
不同密度下小陇山蒙古栎次生混交林树冠结构与密集度的关系
2024-02-22 11:35蒋孟多马映栋李平英包永平
林业科学研究 2024年1期
蒋孟多,马映栋,蒋 睿,李平英,包永平
(1. 甘肃省次生林培育重点实验室,甘肃 天水 741020;2. 新疆大学生态与环境学院,新疆 乌鲁木齐 830000;3. 西北师范大学地理与环境科学学院,甘肃 兰州 730000;4. 甘肃省小陇山林业保护中心滩歌林场,甘肃 天水 741314)
树冠结构属于森林结构的范畴,不仅决定林木形态、未来长势与活力[1],其变化还能降低林木复杂的竞争[2]。研究树冠结构能帮助理解林木利用生态空间的能力,揭示其生长对策和适应机制[3]。密集度是林分空间结构的重要属性,与林分密度有区别,通过判断林分空间结构单元中树冠连接来分析林木疏密程度,反映了树冠的密集程度与林冠层的连续遮盖程度和挤压状况[4]。密集度与树冠结构有关系。通常,密集度取值越大,林地微环境林木密集程度越高,树冠遮盖越连续,即林木竞争就越激烈;取值越小则说明林木密集程度越低,即竞争压力减小,说明此时林木生长稀疏,出现林隙的可能性增加。森林经营中,冠幅常用于表征树冠特征,要探究林木密集程度与树冠结构的关系,单一的树冠特征难以起到普遍性作用且意义不大。就整个林冠层挤压与林木竞争而言,林木密集程度很可能受到除冠幅外的其它树冠结构特征,如树冠面积、冠长和树冠体积等的影响。
针对树冠结构与林木密集程度关系的专门研究迄今极为少见,多数研究集中于树冠结构与林木竞争关系层面。早期研究表明,林分密度能够预测林冠层竞争[5],无论树种组成与林木年龄,树冠发育与胸径增长对整个林分的竞争都非常敏感[6]。尽管树冠结构与林木大小密切相关,但就混交林而言,冠幅因受林木径阶差异的影响,因此很难量化林分种间、种内竞争[7],这就说明树冠结构与林木竞争关系可能与林木大小差异有关。树冠结构的变化与林分密度有关。在分布较稠密的森林中,外力干扰(如大风侵袭)导致冠层出现林隙,树冠结构变的更加复杂[8]。因为风的频繁干扰,树冠脆弱部位容易折断,单木树冠完整性及林冠层均遭到不同程度的破坏,冠幅、冠长和冠幅面积等因此改变。一旦完整的树冠被破坏,树冠结构内部、树与树的挤压状态随之也被调整,林木密集程度可能相应发生变化,且这种变化很可能因林分密度的差异有所不同。对于未经干扰或干扰较轻的林分而言,树冠结构变化可能由林木自疏所致。Gause 假说认为,对同一资源产生竞争的2 个种无法长期共存,结果会导致1 个种占优势,另1 个种则被淘汰[9]。随着树的发育,个体对营养资源与生长空间的需求增加,导致林木竞争更加激烈[10],当光热资源不足以支持所有林木生长时,一些林木生长将减缓,而一些林木将最终死亡[11]。表明无论林木个体还是整个林分,树冠结构与密集度的关系都避免不了林木竞争的作用,它们的关系很可能与林分密度有关。为此,本研究将探究林分密度如何影响树冠结构与密集度的关系。
蒙古栎(Quercus mongolicaFisch. ex Ledeb.)林在小陇山林区北部属主要的次生混交林群落和典型的地带性顶级植被,对维持当地生态平衡、提高环境质量和保护生物多样性具有重要的生态价值。近年来,国内学者从优化森林结构的角度对小陇山蒙古栎林展开了大量研究,比如森林经营措施与林分空间结构[12-14]、群落结构与稳定性[15]等,而有关蒙古栎林树冠结构的研究相对缺乏。近期研究表明,小陇山蒙古栎林的密集度处于中等至密集状态,除与林木冠幅大小有关外,可能与林分密度也有关[15],尚需进一步论证。基于以上背景,本研究依托小陇山林区北部4 个30 m × 30 m 的蒙古栎次生混交林样地,分析不同密度林分树冠结构特征的差异,探究不同密度林分的林木胸径与树冠结构的关系,从而厘清林分密度与树冠结构及密集度的关系,以期深入掌握蒙古栎次生混交林结构特征,为蒙古栎林可持续经营和资源可持续利用提供更加充分的科学依据,也为发挥山地森林的多种生态效能提供理论基础。
1 研究地概况与研究方法
1.1 研究地概况
样地位于甘肃省小陇山林业保护中心滩歌林场境内(104°40'~104°58' E,34°27'~34°39' N)。气候属温带大陆性气候,受季风影响较显著,具有冬冷无严寒、夏热无酷暑的特点。根据武山县气象站资料记载,本地年均温9.6 ℃,年降水量490 mm以上,相对湿度65%,降水集中于7—9 月,约占全年降水量的70%,无霜期140~218 d。研究样地海拔在1 800~2 800 m,坡度为25°~40°,山地垂直带谱明显,土壤主要为棕色土,多石砾。研究区森林覆盖率达47.9%,除次生树种蒙古栎、复叶槭(Acer negundoL.)、椴树(Tilia tuanSzyszyl.)、华山松(Pinus armandiiFranch.)、山杨(Populus davidianaDode)和白桦(Betula platyphyllaSuk.)外,人工栽培树种包括青海云杉(Picea crassifoliaKom.)、华北落叶松(Larix principis-rupprechtiiMayr)、 油 松 (Pinus tabuliformisCarrière)等。
1.2 研究方法
1.2.1 数据采集 野外调查于2020 年8 月进行,选取立地条件典型且均一性较强、人为干扰较少的林分。检尺并记录胸径(DBH)>5 cm 的活立木,用北斗手持导航仪GPS( 型号:G130BD)确定每株树的坐标。用皮尺、围尺、测高仪(型号:BERTHA DENTON-G 系列)测定林木胸径、树高、活枝下高、冠幅(东西与南北方向)等林分因子,编号及记录每株树的健康状态(是否病害、倒伏、弯曲与直立)。按照5 点取样法”在每个样地内取2 m × 2 m 的小样方5 个,调查幼树更新、灌木种类、盖度、高度、株数、基径等(表1)。
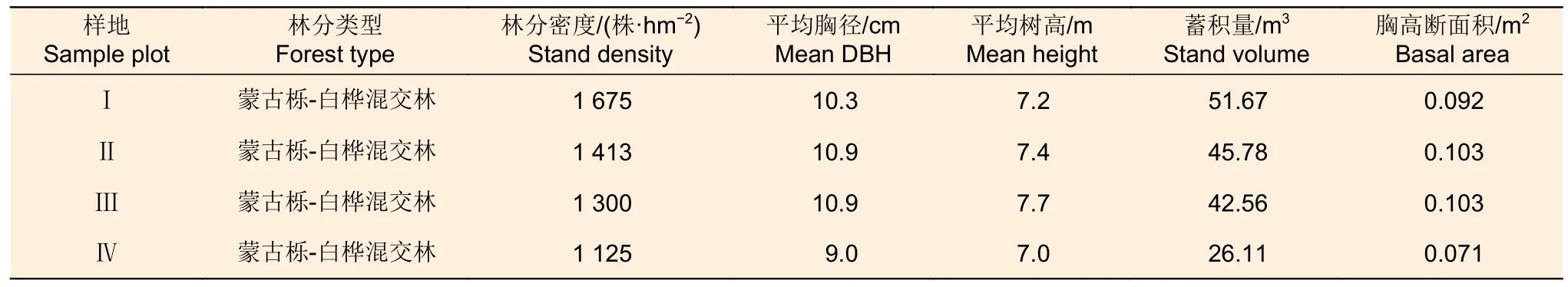
表1 样地概况 Table 1 Details of sample plot in different densities
1.2.2 树冠结构的表达 采用冠幅(CD)、冠长(CL)、冠长率(CR)、冠形率(CS)、树冠面积(CPA)和树冠体积(CVOL)等6 种指标表征树冠结构,其中,冠幅为树冠在东西与南北方向冠宽的均值;冠长为树高与活枝下高之差;冠长率为冠长与树高之比;冠形率为冠长与冠幅之比;树冠面积与树冠体积分别由冠幅和树冠投影计算得到。
1.2.3 密集度与竞争强度的判定 利用重要值法[16]确定蒙古栎林的优势树种。依据惠刚盈等[4,14]对密集度(C)的表述与取值,从林木个体与林分2 种尺度进行密集度分析,同时采用Hegyi指数[13]判别蒙古栎林个体与林分的竞争强度(表2)。

表2 密集度与竞争强度Table 2 Formula of crowding and competition
1.2.4 数据分析 利用Office 软件进行数据预处理,分别利用森林结构分析专业软件Winklemass与R studio 计算密集度与竞争指数。采用最小显著性差异法(LSD)对树冠结构特征进行多重比较,采用Pearson 相关系数评价树冠结构特征与林木胸径大小的相关性。采用结构方程模型(SEM)分析林分密度、树冠结构和密集度的关系,显著性水平均采用P=0.05,其中,依据拟合指数,包括不显著(p>0.05)卡方检验(χ2),SRMR<0.08,GFI>0.95,CHI>0.95,以及最小的AIC 值选取最优的结构方程模型[17]。数据分析主要在R studio 中完成,利用Origin9.4 绘图。为避免边缘效应造成的误差,在样地周围设1.5 m 的缓冲区域。
2 结果分析
2.1 树冠结构的差异
蒙古栎林中,冠幅与树冠面积随着林分密度的递增而递增,冠长与树冠体积的变化趋势相似,并随林分密度的变化呈单峰型,虽然冠长率与冠形率也呈单峰型,但其变化趋势相对平缓,说明不同密度林分的树冠结构特征存在差异(图1a)。分析样地间、样地内树冠结构的变异(图1b),结果表明树冠体积的变异最强,变异程度介于28%~45%;其次为树冠面积与冠长,分别为18%~28%与13%~20%;而冠幅、冠长和冠长率的变异相对较弱,分别介于9%~14%、7%~9%和13%~16%。总体上,蒙古栎林树冠结构的变异未与林分密度保持一致性,而是随着林分密度增大呈现出“先增后减”的单峰型特点。
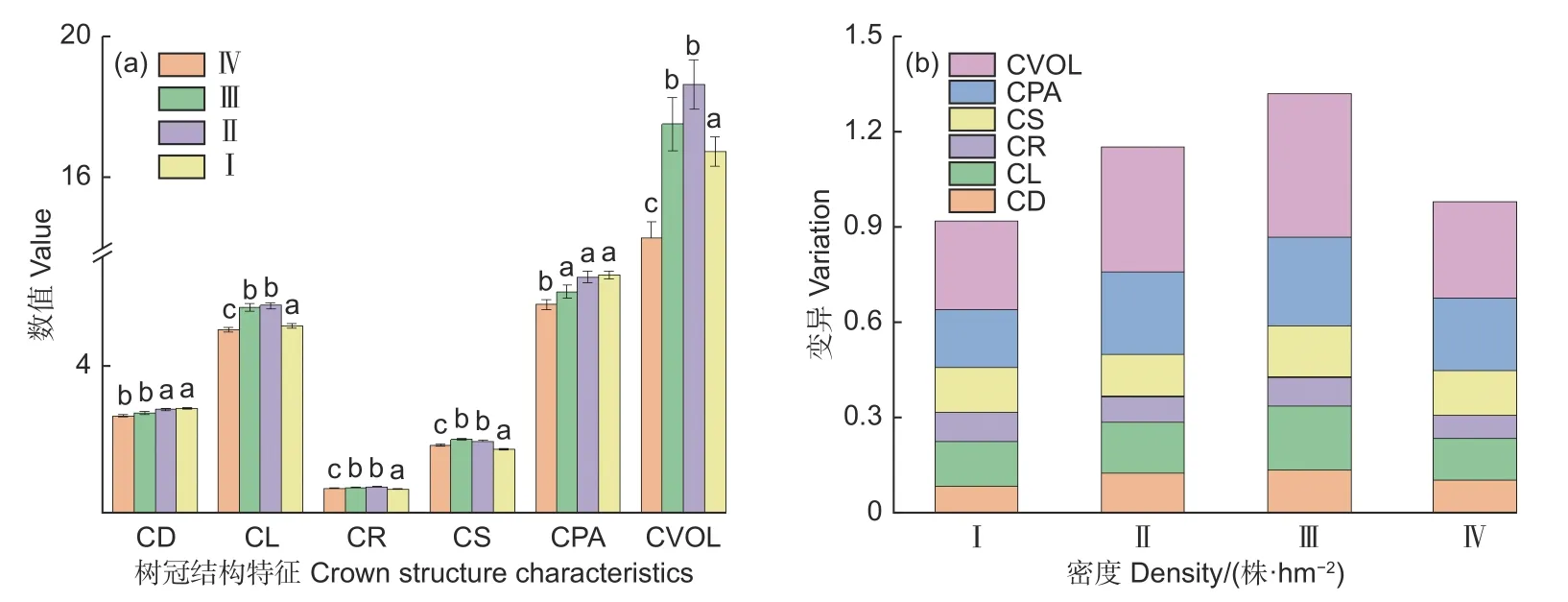
图1 树冠结构的差异Fig. 1 Difference of crown structural characteristic in different stands
2.2 胸径与树冠结构的关系
如图2 所示,样地Ⅰ中林木胸径与树冠结构特征的相关性均显著,胸径与树冠体积的相关性最强(r=0.823,P<0.01),而与冠形率(r=0.188,P<0.05)和冠长率(r=0.216,P<0.05)的相关性较弱。样地Ⅱ中树冠面积(r=0.68,P<0.01)、冠幅(r=0.67,P<0.01)分别与胸径的相关性最强,其次为树冠体积(r=0.65,P<0.01)。与样地Ⅱ相似,样地Ⅲ中冠幅、树冠面积分别与胸径的相关性最强(P<0.01),其次为树冠体积(P<0.01),但2 个样地的冠形率、冠长率与胸径无显著相关性。样地Ⅳ中树冠面积、冠幅、树冠体积分别与胸径的相关性最强(P<0.01),而冠形率、冠长率与胸径的相关性较弱(P<0.05)。总体上,随林分密度的递减,冠幅、树冠面积、树冠体积分别与胸径的相关性趋势近似于“L”型,而冠长—胸径的相关性随林分密度的递减逐渐减弱,林分密度的改变对冠长率—胸径、冠形率—胸径相关性的影响不大。
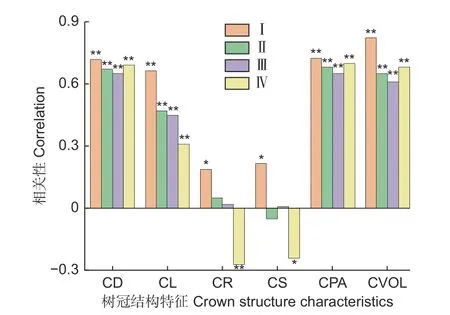
图2 树冠特征与胸径的相关性Fig. 2 Correlation between crown structural characteristics and DBH
2.3 个体与林分的密集程度
蒙古栎林中,4.5%的林木处于很稀疏状态,15.9%为稀疏,28.8%为中等密集,20.9%为较密集,29.7%为很密集,林分密集程度处于中等密集至很密集状态。具体而言(图3),样地Ⅰ与样地Ⅲ中最大相对频率分别为45.2%、27.8%的林木属于很密集,而样地Ⅱ和Ⅳ中最大相对频率分别为32.7%、40.6%的林木属于较密集与中等密集。分析优势树种的密集程度可知(表3),其最大值为0.909,最小值为0.292,包括稀疏、中等密集、较密集和很密集4 种状态,但多数优势树种处于中等密集、较密集、很密集状态。就各样地优势树种的密集程度而言,样地Ⅰ中蒙古栎、华山松和椴树的值最大,且至少54.1%的林木达到很密集,尽管处于很密集的白桦较少,但其中也有46.2%达到中等密集。样地Ⅱ中34.1%的蒙古栎达到较密集,40%的白桦很密集,而华山松与椴树分别均匀分布于中等密集至很密集状态。样地Ⅲ中35%的白桦与57.1%的椴树达到了很密集,而此时多数蒙古栎和华山松处于稀疏与较密集状态。样地Ⅳ的优势树处于很稀疏至中等密集状态,其中蒙古栎31.2%、华山松50%、白桦41.7%以及椴树50%分别为稀疏、很稀疏、稀疏和中等密集,说明优势树种对蒙古栎林密集程度起重要作用。
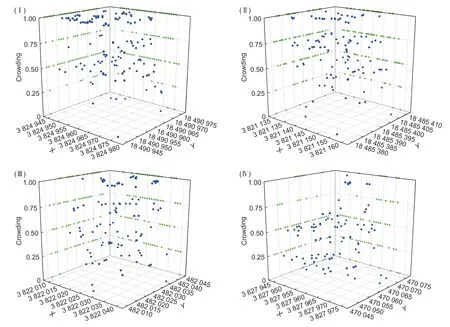
图3 林木个体的密集程度Fig. 3 Crowding of individual trees in different stand densities

表3 优势树种的密集程度 Table 3 Crowding of dominant tree species in different densities
2.4 树冠结构与密集度的关系
最优结构方程模型表明(图4,表4),林分密度对树冠结构与密集度关系的影响包含两种路径:一方面,林分密度通过林木竞争间接影响了树冠结构(p<0.05),通过树冠结构的介导作用从而间接影响密集度(p<0.05);另一方面,林分密度通过林木竞争先间接影响林木胸径(p<0.001),再通过林木胸径的介导作用影响树冠结构,进而对密集度产生影响(p<0.05);其中,就树冠结构特征而言,冠幅与冠长的显著作用不可忽视(p<0.001)。尽管如此,但林分密度直接通过林木竞争也可间接影响密集度(p<0.001)。另外,林分密度通过影响林木竞争与胸径大小进而对密集度产生间接影响,但此影响并不显著(p>0.05)。进一步判定蒙古栎林的林分密度、林木胸径与树冠结构对密集度的联合影响可知,林分密度能解释林木竞争47.1%的变异,林木竞争解释了胸径0.1%的变异,林木胸径与竞争共同解释了树冠结构62.8%的变异,林木竞争、胸径大小与树冠结构共同解释了密集度23.6%的变异。

图4 林分密度、林木竞争、树冠结构和密集度的最优结构方程模型Fig. 4 The best-fitness SEM of stand density, tree competition, crown structure and crowding
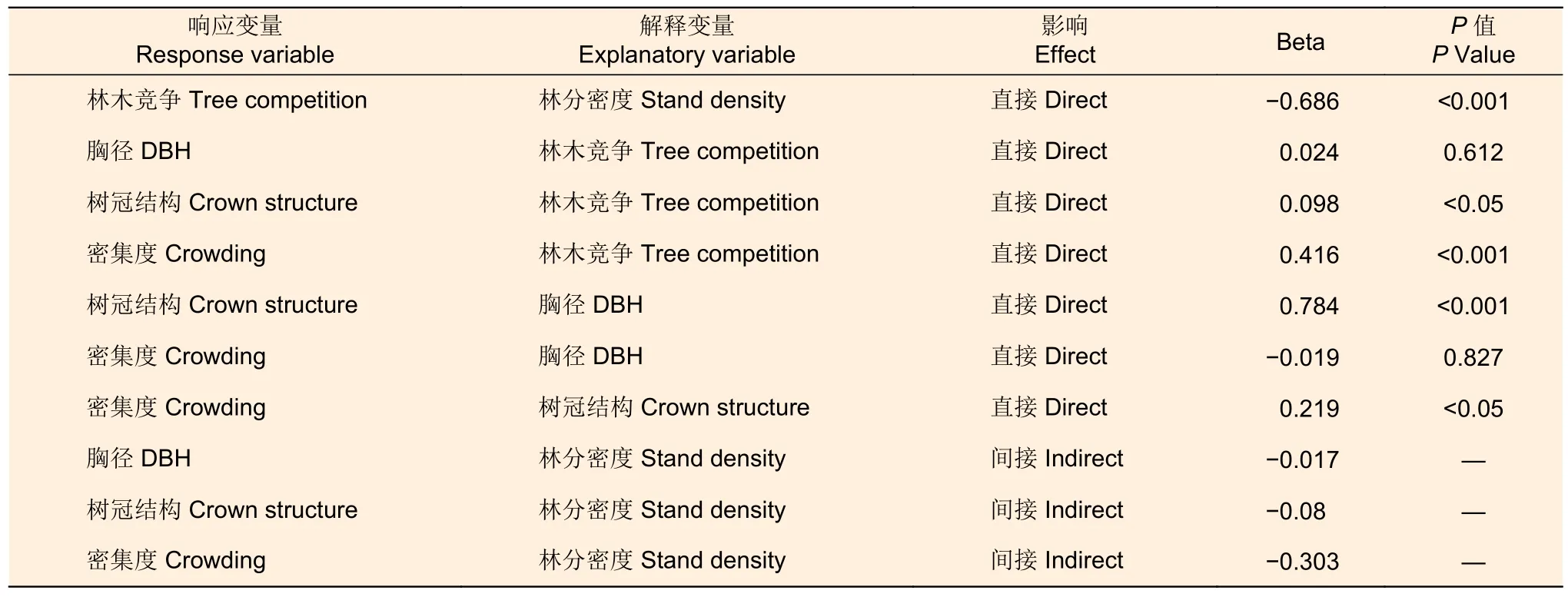
表4 林分密度、林木竞争、树冠结构对密集度的直接与间接影响Table 4 Direct and indirect effect of stand density, tree competition, crown structure on crowding
3 讨论
树冠结构的变化体现了林木适应不同生境的生态策略,是林木在环境驱动下兼容个体特征、树种特征、群落特征、资源权衡等多种因素的综合表征。树冠垂直与横向扩展可衍生多种林木特征与功能,这些特征能替代性的指示林木生长,以维持森林正常的功能,如冠幅能预测树木长势[1]。Hemery[18]认为,冠幅、树冠面积与林木胸径大小有关,其在蒙古栎林中得到了进一步证实。万志兵[19]等认为檫树人工林中胸径与树冠体积、冠长能达到显著相关,该结论在蒙古栎次生林中亦成立;不仅如此,蒙古栎林的林木冠长率、冠形率与林木胸径也存在显著相关。林木胸径与树冠结构的这种密切关系可能是林木为适应环境,长期进化形成的生长对策,因为林木生存空间一旦扩大后,目标树主要将同化物质向侧枝分配促进了树冠生长,目的是为获取更多的光热资源。Gonzalez-Benecke[20]等认为立地条件(气候、土壤)影响林木胸径与树冠结构的关系。然而,本研究由于调查样地相邻,立地环境基本一致,因此胸径与树冠结构的相关性并未受立地条件的影响;反而,林分密度对胸径与树冠结构的关系有影响,这与Brumelis[21]等在云杉林的研究结果基本一致。林分密度反映了林木占据空间资源和获取光照条件的能力,本研究发现,蒙古栎林树冠结构与林木胸径的关系在不同密度林分间的差异较大,尤其随着林分密度增大,冠长与胸径的相关性越强。这可能是林木在树冠水平空间严重受限时采取的向上生长的生态对策,林木通过变粗、延伸冠长促使树冠上下不同部位的枝条获取更多的光照资源[3]。
树冠结构对林分挤压条件的变化非常敏感,能反映林木在同一林分的竞争状况[22],树冠结构越复杂, 不同冠层的林木可通过树冠间隙获得越多光资源,形成单位面积上更加密集的树冠[23],表明树冠结构与密集度之间存在相互影响的关系。本研究,林分密度不仅通过林木竞争间接影响密集度,还通过林木竞争—树冠结构的共同介导作用间接影响密集度。究其原因可能是:一方面,林分密度过大加剧了林木竞争,进而引发自疏,一旦自疏发生后,长势良好的林木优先利用扩大的生长空间伸枝展叶获取营养资源,与一些弱势木及幼树在空间上形成复层林,最终冠层挤压条件因林分密度的调整而改变,从而影响了密集度。调查发现,蒙古栎林内林隙现象较严重且呈斑块状分布,林隙中间及其边缘木树冠形态、胸径长势差异较大,尽管林隙内幼树更新较好,但个别优势树种,如蒙古栎、华山松、白桦与椴树等处于林上层,林隙内与其边缘木的密集度差异较大。另一方面,也可能与蒙古栎林不同树种的生长特性有关。研究认为,具有不同生长潜力的树种占据不同的冠层位置, 通过延伸不同高度的树枝和叶片, 共同促进了物种共存[24]。由于资源互补效应,蒙古栎林不同树种间的生态位差异导致了林木对光热资源更加充分的利用,使林木之间的竞争作用减少,从而导致树冠结构对密集度的直接影响。另外,优势树种蒙古栎、华山松、白桦和椴树对林分密集度起了关键性作用,因此树冠结构的这种直接影响很可能与优势树种有关。本研究,林分密度并未通过林木竞争—胸径的共同作用显著影响密集度,而是通过林木竞争—胸径—树冠结构对密集度产生显著影响,其中,胸径对树冠结构有极显著影响,且林木竞争与胸径对树冠结构变异的解释度最高。这与石小龙等[25]、龙时胜[26]等在人工油松林、青冈栎次生林中的发现一致,即林木胸径与冠幅呈线性关系,林木越粗,其冠幅也越大,且随着林木径阶越大,林木树冠变大的同时,树冠的影响半径也在增大。这可能是由于随着林木生长发育,其对空间资源的需求越强,林木通过扩大树冠(延展冠幅横向生长,增加冠长向上生长)的策略提高光合能力的同时也要依靠增大胸径来提高树干的支撑能力。
树冠结构的变异能保证林木更能适应林分拥挤状态[27]。蒙古栎林的树冠体积、树冠面积、冠幅与冠长的变异较强,其中,冠幅与冠长对构建树冠结构与密集度的路径关系(结构方程模型)起了主要作用,林分密度影响蒙古栎林树冠结构的变异。Vieilledent[28]等认为,树冠结构的强度变异导致了光照条件随时间的高度异质性,反过来,光照不够或采光不均匀又影响树冠生长,表明树冠结构的变异可能还与林地光环境有关。本研究,蒙古栎林的密集度处于中等密集至很密集状态,其中,处于很密集状态的林木比例最大(29.7%),这意味着林分内树冠连接很密,树冠相互重叠过高,可能一定尺度内密集连接的树冠降低了林木对光热资源的利用强度,从而限制了林木生长且降低了林下更新。然而,关于光环境驱动树冠结构变异的机理尚需深入研究,尤其树冠体积、冠幅与冠长应作为树冠结构的重要指标加以关注。此外,树冠生长过程中林冠层过于拥挤会直接导致树体损伤,应采用疏伐扩大生长空间的方式来预防[29]。
4 结论
森林的密度效应一直是森林经营关注的问题,以小陇山蒙古栎次生混交林为研究对象,采用单因素方差分析、相关性分析和最优结构方程模型,探究了蒙古栎林树冠结构与密集度的关系。研究发现,不同密度蒙古栎林的树冠结构有差异,且树冠结构特征与林木胸径大小有关,林分密度影响了蒙古栎林树冠结构与密集度的关系。要实现蒙古栎林可持续经营,不仅要考虑林木胸径大小,更要关注树冠结构特征,通过调整林分密度可优化树冠结构,改善林冠层挤压状况,以促进森林更新与增加物种多样性。然而,本研究仍存在不足之处,如研究样地较少,林分类型单一,以至于设置的林分密度梯度较少,导致无法深入分析树冠结构与密集度的关系。今后的研究应选择补充不同立地条件的样地,并考虑加入环境要素(海拔、水分、土壤等),以充分揭示蒙古栎林结构中的密度效应问题,为小陇山蒙古栎林的合理经营提供科学依据。
猜你喜欢
东北林业大学学报(2023年6期)2023-05-31
丽水学院学报(2022年2期)2022-04-19
今日农业(2021年9期)2021-11-26
林业科技情报(2021年3期)2021-09-01
英语文摘(2021年2期)2021-07-22
兵器装备工程学报(2020年11期)2020-12-16
弹道学报(2020年2期)2020-07-06
电子测试(2018年4期)2018-05-09
中南林业调查规划(2017年1期)2017-12-19
光学精密工程(2016年3期)2016-11-07
