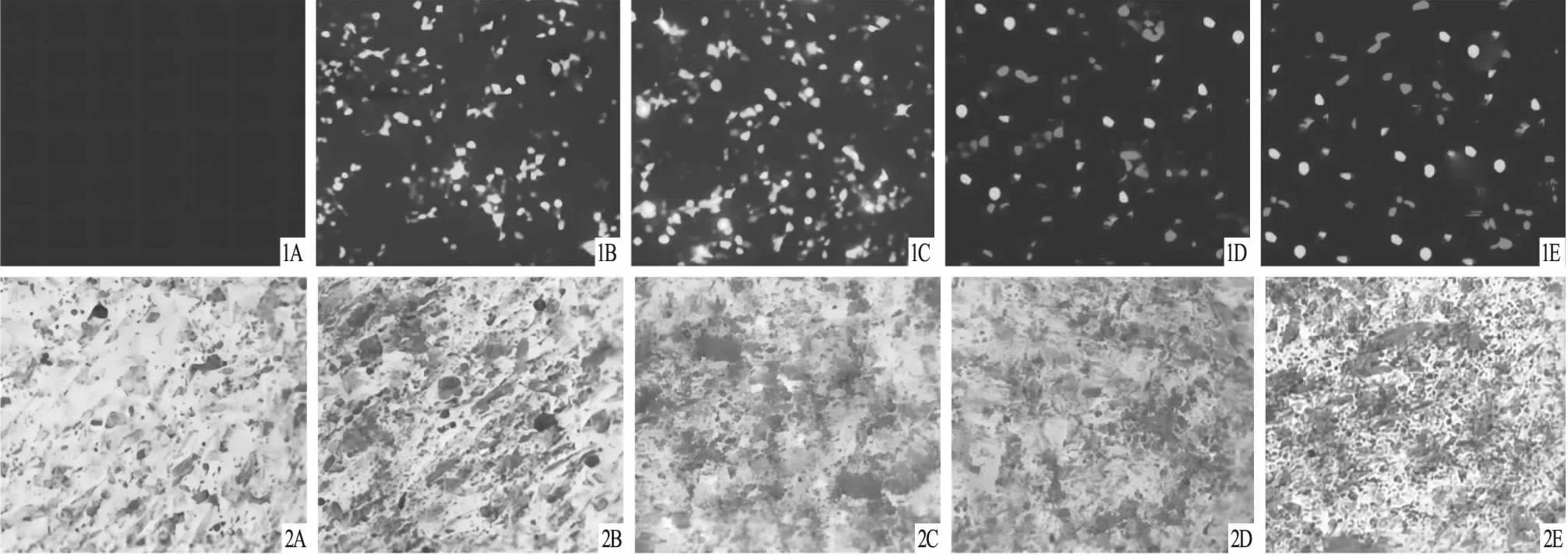
图1 荧光显微镜下5组C3H10T1/2细胞转染情况(×200) A.NC组;B.Ad-GFP组;C.Ad-BMP9组;D.pcDNA3.1-mRFP组;E.pcDNA3.1-HIF-1α组 图2 茜素红S染色检测5组C3H10T1/2细胞中钙化结节情况(×200) A.NC组;B.Ad-GFP组;C.Ad-BMP9组;D.pcDNA3.1-mRFP组;E.pcDNA3.1-HIF-1α组
表1 5组C3H10T1/2细胞ALP活性比较
2.2 5组C3H10T1/2细胞中钙化结节情况比较茜素红S染色结果发现,与NC组、Ad-GFP组比较,Ad-BMP9组钙盐沉积明显增加;与Ad-BMP9组、pcDNA3.1-mRFP组比较,pcDNA3.1-HIF-1α组钙盐沉积明显增加;见图2。定量分析结果显示,NC组、Ad-GFP组、Ad-BMP9组、pcDNA3.1-mRFP组、pcDNA3.1-HIF-1α组的A值分别为0.80~0.92(0.85±0.03)、0.81~0.94(0.87±0.05)、1.23~1.50(1.36±0.09)、1.22~1.56(1.38±0.11)、1.53~1.91(1.73±0.15);NC组与Ad-GFP组比较差异无统计学意义(P>0.05);Ad-BMP9组、pcDNA3.1-mRFP组、pcDNA3.1-HIF-1α组均较NC组、Ad-GFP组增加(P<0.05);Ad-BMP9组与pcDNA3.1-mRFP组比较差异无统计学意义(P>0.05);pcDNA3.1-HIF-1α组较Ad-BMP9组、pcDNA3.1-mRFP组均增加(P<0.05)。
2.3 5组C3H10T1/2细胞中BMP9及HIF-1α蛋白表达水平比较见图3、表2。NC组与Ad-GFP组比较差异无统计学意义(P>0.05);Ad-BMP9组、pcDNA3.1-mRFP组、pcDNA3.1-HIF-1α组均较NC组、Ad-GFP组增加(P<0.05);Ad-BMP9组与pcDNA3.1-mRFP组比较差异无统计学意义(P>0.05);pcDNA3.1-HIF-1α组较Ad-BMP9组、pcDNA-3.1-mRFP组均增加(P<0.05)。
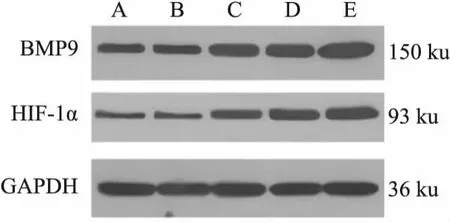
图3 5组C3H10T1/2细胞中BMP9及HIF-1α WB图 A:NC组;B:Ad-GFP组;C:Ad-BMP9组;D:pcDNA3.1-mRFP组;E:pcDNA3.1-HIF-1α组
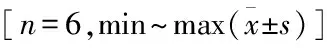
表2 5组C3H10T1/2细胞BMP9及HIF-1α蛋白表达水平比较
2.4 5组C3H10T1/2细胞中OC、OPN蛋白表达水平比较见图4、表3。 NC组与Ad-GFP组比较差异无统计学意义(P>0.05);Ad-BMP9组、pcDNA3.1-mRFP组、pcDNA3.1-HIF-1α组均较NC组、Ad-GFP组增加(P<0.05);Ad-BMP9组与pcD-NA3.1-mRFP组比较差异无统计学意义(P>0.05);pcDNA3.1-HIF-1α组较Ad-BMP9组、pcDNA3.1-mRFP组均增加(P<0.05)。
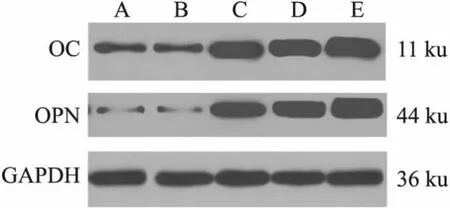
图4 5组C3H10T1/2细胞中OC、OPN WB图 A:NC组;B:Ad-GFP组;C:Ad-BMP9组;D:pcDNA3.1-mRFP组;E:pcDNA3.1-HIF-1α组
表3 5组C3H10T1/2细胞OC、OPN蛋白表达水平比较
2.5 5组C3H10T1/2细胞中Runx2蛋白表达水平比较见图5、表4。NC组与Ad-GFP组比较差异无统计学意义(P>0.05);Ad-BMP9组、pcDNA3.1-mRFP组、pcDNA3.1-HIF-1α组均较NC组、Ad-GFP组增加(P<0.05);Ad-BMP9组与pcDNA3.1-mRFP组比较差异无统计学意义(P>0.05);pcDNA3.1-HIF-1α组较Ad-BMP9组、pcDNA3.1-mRFP组均增加(P<0.05)。

图5 5组C3H10T1/2细胞中Runx2 WB图 A:NC组;B:Ad-GFP组;C:Ad-BMP9组;D:pcDNA3.1-mRFP组;E:pcDNA3.1-HIF-1α组
表4 5组C3H10T1/2细胞中Runx2蛋白表达水平比较
3 讨论
3.1 BMP9对MSCs成骨分化的影响骨组织工程中核心问题为定向诱导MSCs等成骨分化。BMPs家族属于转化生长因子β(TGF-β)超家族,可与细胞表面骨形态发生蛋白受体(BMPR)Ⅰ和BMPRⅡ受体结合,调控细胞分化,对诱导MSCs成骨分化调控起重要作用[9-10]。目前临床较多使用BMP2、BMP7诱导MSCs成骨分化[11-12]。近来研究表明,BMP9促进MSCs成骨分化作用更为强效。任格 等[13]报道,BMP9可促进牙囊干细胞成骨分化。Zhu et al[14]报道,Wnt11可能通过增强BMP9诱导的BMPs-Smads和p38 MAPK信号以增强其在MSCs中诱导成骨分化潜能。本研究以小鼠胚胎成纤维细胞系C3H10T1/2细胞为载体,采用腺病毒包装BMP9诱导其成骨分化,结果显示,Ad-GFP组和Ad-BMP9组均出现大量绿色荧光,且BMP9蛋白表达水平Ad-BMP9组较Ad-GFP组均明显增加;pcDNA3.1-mRFP组和pcDNA3.1-HIF-1α组均出现大量红绿色荧光,且BMP9、HIF-1α蛋白表达水平pcDNA3.1-HIF-1α组均较pcDNA3.1-mRFP组明显增加,提示Ad-BMP、Ad-BMP+pcDNA3.1-HIF-1α均转染成功,可进行后续实验。与Ad-GFP组比较,Ad-BMP9组HIF-1α蛋白表达、ALP活性、钙盐沉积、A值均明显增加,其中ALP活性是成骨早期标志,提示BMP9可诱导C3H10T1/2细胞成骨分化。
3.2 HIF-1α对MSCs成骨分化的影响HIF-1α普遍存在于人和动物细胞中,在缺氧环境下可稳定表达,主要参与调节并维持低氧状态下细胞新陈代谢、促进肝细胞分化[15-16]。左新慧 等[17]报道,HIF-1α可参与骨髓MSCs成骨分化。李松涛 等[15]报道,缺氧条件下,干扰HIF-1α表达可通过促进神经纤毛蛋白1蛋白表达,抑制小鼠单核巨噬细胞白血病细胞(RAW264.7)向成熟破骨细胞分化。Zhou et al[18]报道,HIF-1α是BMP2诱导MSCs软骨分化、成骨分化和软骨内成骨的调节因子,HIF-1α和BMP2可协同促进增殖软骨细胞区扩张。本研究结果发现,与pcDNA3.1-mRFP组比较,pcDNA3.1-HIF-1α组HIF-1α蛋白表达、ALP活性、钙盐沉积、A值均明显增加,提示过表达HIF-1α可能协同BMP9诱导C3H10T1/2细胞成骨分化,与上述文献结果一致,但本研究结果尚不能确定两者协同作用的具体机制。
3.3 HIF-1α调控BMP9对MSCs Runx2表达的影响徐丽 等[19]报道,BMP9诱导C3H10T1/2细胞成骨分化过程中,钙盐沉积、OC、OPN蛋白表达显著增加。陈晓婷 等[20]报道,骨化三醇可增强BMP9诱导MSCs成骨分化,增加其钙盐沉积、OC、OPN蛋白表达,与激活磷脂酰肌醇-3-羟激酶/蛋白激酶B信号通路有关。本研究结果发现,BMP9过表达可提高OC、OPN蛋白表达水平;在过表达BMP9的基础上过表达HIF-1α后,OC、OPN蛋白表达水平进一步增加,提示HIF-1α可协同BMP9促进OC、OPN蛋白表达,进而促进成骨分化。OC是由成骨细胞合成分泌的一种维生素K依赖性钙结合蛋白,大部分沉着于骨骼中;OPN是一种带负电荷非胶原性骨基质糖蛋白,可大量表达于骨细胞、成骨细胞、破骨细胞、软骨细胞等,在骨基质和矿化物吸收过程中起重要作用;以上两者均是中晚期成骨标志蛋白[21]。综合以上研究及本研究结果提示,HIF-1α可能通过促进成骨分化标记物OC、OPN蛋白表达促进BMP9诱导C3H10T1/2细胞钙盐沉积及成骨分化。Runx2是介导成骨分化的重要转录因子,是BMP信号通路下游靶标因子,对MSCs成骨细胞分化起重要作用[22]。Sun et al[23]报道,葡萄糖通过上调Sry相关组蛋白9(Sox9)和Runx2表达调节人软骨终板干细胞组织特异性软骨成骨分化。Ren et al[24]报道,鹿茸水提液通过刺激BMP-2/Smad1、5/Runx2信号通路,促进骨髓MSCs增殖和成骨分化。本研究进一步检测Runx2蛋白表达发现,与Ad-GFP组比较,Ad-BMP9组Runx2蛋白表达水平明显增加;与pcDNA3.1-mRFP组比较,pcDNA3.1-HIF-1α组Runx2蛋白表达水平明显增加,与Ren et al[24]研究报道一致,提示HIF-1α可能通过促进BMP9表达进而上调其下游转录因子Runx2诱导C3H10T1/2细胞成骨分化,但尚不明确Smad通路在此过程中的作用。
综上所述,HIF-1α可促进BMP9诱导MSCs C3H10T1/2成骨分化,可能与上调OC、OPN及其下游Runx2蛋白表达有关,但该过程涉及具体通路尚不清楚,有待进一步研究。
猜你喜欢
传染病信息(2022年3期)2022-07-15
清华金融评论(2022年4期)2022-04-13
口腔医学(2021年10期)2021-12-02
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
灾害医学与救援(电子版)(2018年1期)2018-06-05
上海农业学报(2017年4期)2017-04-10
中华老年口腔医学杂志(2016年2期)2017-01-15
中国病理生理杂志(2015年8期)2015-12-21
吉林大学学报(医学版)(2015年4期)2015-12-17
