
2016—2021年长江口海域营养盐时空变化特征及其影响因素研究
2024-02-21 09:55何柄震卢士强李立群程全国
环境科学研究 2024年2期
何柄震,王 艳,王 彪,卢士强,雷 坤,李立群,程全国
1. 沈阳大学,区域污染环境生态修复教育部重点实验室,辽宁 沈阳 110044
2. 中国环境科学研究院流域水环境污染综合治理研究中心,北京 100012
3. 上海市环境科学研究院,上海 200233
海洋水体中的氮、磷、硅等营养盐物质不仅是海洋中浮游植物生长、发育和繁殖所必需的要素,同时还是海洋初级生产力和生态系统中食物链循环的物质基础[1],营养盐的可调节性与可利用性是浮游植物生长发育和繁殖、海洋物种丰富度以及生物数量的关键因素[2]. 因此,研究氮、磷、硅等营养盐的时空分布变化规律对于河口地区和海洋的生态环境变化具有重要的科学意义.
长江口是长江河水和东海海水进行物质运输与交换的重要通道[3-4]. 河流在由陆源向海洋输送营养物质的过程中起着重要作用[5]. 长江河水中含有丰富的氮、磷等营养物质,它们通过长江水流输送到长江口海域,故长江的陆源输入是长江口海域营养物质的重要来源之一. 长江口海域营养盐的浓度变化和空间分布特征受到长江冲淡水的输入与台湾暖流、苏北沿岸流等洋流交汇混合效应的影响[6-7]. 近年来,由于陆地径流、大气沉降以及人类活动等多种因素的影响,污水排放、大型水文工程设施的建设、海水养殖以及颗粒物沉降等因素对河口和近岸海域造成了较强的污染负荷,这也是导致长江口海域出现富营养化等环境问题的主要原因[8]. 因此,探究长江口海域氮、磷、硅等营养盐的时空变化情况及影响因素,对于维护该海域的生态平衡和生态系统健康状况至关重要.
由于长江口地区具有独特的地理特征,已有大量学者对长江口海域做了定性与定量研究,包括营养盐变化特征[9-11]、营养盐结构及限制因子[12-15]、营养盐迁移转化过程[16-18]、水体富营养化[19-21]、水质评价方法[22-24]、水质预测模型[25-27]以及通量研究与估算模型[28-31]等. 但大多数研究皆是基于某一特定航次或部分年份、季节的监测数据. 本文利用长时间序列的多年、多季节长江口海域营养盐实测数据,从分区的角度,分析了长江口在“十三五”期间(2016-2020年)和“十四五”初期(2021年)的营养盐通量变化情况、营养盐时空分布特征、营养盐结构特征以及主要影响因素,并探讨了长江口海域主要潜在限制因子及其与Chla浓度的耦合关系,以期为长江口海域生态环境研究提供依据与参考.
1 研究区域与研究方法
1.1 研究区域
长江口位于东海与黄海的交界处[32],是一个中等潮汐、水沙丰富、规律分汊的三角洲型河口. 长江口的自然环境受到长江径流、海洋潮汐和陆海交汇等多个因素的共同影响,同时呈现出淡水、咸淡水和海水3种不同的特性. 在长江口地区,陆地和海洋的物质相互交融,咸淡水混合,同时径流和潮汐相互影响[33-34],从而产生了多种独特并且复杂的物理、化学、生物反应及沉积过程. 因此,长江口的自然条件、物种资源和生态环境的复杂多样性使其成为一个独特的生态系统. 本研究的调查区域为长江口海域,其地理范围在30°30'N~31°50'N、121°05'E~123°15'E之间.
1.2 数据来源
本研究收集了2016-2021年每年春季(4-5月)、夏季(7-8月)、秋季(10-11月)上海市近岸海域生态环境监测数据,指标包括氨氮(NH3-N)、硝酸盐(NO3-N)、亚硝酸盐(NO2-N)、磷酸盐(PO4-P)、硅酸盐(SiO3-Si). 入海断面朝阳农场站NH3-N、NO3-N、TN及TP的浓度月度数据主要来源于生态环境部长江水系的国控断面水质监测数据. 徐六泾水文站的每月径流量数据来源于2016-2017年的《长江水文年鉴》和水利部长江水利委员会编纂的2018-2021年《长江流域重要控制断面水资源监测通报》. 长江口海域调查站位分布示意如图1所示.

图1 长江口海域调查站位分布示意Fig.1 Distribution of survey stations in the Yangtze River Estuary
1.3 研究方法
1.3.1 数据处理方法
由于监测年份不同,站位位置、日期和时间均存在一定差异,因此,首先需要将数据进行预处理[35],将2016-2021年具有相同名称的调查站位统一到相同的经纬度,数据缺失部分以及异常值采取剔除处理,并按照时间序列对营养盐数据进行整合. 由于长江口海域盐度的空间分布表现为自长江口内向口外逐渐升高的趋势[36],并且长江口海域生物群落分布特征与盐度具有明显的契合关系[37],故根据2011-2021年长江口海域盐度(S)数据,利用克里金空间插值法计算出S=5‰和S=25‰的盐度等值线,并且将长江口海域分为3个区域,即口门区(S<5‰)、河口区(5‰<S<25‰)和海水区(S>25‰).
本研究采用SPSS 26软件完成显著性检验和相关性分析,利用Excel软件完成所有基本统计分析,用Origin 2021软件完成折线图、柱状图、散点图以及3D图的绘制,用ArcGIS 10.2软件绘制所有空间分布图.
1.3.2 通量计算方法
通量计算采用长江入海断面的实测水质数据以及距离入海断面距离最近水文站的流量数据来计算入海通量,即选择朝阳农场断面监测站点的水质数据以及徐六泾水文站的径流量数据. 目前国内外学者已对通量计算方法及其产生的各类误差修正进行了系统总结[38-41],分别采用式(1)和式(2)进行河流断面月入海通量(Wmi)和年入海通量(Wai)的计算:
式中:Wmi为河流断面月入海通量,104t;Wai为河流断面年入海通量,104t;Ci为河流断面某营养盐某年i月的平均浓度,mg/L;Qi为该河流断面某年i月的径流量,108m3.
2 结果与讨论
2.1 径流量与通量变化
2.1.1 长江入海径流量变化
徐六泾水文站径流量数据(见图2)显示,2016-2021年长江的入海径流经历了不同的水文年份,2016-2017年和2020-2021年为丰水年,年径流量较大并保持在多年平均值以上,2018-2019年为枯水年. 从月径流量变化可以看出,2016-2021年入海径流量的月度变化整体上表现为先升高再降低的趋势,径流量存在明显的高值区,长江口入海水量主要集中在每年5-10月的汛期间,长江径流的水量占全年总量的67.05%,相比之下,在11月-翌年4月的非汛期间,长江的径流量相对较小,仅占全年总量的32.95%.
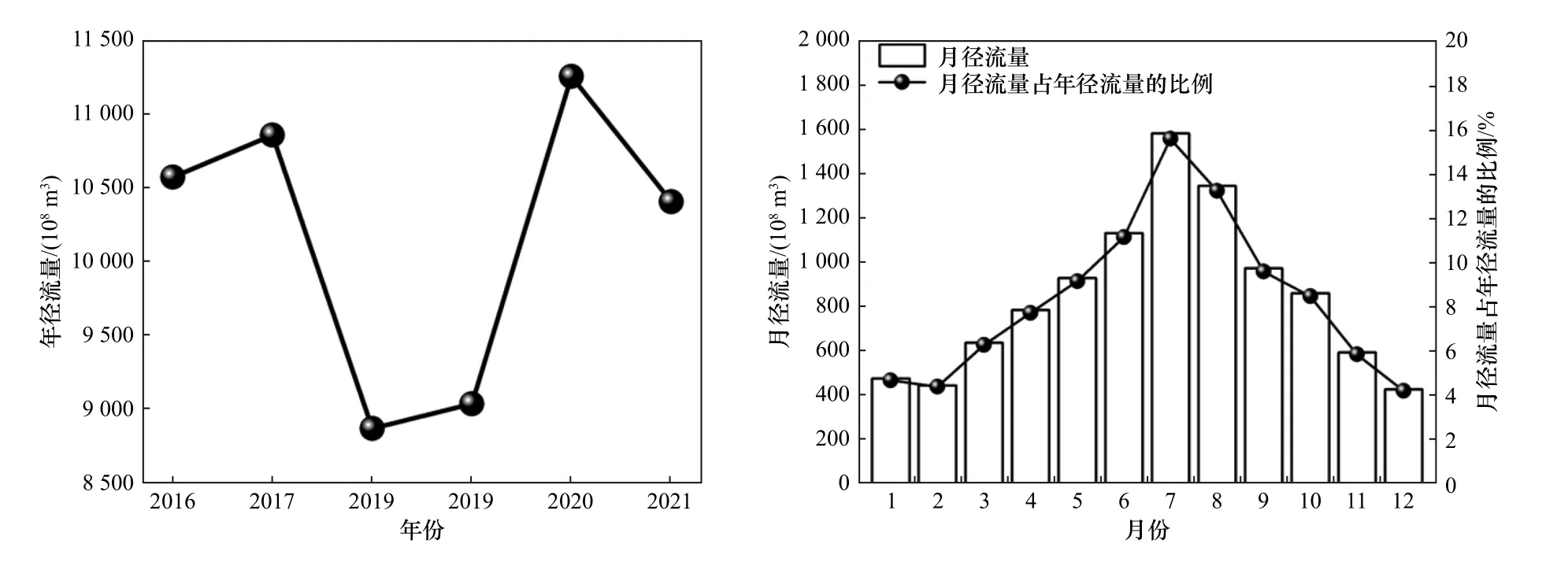
图2 徐六泾水文站年径流量和多年平均月径流量的变化Fig.2 Variation of annual runoff and multi-year average monthly runoff at Xu Liujing Hydrological Station
2.1.2 营养盐入海通量变化
按照式(1)(2)计算得到2016-2021年长江水体中各营养盐年入海通量(见表1)和月入海通量(见图3). 为了考察各营养盐入海通量的年际变化趋势,通过计算得到NH3-N、NO3-N、TN、TP年入海通量的年际轶相关系数(Rs),分别为-0.916(p<0.01)、-0.543(p<0.05)、-0.829(p<0.01)和-0.824(p<0.01),这表明这几种物质的年入海通量整体上均呈现下降趋势. 由图3可知,NO3-N、TN、TP月入海通量与长江径流量变化趋势类似,均呈现出丰水期高、枯水期低的变化特征,与此同时也说明NO3-N、TN、TP入海通量主要受到长江径流作用的影响[42].

表1 2016—2021年长江口海域营养盐入海通量Table 1 Annual nutrients flux into the Yangtze River Estuary from 2016 to 2021

图3 2016—2021年长江口海域营养盐多年平均月入海通量变化Fig.3 Monthly nutrients flux into the Yangtze River Estuary from 2016 to 2021
2.2 营养盐浓度时空变化特征
2.2.1 DIN浓度时空变化特征
海水中的DIN包括NO3-N、NO2-N及NH3-N,它们在一定条件下可以进行相互转化,同时在海洋生物食物链中发挥着非常重要的作用[43]. 在长江向河口输送的DIN中,NO3-N占绝对优势,占DIN总含量的95%以上. NO2-N作为硝化和反硝化过程中的中间产物,其状态不稳定且含量很低. NH3-N主要来源于人类活动,如工业、农业及生活污水的排放.
2016-2021年长江口海域DIN浓度的年际及季节分布特征如图4所示. 由图4(a)可知,2016-2021年长江口海域DIN浓度的年均值在0.93~1.02 mg/L之间,2018年出现峰值(1.02 mg/L),2017年、2021年DIN年均浓度最低(均为0.93 mg/L). 6年间,DIN浓度呈波动下降趋势,这与TN入海通量的下降有关.由图4(b)可知,2016-2021年长江口海域DIN浓度季节分布特征主要表现为春季>夏季>秋季.

图4 2016—2021年长江口海域DIN浓度年际及季节分布特征Fig.4 The interannual and seasonal distribution of DIN in the Yangtze River Estuary from 2016 to 2021
从不同空间分区来看,DIN浓度在河口内出现最高值,从入海口到远海方向,浓度呈现逐渐降低的变化规律,低浓度区域主要集中在靠近外海的一侧,即口门区>河口区>海水区,DIN浓度与海水盐度(S)呈显著负相关(R=-0.985,P<0.01),说明DIN的分布主要受长江冲淡水影响,此特征与Wu等[44]研究的结果一致,同时DIN在河口的行为具有良好的保守性[45],即海洋中存在一些物质,它们具有相对稳定性,不易因化学过程而改变浓度. DIN的空间分布特点在年际间均存在差异,这也与长江入海径流量的大小以及外海水团的影响高度相关[46-47].
Chla是表征浮游植物生物量的关键指标[48]. 因此,通过计算DIN浓度与Chla浓度的相关性,探究浮游植物对营养盐的影响. 在口门区,NO3-N入海通量与DIN浓度之间存在较强的正相关关系(R=0.675,P<0.05),并且DIN浓度与Chla浓度的相关关系不显著(见表2),DIN浓度主要受长江陆源输入的影响.在河口区,NO3-N入海通量与DIN浓度存在较强的负相关关系(R=-0.779,P<0.05),推测DIN浓度受浮游植物生长繁殖和长江陆源输入共同作用的影响.从DIN浓度与Chla浓度之间的相关关系可以看出,尤其在夏季,DIN浓度与Chla浓度呈显著负相关(R=-0.583,P<0.01). 这表明浮游植物的生长和吸收作用是导致DIN浓度下降的一个重要影响因素[49].
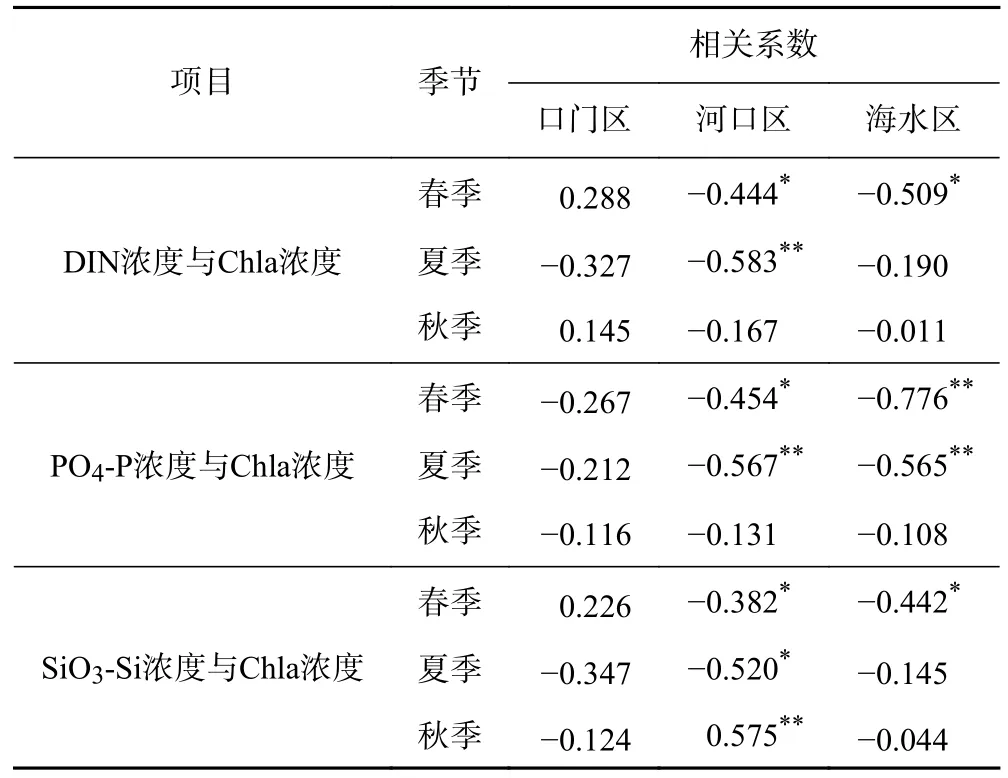
表2 长江口海域不同分区营养盐浓度与Chla浓度的相关关系Table 2 Correlation between concentration of nutrients and Chla in different zones in the Yangtze River Estuary
2.2.2 PO4-P浓度时空变化特征
2016-2021年长江口海域PO4-P浓度的年际及季节分布特征如图5所示. 由图5(a)可知,长江口海域PO4-P浓度总体上呈波动下降的趋势,可能是三峡水库保留了大量的颗粒磷,导致TP的入海通量减少,这也是近几年PO4-P浓度减少的原因[50]. 2016-2021年长江口海域PO4-P浓度的年均值在0.029~0.039 mg/L之间,2016年和2019年出现峰值(均为0.039 mg/L),2021年PO4-P的年均浓度最低(0.029 mg/L).由图5(b)可知,2016-2021年长江口海域PO4-P浓度的季节分布特征主要表现为秋季>夏季>春季.

图5 2016—2021年长江口海域PO4-P浓度年际及季节分布特征Fig.5 The interannual and seasonal distribution of PO4-P in the Yangtze River Estuary from 2016 to 2021
从不同的空间分区来看,PO4-P浓度均表现为河口区>口门区>海水区,其浓度高值区并不出现在口门区,PO4-P作为一种具有非保守性的营养物质,其浓度的变化也存在多种影响因素. 一方面是通过浮游植物的吸收和摄取作用被消耗、转移,另一方面则是受到泥沙等物质的吸附与释放的影响[51]. 此外,黑潮流经东海也成为长江口海域PO4-P的重要来源之一[52]. 通过分析PO4-P浓度与盐度(S)之间的相关关系发现,二者呈显著负相关(R=-0.771,P<0.01),说明PO4-P浓度同样也受到长江冲淡水作用的影响.
虽然PO4-P浓度也大致呈现出由近岸向远海递减的空间分布特征,但与DIN和SiO3-Si相比,其变化规律并不显著. 河口区内的PO4-P浓度整体上要高于口门区和海水区,PO4-P浓度的高值区通常位于最大浊度带附近,与此同时,TSM浓度在一定程度上也影响PO4-P浓度的空间分布情况. 在最大浊度带区域附近,相对于其他营养盐而言,PO4-P更容易与TSM发生吸附或释放作用[53]. 由图6可知,TSM浓度与PO4-P浓度呈显著正相关,且PO4-P浓度的变化梯度较小,浓度相对比较稳定,这说明通过一种缓冲机制使PO4-P从TSM中释放出来,而这种缓冲机制的主要作用就是可以保持海水中PO4-P的浓度基本无明显变化[54]. 悬浮物质可以在富含活性磷酸盐的淡水水体中吸附PO4-P,并将其释放回到PO4-P浓度较低的咸淡水交汇区水体中,这就使得河口水域的PO4-P浓度在一定范围内始终保持稳定[55],长江向河口输入的大量泥沙,受到风、浪、潮汐和海流等多种水动力因素的影响,经过吸附-解析过程,促进了在水体、悬浮物质和沉积物之间的物质交换,这有利于维持水中PO4-P浓度的平衡[56]. 在口门区,入海径流量与PO4-P浓度存在一定的正相关关系(R=0.544,P<0.05),说明径流输入能够在一定程度上增加口门区PO4-P的浓度. 在河口区与海水区,入海径流量与PO4-P浓度之间存在较强的负相关关系(R=-0.465,P<0.05),并且在春夏两季PO4-P浓度与Chla浓度呈显著负相关(见表2),其原因可能是浮游植物的生长摄取决定了PO4-P浓度的变化,并削弱了入海径流量对PO4-P浓度的影响[57].

图6 长江口海域PO4-P浓度与TSM浓度相关性分析Fig.6 Correlation between concentration of PO4-P and TSM in the Yangtze River Estuary
2.2.3 SiO3-Si浓度时空变化特征
SiO3-Si是硅藻生长必需的营养物质,其分布不仅受海水的稀释与扩散影响,也受一些海洋生物活动和悬浮体吸附的影响[58]. 由图7(a)可知,2016-2021年SiO3-Si浓度整体上呈现波动上升趋势,年均值的范围在1.66~2.27 mg/L之间,最高值(2.27 mg/L)出现在2020年,最低值(1.66 mg/L)出现在2017年. 从不同的季节变化〔见图7(a)〕来看,2016-2021年长江口海域SiO3-Si浓度变化主要表现为秋季>夏季>春季.
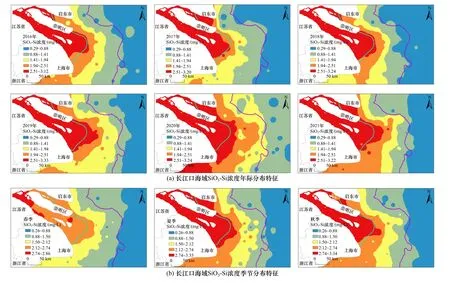
图7 2016—2021年长江口海域SiO3-Si浓度年际及季节分布特征Fig.7 The interannual and seasonal variation of distribution of SiO3-Si in the Yangtze River Estuary from 2016 to 2021
从空间分布上来看,SiO3-Si浓度与DIN浓度分布情况类似,呈现出口门区>河口区>海水区的分布特征,即SiO3-Si浓度整体分布从近岸向外海逐渐减小. 通过分析SiO3-Si浓度和盐度(S)的相关性发现,长江口海域SiO3-Si浓度与盐度(S)呈显著负相关(R=-0.991,P<0.01),这说明SiO3-Si浓度及其空间分布也受到长江冲淡水的显著影响,并且每年长江径流向长江口海域输送大量SiO3-Si,为某些浮游植物(主要是硅藻)的生长繁殖提供了充裕的营养元素.
在口门区,SiO3-Si浓度与入海径流量呈正相关(R=0.768,P<0.05),说明SiO3-Si浓度主要受长江径流的影响. 在河口区,春季SiO3-Si浓度与Chla浓度呈负相关(R=-0.382,P<0.05),夏季二者的负相关性变得更加显著(R=-0.520,P<0.05),说明旺盛的浮游植物生长会降低长江口海域的SiO3-Si浓度,秋季二者主要呈正相关(R=0.575,P<0.01),其原因可能是浮游植物群落中的甲藻比例上升,硅藻比例下降,从而抑制了硅藻对SiO3-Si的摄取[59-60].
2.3 营养盐结构变化及其影响因素
长江口海域营养盐结构的年际变化如图8所示.由图8可见:N/P的平均值分布在59.33~74.21之间,其变化趋势表现为波动上升的状态;Si/N的平均值在0.96~1.21之间,其变化趋势为先降低再升高,最小值出现在2018年,Si/P的平均值介于57.97~80.37之间,其变化趋势与Si/N类似,2017年出现最小值,2017-2021年呈逐渐上升趋势.
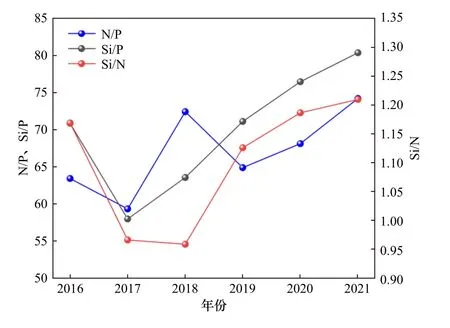
图8 2016—2021年长江口海域营养盐结构的年际变化情况Fig.8 The interannual variation of nutrient structure in the Yangtze River Estuary from 2016 to 2021
长江每年都会向河口地区输送大量的营养盐等物质,通常情况下,伴随着营养盐输入的同时,营养盐之间的摩尔比也会随之发生改变[61]. 通过长江口海域不同分区营养盐的摩尔比(见表3)可以看出,口门区、河口区、海水区春夏秋季N/P平均值分别为(85.66±12.37)(56.30±16.95)(63.39±37.66),Si/P平均值分别为(83.87±10.12)(56.12±12.66)(66.60±26.24),均远大于16∶1,而Si/N则均接近1[62]. 根据N/P、Si/N、Si/P的散点图数据(见图9),发现长江口海域营养盐结构具有分布不均匀的特点[63]. 适宜的N/P、Si/P和Si/N是浮游植物生长发育和繁殖的基本条件. 相反,如果缺乏某种营养元素,将会限制浮游植物的生长;某种物质浓度过高同样也会对浮游生物的群落结构产生重要的影响[64]. 当N/P>22、Si/P>22时为磷限制,当Si/P<10、Si/N<1时为硅限制,当N/P<10、Si/N>1时为氮限制[65],根据该判定标准显示,磷是长江口海域浮游植物生长和繁殖的主要潜在限制因子.
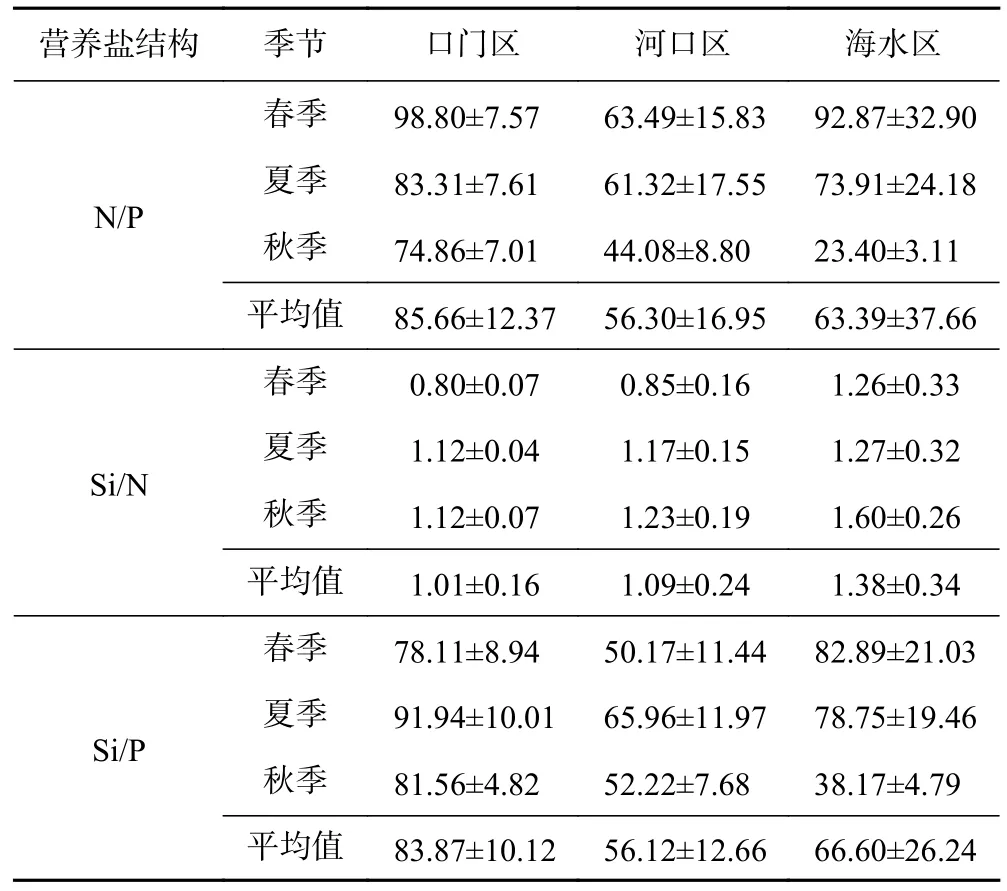
表3 长江口海域不同分区春夏秋季营养盐摩尔比Table 3 The nutrients structure in different zones during spring,summer and autumn in the Yangtze River Estuary
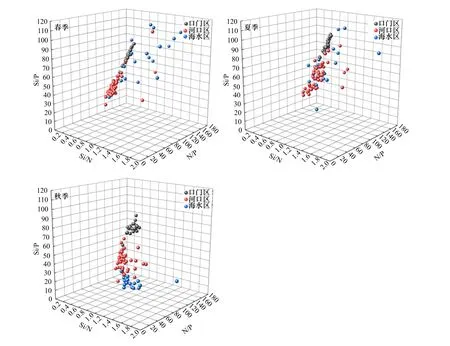
图9 长江口海域不同分区春夏秋季N/P、Si/N、Si/P的散点分布Fig.9 The scatter distribution of N/P, Si/N, Si/P in different zones during spring, summer,and autumn in the Yangtze River Estuary
据长江口海域营养盐结构空间分布(见图10)显示,在口门区,由于盐度较低、水质浑浊,DIN和SiO3-Si浓度高,而PO4-P浓度较低,因此表现出N/P和Si/P值较高. N/P、Si/P在河口区较低,特别是位于最大浑浊带地区附近,由于该区域透明度相对较低,浮游植物生长繁殖活动能力较弱,大量的磷物质从TSM中释放到周围水体,因而导致N/P与Si/P较低[66]. 为探讨浮游植物与营养盐结构的耦合关系,通过筛选2016-2021年长江口海域高浓度的Chla(>10 μg/L)以及对应的N/P、Si/P、Si/N(见图11)发现,在Chla浓度大于10 μg/L的区域普遍存在高N/P的特点[67],而Si/N的低值区、N/P的高值区与之对应,这可能与硅藻爆发产生的赤潮密切相关[68].高浓度的Chla所在的站位,磷是浮游植物的主要潜在限制因子. 由于生物的吸收作用会使磷的限制程度加剧[69],所以lg(N/P)和lg(Si/P)普遍偏大. 这说明浮游植物对长江口海域营养盐结构具有重要的影响.

图10 长江口海域营养盐结构空间分布Fig.10 Space distribution of nutrients structure in the Yangtze River Estuary
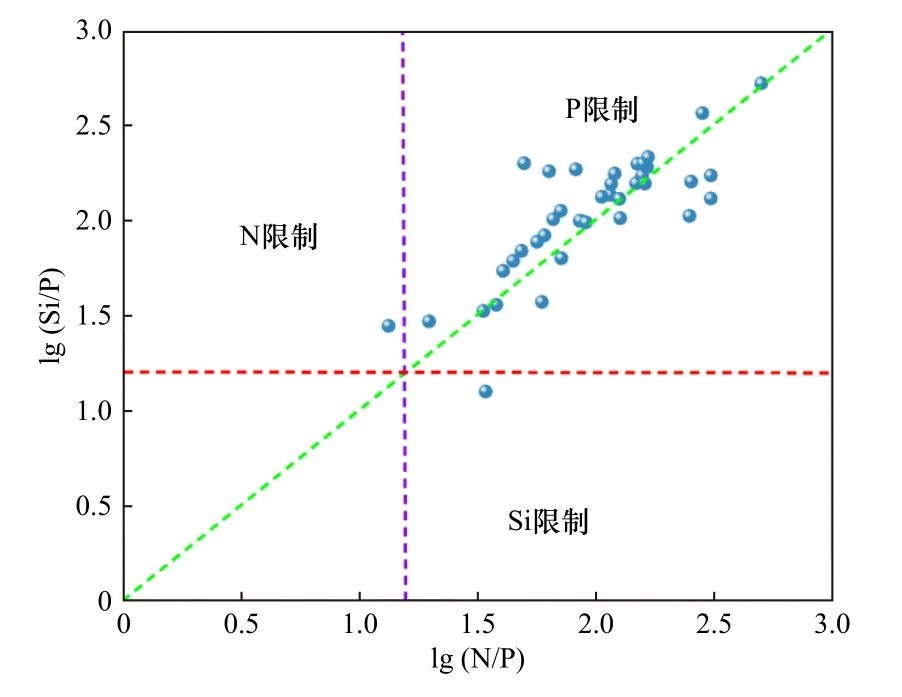
图11 长江口海域高浓度Chla(>10 μg/L)对应的营养盐比例散点分布Fig.11 Scattered distribution of nutrient ratios corresponding to high concentrations of Chla(>10 μg/L) in the Yangtze River Estuary
3 结论
a) 2016-2021年长江口海域DIN、PO4-P浓度呈波动下降趋势,SiO3-Si则呈波动上升趋势. DIN浓度主要表现为春季>夏季>秋季,而PO4-P和SiO3-Si浓度变化则呈现出秋季>夏季>春季的趋势,各种营养盐呈现明显的季节性变化特点.
b) 空间上,DIN、SiO3-Si浓度都是由近岸向远海逐渐递减的趋势,均呈现出口门区>河口区>海水区的分布特征;PO4-P浓度则表现为河口区>口门区>海水区. DIN、PO4-P、SiO3-Si浓度都受到长江冲淡水的影响. PO4-P在长江口受到一种缓冲机制的影响,其高值区通常位于最大浊度带附近,TSM浓度在一定程度上也影响PO4-P的分布情况. Chla浓度是表征浮游植物生物量的关键指标,DIN、PO4-P和SiO3-Si的浓度都会受到Chla浓度的影响,即浮游植物的生长繁殖是营养盐浓度变化的重要影响因素之一.
c) 长江口海域营养盐结构具有分布不均衡的特点,N/P、Si/P均远大于16:1,Si/N则均接近1:1. 磷是长江口海域浮游植物生长和繁殖的主要潜在限制因子. N/P和Si/P表现为口门区较高,河口区较低的特点. Chla浓度的相对高值区域普遍都是磷限制,生物的吸收作用会使磷的限制程度加剧,表现出Chla的浓度越高,N/P和Si/P越大,说明浮游植物对长江口海域营养盐结构具有重要的影响.
猜你喜欢
金桥(2021年11期)2021-11-20
少儿美术(快乐历史地理)(2020年5期)2020-09-11
水道港口(2017年5期)2017-11-22
中学生数理化·八年级物理人教版(2017年5期)2017-11-13
水科学与工程技术(2016年2期)2016-07-10
海洋渔业(2016年6期)2016-04-16
海洋渔业(2016年6期)2016-04-16
中国水利(2015年20期)2015-02-01
华东师范大学学报(自然科学版)(2014年3期)2014-03-11
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
