
松材线虫入侵对湖北三峡地区马尾松林水源涵养能力的影响
2024-02-22 11:35宋学雨简尊吉王少博党英侨王小艺肖文发
林业科学研究 2024年1期
宋学雨,简尊吉,王少博,党英侨,魏 可,王小艺,肖文发
(中国林业科学研究院森林生态环境与自然保护研究所,国家林业和草原局森林保护学重点实验室,北京 100091)
松材线虫(Bursaphelenchus xylophilus)是目前对我国松林危害最为严重的外来有害生物[1]。松材线虫入侵自1982 年在江苏省南京市被首次发现以来,40 年间在我国不断扩散蔓延,并在未来气候变暖情景下呈进一步向北、向西扩张趋势[2]。2021 年全国共有19 个省(自治区、直辖市)的742 个县级行政区发生松材线虫病(Pine wilt disease),发生面积171.65 万公顷,病死树数量1 407.92 万株[3]。松材线虫的入侵改变了森林的结构与稳定性,进而影响了森林提供各种生态系统服务功能的正常发挥[4-5]。
三峡地区是我国重要的水源涵养区,同时也是生态脆弱区。马尾松(Pinus massonianaLamb.)是我国南方山地主要的造林树种之一,发挥着木材生产、水土保持、水源涵养等生态系统服务功能,也是三峡地区非常重要的防护林类型之一[6]。近年来,松材线虫病疫情在三峡地区持续扩散蔓延。以宜昌市为例,该病疫情经过15 年发展已经扩展到12 个县(市、区)疫区74 个乡镇(街办、林场)522 个村(居委会),松林受到不同程度危害[7]。马尾松林群落多样性下降和稳定性降低[8-9],水土保持和水源涵养等功能受到影响,甚至威胁三峡地区生态环境和长江水质安全。林分内枯落物层和土壤层的持水能力是森林水源涵养功能研究重点[10],主要体现在拦蓄降水、净化水质和削减洪峰等方面[11]。目前,已有研究分析了松材线虫入侵对三峡地区马尾松林的生物多样性、土壤养分维持等生态系统功能的影响[12-14],而有关松材线虫入侵对三峡地区马尾松林枯落物层和土壤层水源涵养能力影响的研究尚未见报道。此外,依据群落演替理论和生态位补偿假说,森林群落恢复将会缓解和改善松材线虫入侵带来的危害[15],但松材线虫入侵对马尾松林生态系统服务功能的影响及其长期效应还不甚清楚,这不利于科学制定相应的防控措施和林分恢复管理策略。
本研究通过空间替代时间的方法,以湖北三峡地区不同入侵年限(未入侵(0 年)、入侵6 年、入侵10 年和入侵16 年)的马尾松林为对象,采用室内浸泡法测定林下枯落物的有效拦蓄量和最大持水量,利用环刀法测定林下土壤层的有效蓄水量、毛管持水量和饱和蓄水量,并使用枯落物有效拦蓄量和土壤有效蓄水量来估算林分综合蓄水量,量化马尾松林枯落物层和土壤层的水源涵养能力,比较其在松材线虫入侵不同年限间的差异,揭示松材线虫入侵对湖北三峡地区马尾松林枯落物层和土壤层水源涵养能力的影响。为松材线虫入侵后的马尾松林生态系统服务功能的维持和恢复,是否需要采取人为干预措施以及干预程度提供理论依据。
1 研究区概况
研究地点(表1)位于湖北省宜昌市,地处长江上游与中游的结合部,鄂西山区向江汉平原的过渡地带,共辖5个市辖区、3个县级市、3个县、2个自治县。气候类型为亚热带季风气候,四季分明,水热同季,寒旱同季。年均气温16.9 ℃,多年平均降水量1 215.6 mm,多集中在4—9 月。土壤类型以黄壤、黄棕壤为主。森林资源丰富,生物种类多样。森林覆盖率(不含灌木林)达到55.3%,活立木蓄积量3 986 万m3,地带性植被为常绿阔叶林。乔木树种主要有马尾松、槲栎(Quercus alienaBlume)、猴樟(Cinnamomum bodinieriH.Lév.)等,其中以马尾松为优势种。林下灌木主要有宜昌荚蒾(Viburnum erosumThunb.)、木姜子(Litsea pungensHemsl.)、中华绣线菊(Spiraea chinensisMaxim.) 和 檵 木 (Loropetalum chinense(R. Br.) Oliver)等,林下草本植物主要有鸡矢藤(Paederia foetidaL.)、蕺菜(Houttuynia cordataThunb.)等。
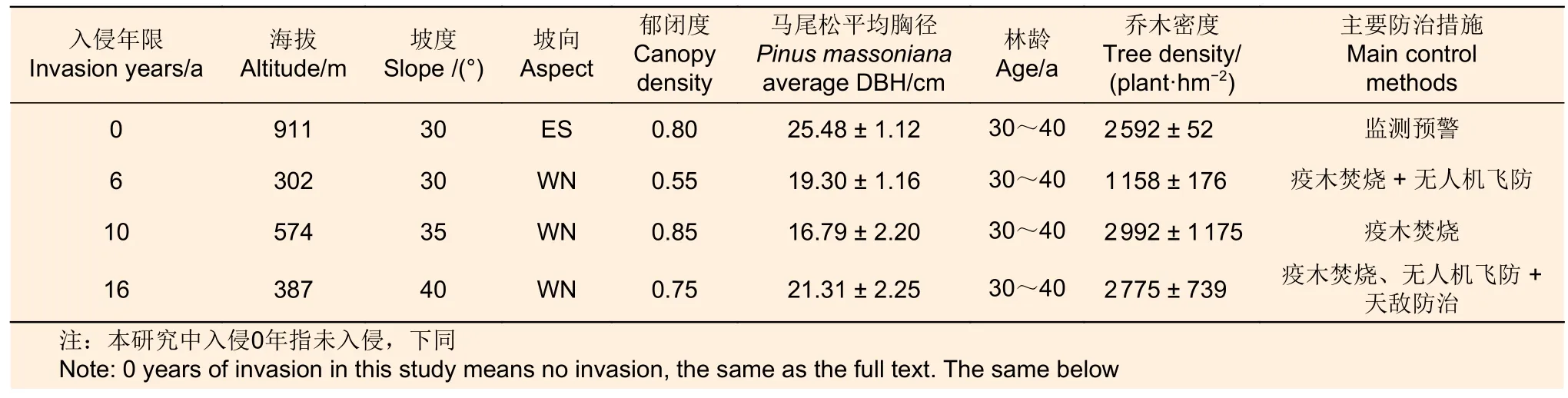
表1 样地基本情况Table 1 Basic situation of sample sites
2 研究方法
2.1 样地设置与调查
2022 年6 月,根据前期踏查结果,在湖北省宜昌市秭归县和夷陵区选取林龄相对一致(约35 a)的马尾松人工林为对象,马尾松林分受松材线虫病入侵前均为纯林。自2006 年发生松材线虫入侵以来,林业部门每年都会对马尾松林进行踏查并伐除、焚烧受害木。本研究根据立地条件基本一致的原则,按照松材线虫入侵年限的不同,选取未入侵(0 a)、入侵6 a、入侵10 a 和入侵16 a 马尾松林作为调查样地。每种林分设置3 个重复样地,每样地大小为20 m × 20 m,3 个重复样地基本具有一致的立地条件、林分条件及土壤类型,共12 个样地。调查样地内自发生病害以来持续至今,每年仍有因松材线虫入侵导致的病死树,说明选择的调查样地长期受松材线虫病的影响。对每个样地内乔木层进行每木检尺,测定其胸径、树高、冠幅等指标,并记录经纬度、海拔和地形等因子,样地详情见表1。
2.2 枯落物收集与持水能力测定
采用五点取样法进行枯落物样品采样。在每个样地内的四角和中心位置,分别设置1 个0.5 m ×0.5 m 的小样方,按全收获法收集枯落物。枯落物按未分解和半-已分解层,分别测其厚度并分层带回,称鲜质量。将称量后的枯落物,在65 ℃烘箱中烘干至质量恒定后,称其干质量,计算单位面积枯落物蓄积量。
枯落物持水量采用室内浸泡法[16]。取烘干后的部分枯落物样品装入100 目尼龙袋中,放入清水容器中,分别浸泡0.5、1、2、4、6、8、10、12、24 h 后取出,静置5 min,直至不滴水为止,迅速称其湿质量。计算其吸水速率、持水量、有效拦蓄量[17]和最大持水量[7],有效拦蓄量和最大持水量计算公式为:
式中,P为有效拦蓄率/%;Rm为最大持水率/%;R0为自然含水率/%;W为有效拦蓄量/(t·hm-2);Wm为最大持水量/(t·hm-2);M为枯落物蓄积量/(t·hm-2);Km为浸泡24 h 后枯落物样品质量/g;K为枯落物样品干质量/g。
2.3 土壤采样与持水能力测定
采用五点取样法进行土壤样品采样。在每个样地内的四角和中心位置,收集完枯落物后,使用100 cm3的标准环刀,按照0~10、10~20 cm 土层取原状土,带回实验室,按照《森林土壤水分一物理性质的测定(LY/T 1215—1999)》[18]测定土壤自然含水率、土壤密度、土壤孔隙度等物理性质,并计算土壤有效蓄水量、毛管持水量和饱和蓄水量[19],计算公式为:
式中,W0为有效蓄水量/(t·hm-2);Wc为毛管持水量/(t·hm-2);Wt为饱和蓄水量/(t·hm-2);P0和Pc分别为土壤非毛管孔隙度/%和毛管孔隙度/%;h为土层厚度/m。
2.4 数据处理
使用Microsoft Excel 2019 进行数据整理。使用SPSS 22 对不同入侵年限马尾松枯落物层蓄积、持水特性和土壤层物理性质进行单因素方差分析(ANOVA)和多重比较(LSD)。采用Origin 2017 作图。
3 结果与分析
3.1 枯落物层水源涵养能力变化
3.1.1 枯落物蓄积量 松材线虫入侵不同年限间马尾松林枯落物厚度存在显著差异,其中,入侵6 a 林分枯落物厚度未分解层厚度(F=12.722,P<0.01)、半-已分解层厚度(F=3.611,P=0.019)和总厚度(F=9.606,P<0.01)均显著小于其他林分(图1A)。枯落物总厚度由大到小依次为入侵0 a(5.34 cm)>入侵16 a(5.06 cm)>入侵10 a(5.03 cm)>入侵6 a(3.47 cm)。松材线虫入侵不同年限间枯落物未分解层蓄积量、半-已分解层蓄积量和总蓄积量均差异不显著(P>0.05,图1B)。枯落物厚度在松材线虫入侵6 a 达到最低值,可能是由于松材线虫侵染和人为干扰导致。蓄积量总体呈波动变化但差异不显著,表明松材线虫的入侵未使松林枯落物蓄积量受到严重干扰,且林分具有一定自我恢复能力。

图1 松材线虫入侵不同年限间马尾松林枯落物厚度和蓄积量Fig. 1 Litter thickness and accumulation of Masson pines in different invasion years by pinewood nematode
3.1.2 枯落物持水能力 松材线虫入侵不同年限,4 种马尾松林分未分解层、半-已分解层枯落物最大持水量和有效拦蓄量均差异不显著(P>0.05, 图2)。未分解层有效拦蓄量为3.43~6.65 t·hm-2;半-已分解层有效拦蓄量为5.24~8.11 t·hm-2(图2A)。未分解层最大持水量为4.12~8.03 t·hm-2;半-已分解层最大持水量为6.44~9.77 t·hm-2(图2B)。枯落物层持水能力与枯落物层蓄积量规律一致,即蓄积量越大,枯落物层持水能力越强。4 种林分未分解层有效拦蓄量和最大持水量,随松材线虫入侵年限呈先下降后回升趋势。入侵16 a 枯落物持水能力相对入侵10 a 林分略有下降但高于入侵6 a 林分和未入侵林分,可能是由于松材线虫持续危害导致。

图2 松材线虫入侵不同年限间马尾松林枯落物持水能力Fig. 2 Water holding capacity of litter of Masson pines in different invasion years by pinewood nematode
3.1.3 枯落物持水过程 不同入侵年限不同层次马尾松林枯落物持水量,均随浸水时长增加而不断增加,枯落物未分解层和半-已分解层吸水量均在前1 h 内增加最快,在1~12 h 浸水时间内持水量缓慢增加,速率逐渐减缓,持水量呈对数型增长,24 h 后达到饱和(图3)。半-已分解层持水量显著大于未分解层,但两者持水量排序不一致,未分解层持水量由大到小依次为入侵10 a>入侵16 a>未入侵林分>入侵6 a(图3A),半-已分解层持水量由大到小依次为入侵10 a>入侵6 a>未入侵>入侵16 a 林分(图3B)。对不同入侵年限林分枯落物持水量与时间进行函数拟合,拟合关系式为:QQQ=aaalllnnn(ttt)+bbb。式中:Q为枯落物持水量/(t·hm-2),t为枯落物浸水时间/h,a为系数,b为常数项,拟合系数在0.83 以上,表明拟合结果良好(表2)。4 种林分枯落物未分解层和半-已分解层吸水速率均随浸水时间延长而不断减小,呈现反“J”型(图3)。在前1 h 内速率最大,1 h 后达到动态平衡,2 h 后速率逐渐趋于0。半-已分解层吸水速率显著大于未分解层,未分解层吸水速率由大到小依次为入侵10 a>入侵16 a>入侵6 a>未入侵林分(图3C),半-已分解层吸水速率由大到小依次为入侵10 a>入侵6 a>未入侵>入侵16 a(图3D)。对不同入侵年限林分枯落物持水速率与时间进行函数拟合,拟合关系式为:V=kt-d。式中:V为枯落物持水速率/(t·hm-2·h-1),t为枯落物浸水时间/h,k为系数,d为常数项,拟合系数在0.90 以上,表明拟合结果良好(表2)。
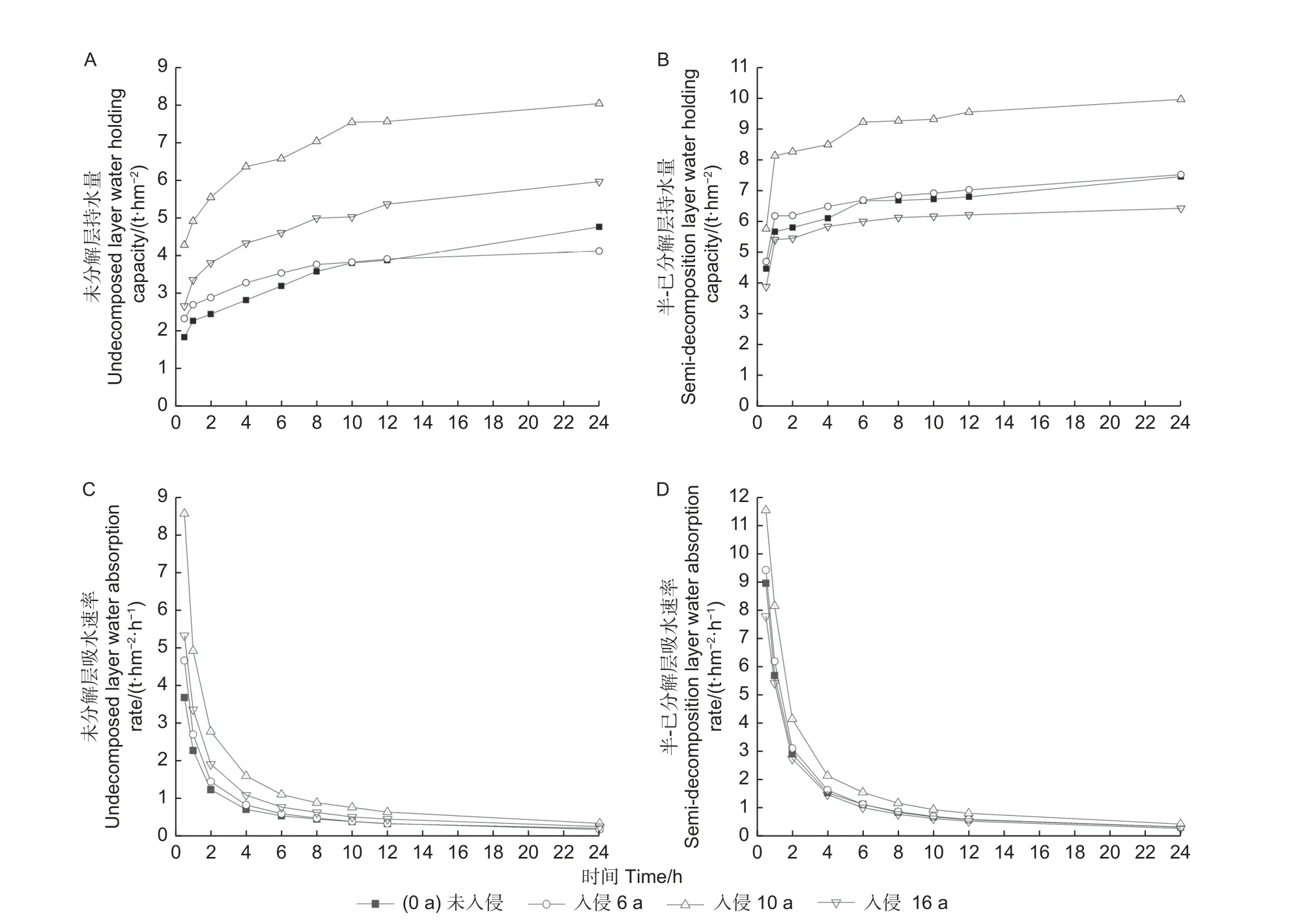
图3 松材线虫入侵不同年限间马尾松林枯落物持水量(A、B)与吸水速率(C、D)变化Fig. 3 Water holding capacity (A, B) and water absorption rate (C, D) of litter of Masson pines in differentinvasion years by pinewood nematode

表2 枯落物持水量、吸水速率与浸水时间的关系Table 2 Relationship between water holding capacity, water absorption rate, and soaking time of litter
3.2 土壤层水源涵养能力变化
3.2.1 土壤物理性质 松材线虫入侵不同年限间马尾松林土壤0~10 cm(F=4.528,P=0.007)、10~20 cm(F=8.303,P<0.01)土层含水量差异极显著,其中松材线虫入侵6 年的马尾松林0~10 cm 和10~20 cm 土层土壤含水量(11.09%和8.89%)最小(表3)。不同入侵年限间土壤0~10 cm(F=7.28,P<0.01)、10~20 cm(F=6.254,P<0.01)土层土壤密度差异极显著,其中未入侵林分0~10 cm 和10~20 cm 土层土壤密度(1.41 g·cm-3和1.51 g·cm-3)最大,入侵10 a 林分土壤密度(1.22 g·cm-3和1.33 g·cm-3)最小(表3)。不同入侵年限间马尾松林土壤0~10 cm(F=3.612,P=0.019)和10~20 cm(F=6.691,P<0.01)土层非毛管孔隙度之间存在极显著差异,0~10 cm 和10~20 cm 土层非毛管孔隙度由大到小均为入侵10 a>未入侵>入侵16 a>入侵6 a(表3)。不同入侵年限马尾松林0~10 cm 和10~20 cm 土层毛管孔隙度和总孔隙度均差异不显著(P>0.05)。各林分土壤含水量、非毛管孔隙度和总孔隙度均值均随入侵年限增加均呈先降低后上升趋势,表现出感病初期土壤物理性质受松材线虫一定程度干扰,随着年限增长逐渐恢复。

表3 松材线虫入侵不同年限间马尾松林土壤物理性质Table 3 Soil physical properties of Masson pines in different invasion years by pinewood nematode
3.2.2 土壤持水能力 松材线虫入侵不同年限间马尾松林0~10 cm(F=3.612,P=0.019)和10~20 cm(F=6.691,P<0.01)土层土壤有效蓄水量存在极显著差异,均以入侵10 a 土壤有效蓄水量最大(100.33 t·hm-2和104.62 t·hm-2),入侵6 a 土壤有效蓄水量最小(62.08 t·hm-2和50.08 t·hm-2,表4)。土壤各土层毛管持水量与0~10 cm 土层饱和蓄水量差异不显著(P>0.05),但10~20 cm 土层饱和蓄水量存在极显著差异(F=8.063,P<0.01),入侵6 a 林分饱和蓄水量(386.27 t·hm-2)显著小于其余林分(表4)。4 种林分土壤有效蓄水量与饱和蓄水量总和均在入侵初期与未入侵相比呈下降趋势,随入侵年限增加逐渐上升,表明土壤储水能力提升。
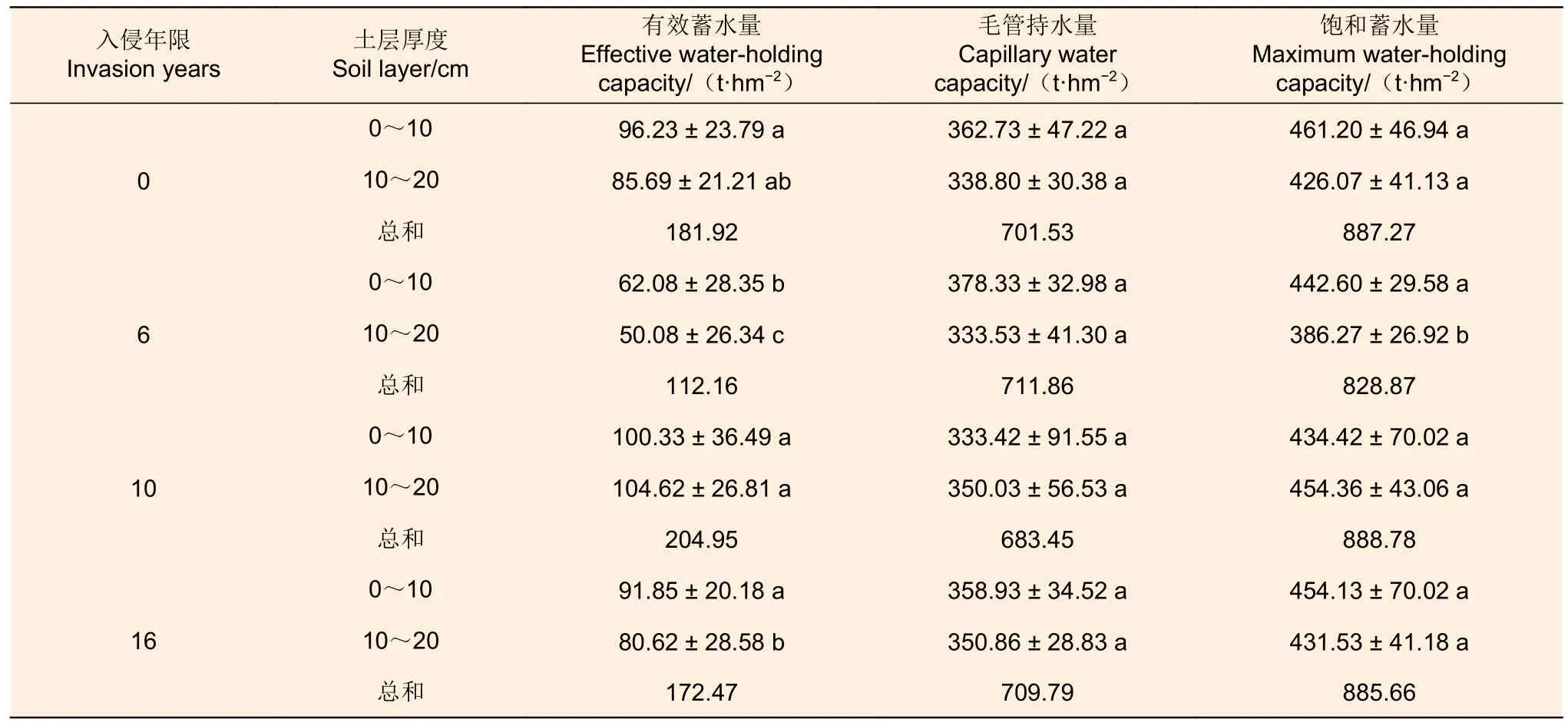
表4 松材线虫入侵不同年限间马尾松林土壤持水能力Table 4 Soil water holding capacity of Masson pines in different invasion years by pine wood nematode
3.3 不同入侵年限间林分枯落物和土壤水源涵养能力综合分析
计算不同入侵年限枯落物层有效拦蓄量和土壤层有效蓄水量之和,定量比较4 种林分类型马尾松林枯落物层和土壤层水源涵养能力。结果显示,松材线虫入侵不同年限间马尾松林枯落物层有效拦蓄量为9.71~14.77 t·hm-2,松材线虫入侵不同年限间马尾松林土壤层有效蓄水量占比先下降后上升(表5)。入侵6 a 马尾松林枯落物层和土壤层的综合蓄水量明显低于其余3 种林分,表明入侵6 年林分马尾松林下水源涵养能力最差,入侵10 a 和16 a 马尾松林综合蓄水量明显回升,表明马尾松林下水源涵养能力逐渐恢复。

表5 松材线虫入侵不同年限间马尾松林综合蓄水量评估Table 5 Comprehensive water-holding capacity of Masson pines in different invasion years by pinewood nematode
4 讨论
4.1 松材线虫入侵对枯落物层水文效应的影响
枯落物层可为土壤表层提供保护,有效截持降水以减小降雨对土壤的直接冲刷和抑制土壤水分蒸发,阻延地表径流[20]。本研究发现松材线虫入侵6 a 的马尾松林枯落物层总厚度和总蓄积量均低于未入侵、入侵10 a 和入侵16 a 的马尾松林,表明松材线虫入侵在初期阶段对马尾松林枯落物蓄积量造成一定影响。其原因在于当马尾松林受到松材线虫入侵后,大量马尾松受害死亡,当地有关部门会对林内受害的马尾松进行择伐焚烧处理,使得林分密度大大降低[8],减少了枯落物的输入量和现存量。松材线虫入侵10 a 和入侵16 a 马尾松林枯落物总蓄积量高于未入侵马尾松林,这可能是受松材线虫入侵后的死亡马尾松被伐除焚烧,林内林窗变大,给其他阳性树种和灌草创造了生存与生活空间。林分植被有所恢复,群落的物种多样性增加,林分生物量和林内枯落物也将增加[9,21-22]。在样地调查中发现,松材线虫入侵10 a 和入侵16 a 的马尾松林内已出现以檵木、栎树和樟树等为主的更新层,群落物种多样性和林分生物量大大增加。而入侵16 a 蓄积量小于入侵10 a 蓄积量,可能是由于入侵16 a 林分密度小于入侵10 a 林分,枯落物输入量小于前者。
4种林分未分解层、半分解层枯落物随时间变化的持水过程相似,随浸水时间的增加,各植被类型枯落物持水量均随之增大,持水速率不断减小,与前人研究结果表现一致[11]。不同林分持水能力与速率有所不同,这与林内枯落物组成与分解程度有关[23]。在本研究样地中,松材线虫未入侵的林分是以马尾松为优势种的单优群落,枯落物以松针为主,而受松材线虫入侵16 a 的林分逐渐演变为针阔混交林,阔叶枯落物所占比例增加,可能导致了两种林分枯落物层持水能力的差异。未入侵林分枯落物未分解层单位重量持水量和持水速率最小,可能是由于针叶树种枯落物持水能力小于阔叶树种[24]。半-已分解层持水量和持水速率均大于未分解层,说明半-已分解层在枯落物层持水能力中发挥主要作用,可能是由于分解程度较大,枯落物比表面积相应更大,从而对水分的吸持能力更强,吸持水量更多[25],这与张轶超等研究结果一致[26]。松材线虫入侵6 a 的马尾松林枯落物的未分解层有效拦蓄量和最大持水量最小,与枯落物蓄积量规律一致,反映出松材线虫入侵初期马尾松林枯落物层水源涵养功能受损。随入侵年限增加,入侵10 a 和16 a 林分枯落物有效拦蓄量大于未入侵林分,一方面是由于林分植被恢复,蓄积量有所增加,另一方面,由于阔叶树种相比针叶拥有更大的表面积,能够有效拦蓄降雨、减少径流和增加入渗[27],表明枯落物层持水能力有所恢复。
4.2 松材线虫入侵对土壤层水文效应的影响
土壤层作为森林生态系统水分贮蓄的主要界面层,其持水能力是度量马尾松林生态水文功能维持与恢复的重要指标[27]。松材线虫入侵显著影响了马尾松林土壤物理指标和土壤储水能力,入侵6 a 的马尾松林土壤含水量、非毛管孔隙度显著低于其他林分,表明松材线虫入侵初期土壤水分物理性质发生较大改变,这与其他研究结论相似[28]。此外,每年人为择伐进行焚烧死树和林下清理踩踏使得土壤夯实板结,也可能是影响因素之一[29]。已有的研究证明,松材线虫入侵也会对土壤养分元素含量[30]、微生物群落[31]、根生物量[32]造成一定影响,并进一步降低土壤自我调节的能力[33]。与前人研究中入侵初期土壤总孔隙度、非毛管孔隙度、毛管孔隙度和毛管持水量等与土壤持水能力相关的物理指标均随着危害年限的增加而增加[8]的短期结果不同,本研究发现,湖北三峡地区马尾松林土壤总孔隙度、非毛管孔隙度、毛管孔隙度和毛管持水量等物理指标总体上随松材线虫入侵年限的增长呈先降低后上升趋势。说明松材线虫入侵导致马尾松林土壤储水能力短期内下降,但随植被逐渐恢复,土壤物理指标和持水能力不断上升。表明随入侵时间增加,林分土壤储水能力已有较好恢复。本研究中松材线虫入侵10 a 和16 a 马尾松林土壤有效蓄水量、毛管持水量和饱和蓄水量与未入侵林分差异不显著支持了上述推论。这可能是由于随着林分受松材线虫入侵持续影响,感病马尾松的死亡促进林分阔叶树生长,加速了森林的演替方向和演替程度[34-35],入侵林分群落结构和多样性与未入侵林分已有较大差异。本研究中,松材线虫不同入侵年限导致入侵10 a 和16 a 林分的树种组成[36-37]、林分密度[38]和演替程度[39]均发生较大程度变化,可能是使得土壤理化性质恢复的重大因素。而入侵10 a 林分土壤非毛管孔隙、容重、含水量等指标均优于入侵16 a 林分,可能是由于不同林分类型对土壤理化性质、储水能力的影响不同[40],各个土壤类型随着植被的演替而发生一定变化[41-42]。值得注意的是,不管是本研究还是其他研究,主要评估了松材线虫入侵对森林表层土土壤理化性质及其相应功能的影响,而对于深层土壤相应研究和林分长期持续性观测还十分缺乏。因此,当考虑深层土壤的相应指标和长期研究后,土壤水源涵养功能是否随着松材线虫入侵年限增加而恢复或减弱还需要做进一步的研究。
4.3 松材线虫入侵对马尾松林水源涵养能力的影响
综合来看,松材线虫入侵初期对马尾松林枯落物蓄积量产生一定影响,随年限增加,蓄积量逐渐恢复。4 种林分枯落物持水能力变化不显著,表明松材线虫未对马尾松林枯落物层造成较大影响。枯落物层和土壤层综合蓄水量的结果表明,马尾松林土壤水源涵养能力随着松材线虫入侵年限的增加先降低后上升。马尾松林综合蓄水量由土壤有效蓄水量主导,而松材线虫入侵短期内显著影响马尾松林土壤物理性质及其与土壤持水能力相关的其他物理指标,随年限增加逐渐恢复。综上所述,应加强三峡地区马尾松林松材线虫入侵的防治力度,并在此过程中应减轻人为干扰对林分和土壤结构的影响,鼓励以天敌防治、改善林分结构等生态调控策略为主[43-45],以维持和尽快恢复其水土保持和水源涵养功能。
5 结论
(1)松材线虫入侵对马尾松林枯落物蓄积量、最大持水量和有效拦蓄量均无显著影响。松材线虫入侵6 a 的马尾松林枯落物层厚度、蓄积量和有效拦蓄量最小。总体上,随入侵年限增加,马尾松林枯落物蓄积量、持水能力有所恢复。
(2)松材线虫入侵短期内显著影响马尾松林土壤物理性质、有效蓄水量、毛管持水量和饱和蓄水量。随入侵年限增加,马尾松林土壤有效蓄水量和饱和蓄水量呈逐渐恢复趋势。
(3)松材线虫入侵短期内显著明显减弱了湖北三峡地区马尾松林枯落物层和土壤层持水能力,随年限增加逐渐恢复。因此,应加强松材线虫入侵的防控力度,以维持三峡地区马尾松林水源涵养功能。
致谢:感谢湖北秭归三峡库区森林生态系统定位观测研究站、夷陵区林草局和秭归县林草局对本研究的支持,感谢森林生态环境与自然保护研究所王兵研究员、牛香研究员、曾立雄研究员、黄志霖研究员、雷蕾副研究员的指导与帮助。
猜你喜欢
新疆地质(2024年1期)2024-04-17
水资源开发与管理(2019年11期)2019-12-02
绿色中国(2019年13期)2019-11-26
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
河北林业科技(2016年5期)2016-11-08
河北林业科技(2016年3期)2016-10-27
干旱地区农业研究(2016年2期)2016-04-25
防护林科技(2015年12期)2016-01-13
河北林业科技(2015年3期)2015-12-17
