
水菜花种群潜在生境选择与空间格局预测
2024-02-02 06:10吴庭天雷金睿陈宗铸陈小花李苑菱
热带亚热带植物学报 2024年1期
吴庭天, 雷金睿, 陈宗铸, 陈小花, 李苑菱
水菜花种群潜在生境选择与空间格局预测
吴庭天, 雷金睿*, 陈宗铸, 陈小花, 李苑菱
(海南省林业科学研究院(海南省红树林研究院),海南省热带林业资源监测与应用重点实验室,海口市湿地保护工程技术研究开发中心,海口 571100)
国家二级保护野生植物水菜花(),喜生于清洁的水环境中,对环境变化极为敏感,是检验湿地环境及气候变化的关键指示物种之一,在我国仅零星分布于海南北部的火山熔岩湿地区,生存状况不容乐观。研究水菜花种群潜在生境选择及其空间格局演变,有利于加强濒危物种保护保育及湿地生态系统修复、管理。该研究基于GIS平台和MaxEnt模型,结合气候、地形和土壤因子,探究水菜花种群环境限制因子及其在气候变化背景下潜在适宜生境的演变格局。结果表明,水菜花种群对温差与降水量变化敏感,等温性、最冷季度降水量、土壤类型和年均降水量对水菜花种群分布影响显著;全新世中期-当前-2070年气候变化背景下,水菜花适宜生境面积先减小后增大,分布重心呈西南-东北-西南转移格局;未来气候情景下,水菜花种群高度和中度适宜生境缩减,低适宜生境增加,南部地区将出现新增适宜生境,东北、西北及西南部适宜生境将发生消减。该研究从气候环境角度论证了水菜花种群的潜在生境选择及空间变化特征,可为濒危物种保护保育、湿地管理及其生物多样性维护工作提供参考和指导。
水菜花;气候变化;潜在生境;环境因子
水菜花(),隶属于水鳖科(Hydro- charitaceae)水车前属,是一年生或多年生水生草本植物,在2021年9月最新公布的《国家重点保护野生植物名录》中列为国家二级保护野生植物,主要分布于中国、泰国、缅甸和柬埔寨等地区的淡水沟渠及池塘中。水菜花的生长对水质要求极高,喜生于清洁的水环境中,常成单株或几株聚生,野外分布数量较少,在我国仅零星分布于海南北部的海口、文昌、澄迈等地的火山熔岩湿地区[1]。近年来,随着湿地面积减少、水体污染加剧以及气候变化的影响,水菜花适宜生境正被逐渐压缩、种群数量急剧减少,生存状态日渐濒危。目前,有关水菜花的研究极少,1984年赵佐成等[2]对华南地区淡水水鳖科植物的生态特征和群落学进行观察,详细描述了水菜花的生活习性、生长环境、群落组成等群落学特征,此后主要围绕水菜花种群动态[3]、核型分析[4]、基因测定[5–6]和叶片光合效率[7–8]等开展了研究。
随着水菜花种群及生境的逐渐减小,对其适宜生境开展研究尤为重要。而传统仅依靠人力进行野外实地数据采集的生境调查方法,过于耗力费时, 不适宜做大尺度的生境状况评估,运用模型拟合物种分布与环境之间的相互关系,探究物种空间分布情况,已成为现代科学研究的常用手段。在所有模型中,MaxEnt模型可以在样本数量小、物种存在信息不完整以及少量位点偏差的情况下表现出良好的模拟预测结果,加上其简单、友好的操作界面,近年来已被广泛应用于生态、生物、地理等多学科的科学探究中[9–10]。本研究基于实地采集的水菜花真实分布点数据,结合MaxEnt模型,运用物种分布与地形和自然气候环境间的紧密关联,拟合水菜花种群适宜生境分布,并结合气候变化情景预测水菜花种群历史和未来的适宜生境分布特征。旨在明晰以下2个问题:(1) 当前气候条件下,水菜花适宜生境在海南的分布情况,并明确主要限制因子; (2) 通过模拟水菜花种群在历史和未来气候条件下的分布情况,获悉水菜花适宜生境在时间维度上的变化特征。通过以上研究,明确水菜花种群在气候变化背景下的地理分布特征及其主要限制因子,为日渐萎缩的水菜花种群数量及栖息地恢复提供科学参考。
1 材料和方法
1.1 研究区概况
水菜花种群喜生于淡水溪流、池塘中,热量和pH值是影响水菜花生存和分布的主要制约因子[2], 根据野外实测记录的分布点可知,水菜花种群主要分布在海南北部的海口、文昌、澄迈周边的火山熔岩湿地区,该地区属热带海洋性季风气候,年降水量1 600~2 000 mm,pH值5~8[11],年平均气温23 ℃~ 24 ℃,年平均最低温度8 ℃~9 ℃[12–13],地形地貌为火山喷发活动后河溪、湖泊、田洋、水库、池塘、森林沼泽等组成的火山台地和平原[14],良好的水热条件十分适宜水菜花生长。
1.2 数据获取
水菜花分布数据均为实地调查所得,调查地点主要集中在海南省海口市、澄迈县、定安县,共采集分布点45个,为避免因分布点位距离过近造成模型的过度拟合,本研究在每个1 km×1 km的网格中仅保留一个分布点,最终保留20个有效分布点作为模型拟合的基础数据(图1)。
本研究使用的数据主要包括气象、地形、土壤3大类21个变量。气象数据包括气温和降水2部分的19个变量,分全新世中期Mid-Holocene (6 000年以前)、当前(1970—2000年)和未来(2061—2080年) 3个时期,均来自世界气候数据库(https://www. worldclim.org/),空间分辨率为1 km;其中,未来气候数据选用BCC-CSM2-MR气候系统模式中的SSP1-2.6、SSP2-4.5、SSP3-7.0和SSP5-8.5不同社会经济假设驱动的4种共享情景。地形变量主要引用中国科学院计算机网络信息中心(http://www. gscloud.cn)的DEM数据。土壤类型数据来源于中国科学院资源环境科学与数据中心(www.resdc.cn)。

图1 水菜花调查点位分布图
环境因子间大多存在一定的关联性,为避免模型拟合过程中发生数据冗余和过度拟合,本研究在参考相关研究文献的基础上[15–16],对全部21个变量进行相关分析,当2个因子间的相关性大于0.85时,去除贡献率较小的,最后选取14项环境变量用于最终的模型拟合(表1)。
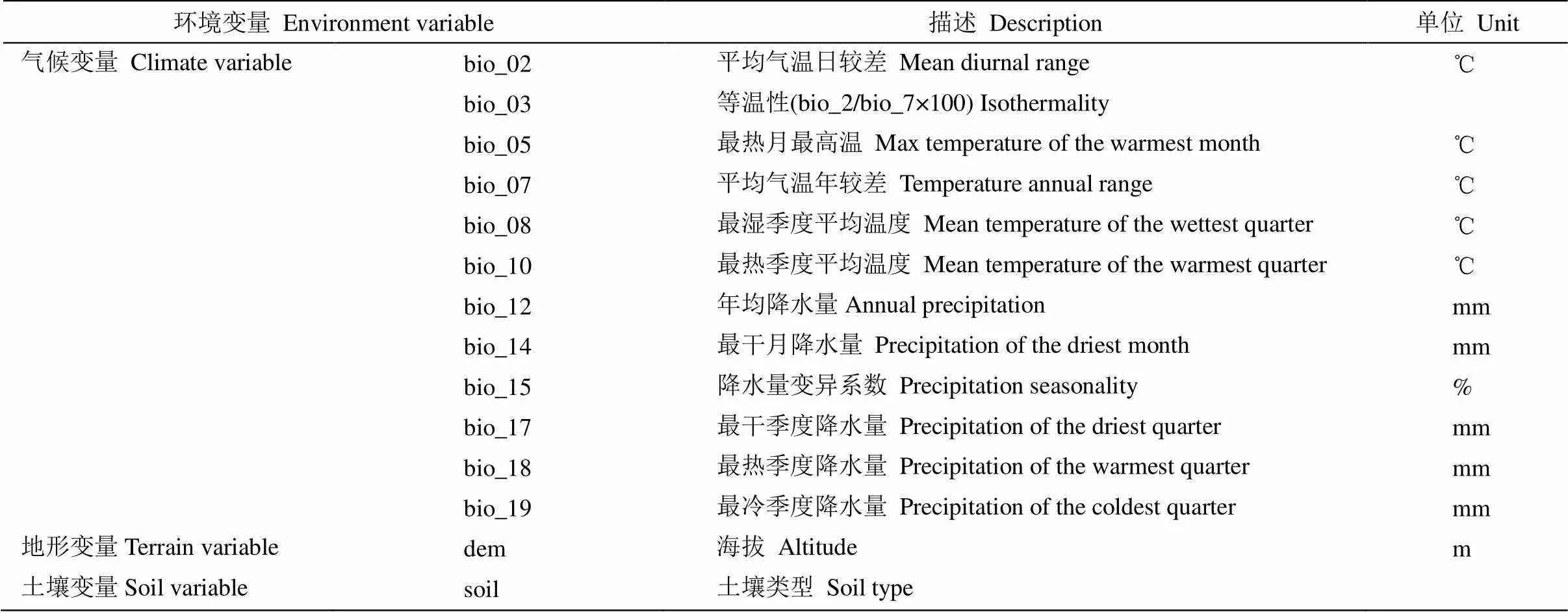
表1 环境变量
1.3 数据准备
(1) 将所有环境因子数据裁剪为同样大小范围并重新投影到相同坐标;(2) 将整理后的物种分布数据转成csv文件;(3) 通过重采样将所有变量数据转成栅格数据,结合空间插值及重采样方法将分辨率统一至30 m;(4) 将所有变量栅格数据转成asc格式文件。
1.4 模型构建
在MaxEnt软件中相继载入经过预处理的濒危物种分布数据和环境变量数据。多次拟合调整后得出,随机测试比例设定为25%,正则化乘数设置为1.2时,结果最贴近物种实际分布特征,同时采用Jacknife检验环境因子重要性,模型运行精度使用ROC曲线(受试者工作特征曲线)下面积(AUC)进行评价,值越大模型预测效果越好[16]。评价标准分一般、好和非常好3级,对应的AUC值区间分别为:0.7~0.8、0.8~0.9和0.9~1.0[17–18]。设置20次bootstrap重复,取最终平均值,其他参数保持默认设置。模拟结果以概率的形式反映物种在空间上存在的可能性大小,取多次重复后的最低培训存在值作为划分物种适宜生境的阈值[19],该值是模型拟合出的物种存在的最低临界值。最后在GIS平台下通过自然断点法[16,20]对水菜花生境拟合结果进行适生等级划分。
2 结果和分析
2.1 影响水菜花分布的环境因子
基于MaxEnt软件20次重复模拟的结果表明, ROC曲线综合AUC值为0.993,模型拟合预测效果表现良好,可靠性高(图2)。
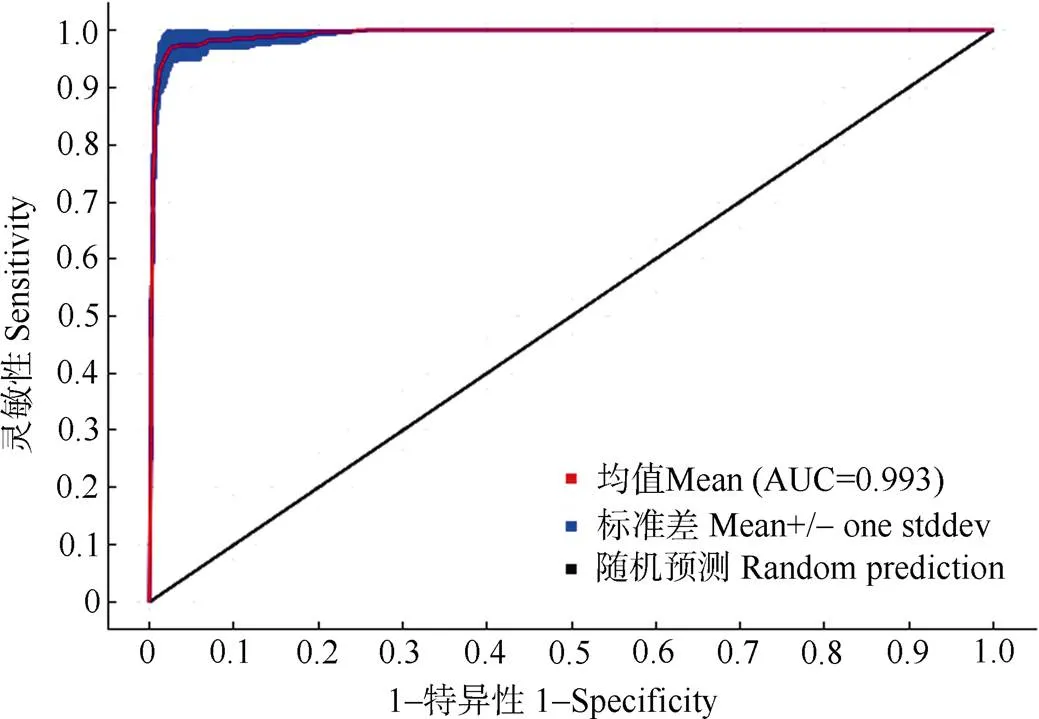
图2 模型拟合的受试者工作特征曲线
MaxEnt模型拟合结果表明(图3),等温性(bio_03, 22.6%)、最冷季度降水量(bio19, 21.4%)、土壤类型(16.6%)和年均降水量(bio_12, 14.1%)对拟合结果贡献率最大,占总量的74.7%。从正规化训练增益影响看(图4),单因子作用时,最冷季度降水量(bio_19)、最干季度降水量(bio_17)、最干月降水量(bio_14)、降水量变异系数(bio_15)对水菜花影响最大。综合来看,降水量是对水菜花分布影响最大的因子。
2.2 水菜花适生区的生态特征
拟合结果表明,水菜花总适生面积约996.7 km2,主要分布在海口市及周边地区;其中,高度适宜生境138.5 km2,中度适宜生境262.0 km2,低适宜生境约596.2 km2。高适宜区主要分布在海口市龙泉镇、遵谭镇、新坡镇交汇处以及龙塘镇、龙桥镇和城西镇的交汇处(图5)。

图3 环境变量的贡献率特征。环境变量见表1。下同
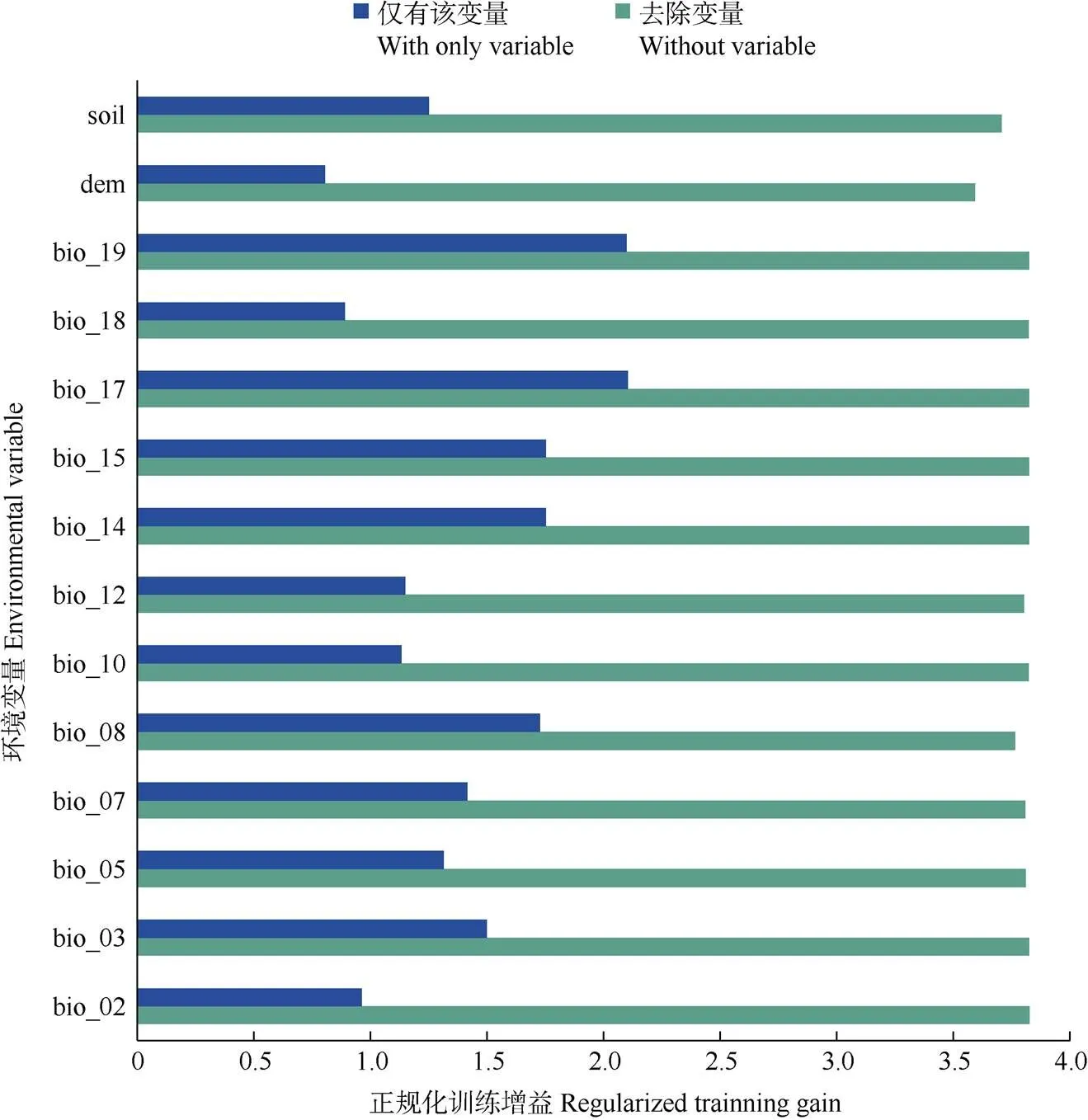
图4 水菜花环境变量的刀切法检验
对水菜花分布影响最大的等温性、最冷季度降水量、土壤类型和年均降水量4个因子单因子建模结果显示:水菜花在等温性因子条件下的适宜生长区间为20%~43%,当等温性为37%时,分布概率达到最高值;年降水量1 200~1 800 mm和最冷季度降水量85~120 mm,适宜水菜花生存,其中1 600和105 mm为最适临界点;在土壤类型方面,石灰土和水稻土最适宜水菜花生长;以上各变量的适生区间阈值与响应曲线具有很高的一致性,结果可信度较高(图6)。
2.3 气候变化对水菜花潜在地理分布的影响预测
对全球气候模式(GCMs)全新世中期和BCC-CSM2-MR气候系统模式2070年4种不同社会经济假设驱动情景下的水菜花适宜生境进行模拟。结合当前气候拟合结果,获取水菜花种群在气候变化背景下适生面积、重心及其潜在分布区的变化。
结果表明(图5),全新世中期适宜生境总面积1 008.23 km2,比当前气候条件下多11.46 km2;2070年4种不同社会经济路径下的适宜生境总面积分别为1 010.99、1 134.76、1 015.40和1 103.49 km2, 较当前气候条件下分别新增14.22、138、18.63和106.72 km2。从面积变化的总体情况看,水菜花种群适宜生境从历史到未来,总面积先减少后增加; 高适生区和中适生区面积均以当前气候条件下最高,未来呈下降趋势;低适生区面积则以当前气候条件最低,未来会有一定增长。高度和中度适宜生境区将在当前气候分布区的基础上逐渐收缩,西部及南部则会出现新增低适宜生境。
2.4 气候变化对生境空间转移的影响
由图7可知,不同气候时期水菜花适宜生境重心呈西南-东北-西南转移的空间格局。全新世中期到当前气候条件下,适宜生境重心向东北方向转移约4 km;当前气候条件到2070年的4种不同社会经济假设路径情景下,适宜生境均向西南方向转移,转移距离分别为SSP1_2.6 (11 km)、SSP2_4.5 (13 km)、SSP3_7.0 (17 km)和SSP5_8.5 (11 km)。
从生境新增和消失情况来看,全新世中期到当前气候条件下,适宜生境新增214.90 km2,减少226.36 km2;新增区主要位于海口市北部及西南部地区,减少区主要分布在海口市东北部和定安县的北部地区。2070年气候条件下4个不同经济路径, 新扩增区域的面积分别为363.13、382.15、381.31和416.06 km2,消失区域面积分别为348.90、244.16、362.67和309.34 km2,其中新增区域主要位于海口市北部、中部、南部,以及澄迈县东部和定安县北部地区;消失区域主要位于海口市西部、西南部和东北部区域(图8)。
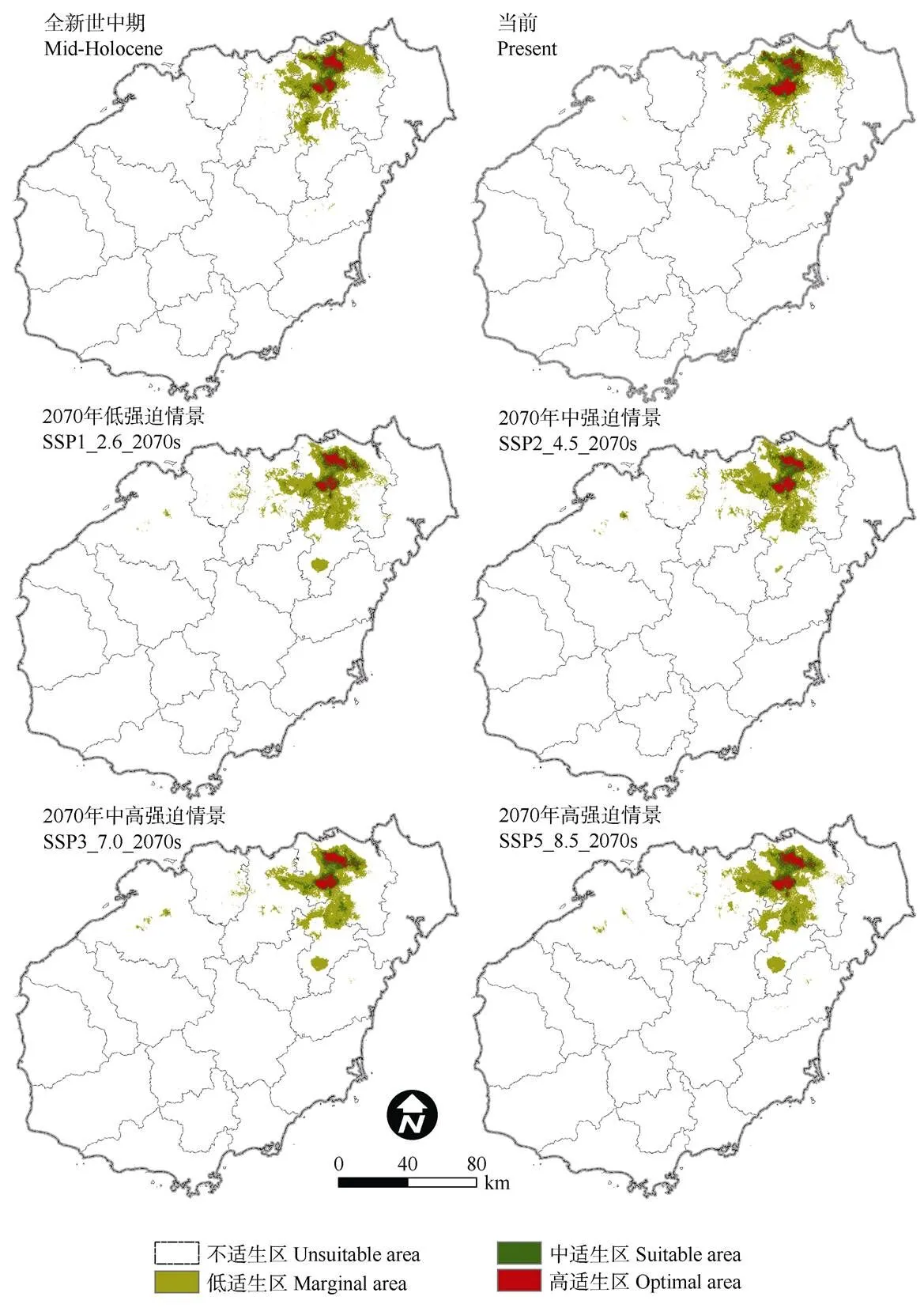
图5 不同气候情景下水菜花适宜生境的空间分布
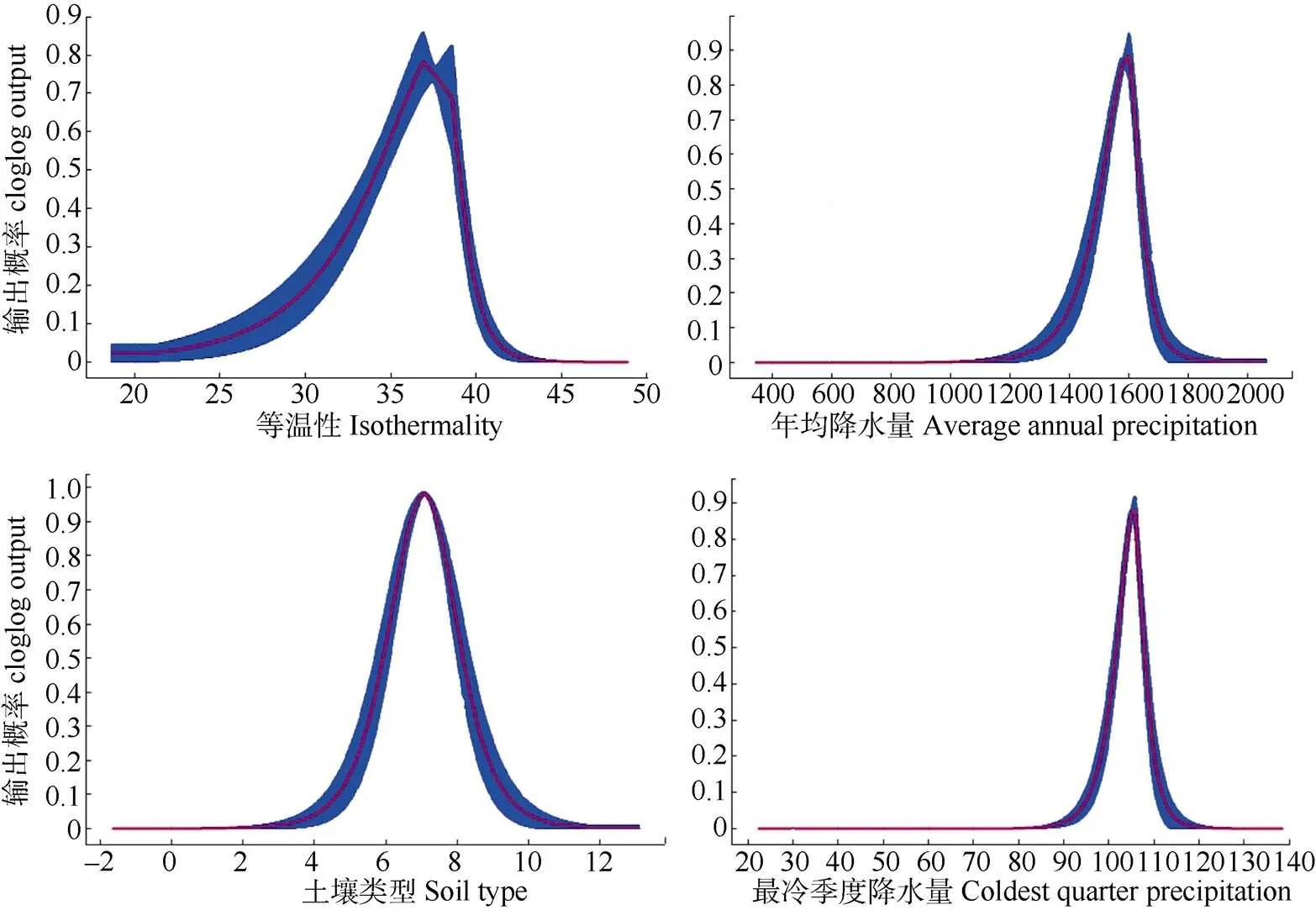
图6 水菜花分布与主导环境因子的关系
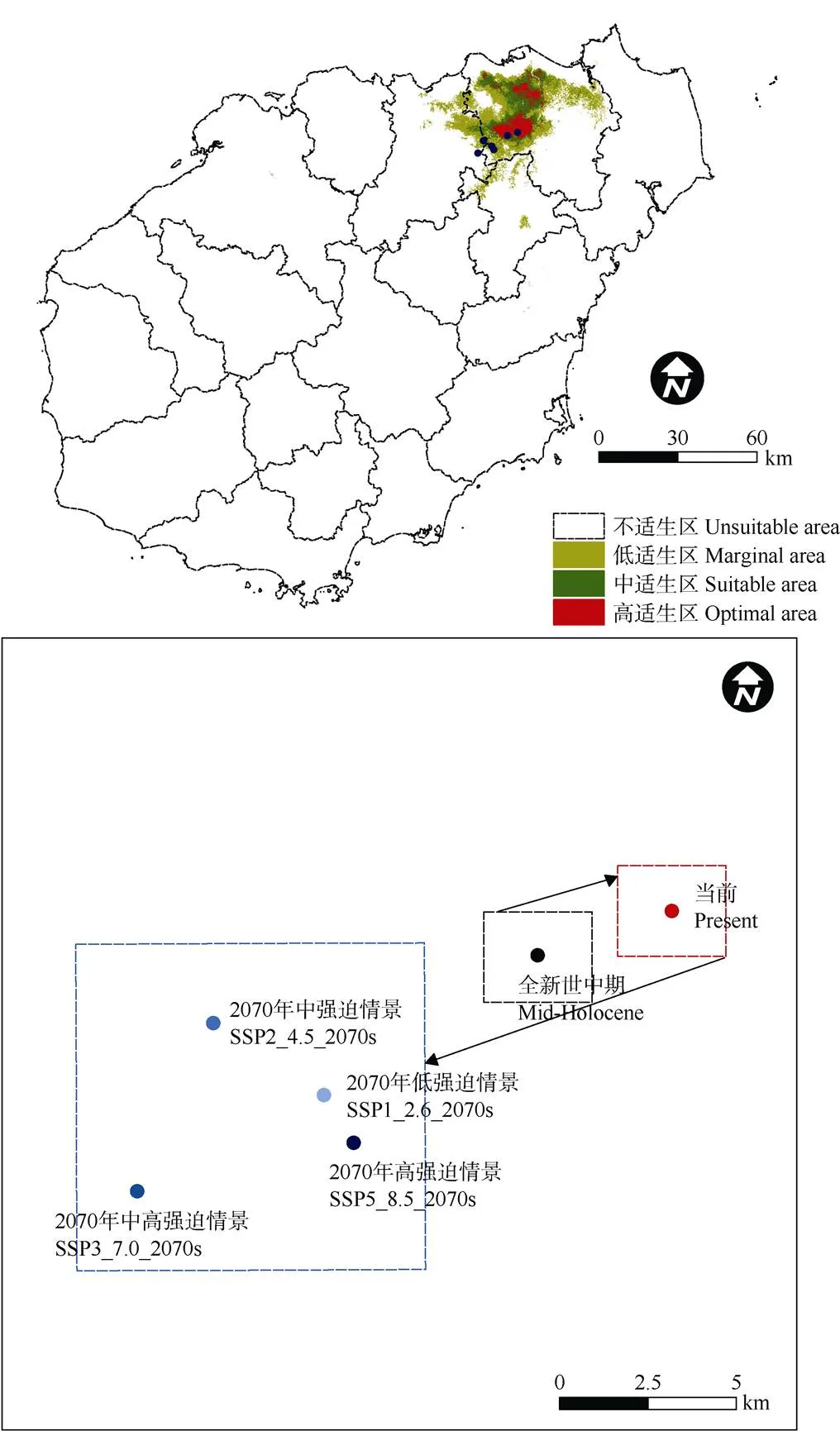
图7 不同气候情景下水菜花适宜生境的质心变化
总体来看,水菜花适宜生境从全新世纪中期到当前气候条件下,重心逐渐向东北方向转移,其中新增适宜生境主要位于原分布区的北部及西南部区域,消失区域位于原分布区外围的东北和西南区域。到2070年,适宜生境重心重新向西南方向更大幅度转移,原分布区的东部、南部会出现较大块新增适宜生境,而东北部、西北部及西南部则会有适宜生境消失。
3 讨论
3.1 气候环境对水菜花种群分布的限制
湿地对气候变化有着响应的敏感,气温和降水的变化能够直接影响湿地生物群落、蒸散发速率以及水文、生物区系的变化,进而改变整个生态系统格局和稳定性[21–22]。本研究中模型拟合结果同样表明水菜花种群的生存对温差与降水量的变化较为敏感,同时在土壤类型上也有着较为特定的要求。Zhang等[23]对洞庭湖湿地植被变化的时空格局和因素的研究表明,气候变化对湿地植被的威胁比重占到全部影响因素的59.19%。水菜花的生存和生命周期的完成与水有着密切关系,外界对水菜花的影响大多是以水为媒介完成的[2]。Shen等[3]对3种不同生境中水菜花种群动态及其影响因素的研究表明, 水深和浑浊度是决定水菜花种群繁育和扩增的关键因子,过浅或过于浑浊的水环境会制约水菜花的生长。湿地面积和径流深度,与温度和降水量有着极显著的相关性[24],湿地生境的稳定和变化直接或间接影响水菜花种群的分布。杨丹等[25]研究了湖泊湿地生态系统稳态转变及对气候变化的响应,认为降水对湿地稳态转变有着显著的影响,湿地稳态转变与长期的水量变化有关。刘志伟等[26]对青藏高原湿地变化及其驱动力的研究结果也表明气温升高、地表蒸散发量增大、降水量增加是影响青藏高原湿地变化的主要自然因素。另外,过高的气温会使入侵植物快速生长[27],入侵植物对植物群落的威胁已得到多方论证,水葫芦()的疯长是水菜花种群繁衍的主要入侵威胁,水葫芦依仗其极强的适生性和快速繁殖能力,广泛分布在研究区域的溪流、沼泽、库塘等湿地中,严重掠夺和挤压水菜花的生存空间[28–29]。
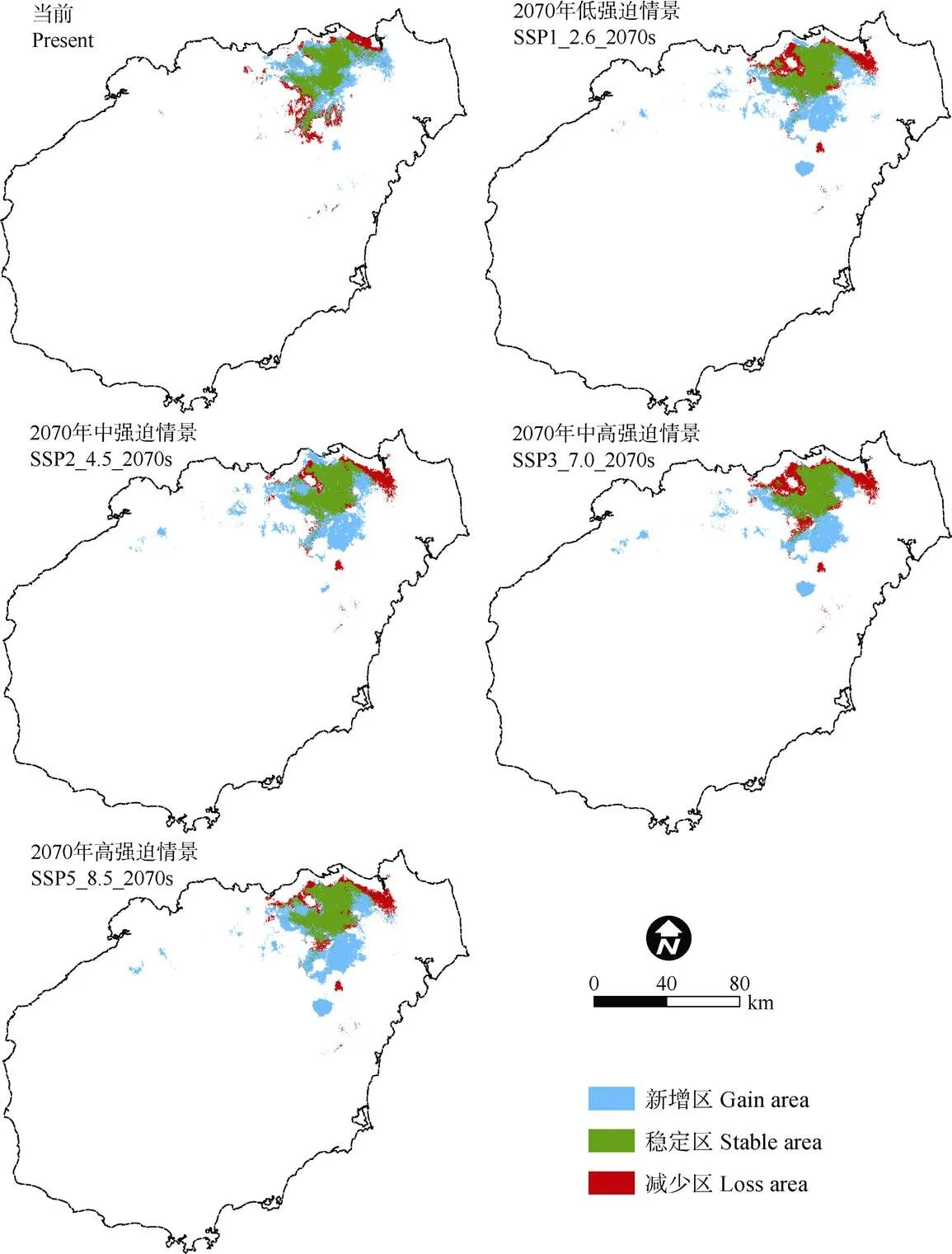
图8 不同气候情景下水菜花适宜生境的空间变化
3.2 气候变化对水菜花生境空间分布的影响
水菜花种群在全新世中期、当前和未来2070年4种不同经济路径气候背景下适宜生境面积呈先减少后增加趋势,以2070年4.5中等强迫模式下的生境面积最大,重心呈西南-东北-西南转移的空间格局。
研究表明,到2050年全球约有15%~37%的物种存在灭绝的风险[30]。本研究中,当前至2070年期间,适宜生境的东部、南部将出现较大块新增适宜生境,而原分布区的东北部、西北部及西南部区域将有大面积适宜生境消失,重心重新向西南方向进行更大幅度转移。另外从不同等级适宜生境的演变情况来看,高适宜区和中适宜区以当前气候环境下面积最大,在未来分别会发生5%~15%和10%~ 21%的衰减,主要围绕在当前分布区的外围区域由外向内缩紧;而生境的增加则主要集中在低适宜生境区,增幅为2%~14%,集中分布在当前适生区的西部及南部。
社会经济因素是研究区及周边湿地变化的主要因素[31]。政府间气候变化专门委员会(IPCC)发布的2070年不同社会经济路径情景下未来气候预测数据表明,2070年等温性较当前气候向南逐渐升高,年均降水量全岛整体提升200~800 mm不等,最冷季度降水量在琼北地区发生下降;综合本研究模拟结果中对水菜花影响最大的环境因素临界阈值得出, 降水量的增大和温度的升高导致了水菜花对环境因子的适生区间不断向南部转移,进而也引发了当前气候条件下水菜花高适宜和中适宜生境不断向内部收缩。
Boniface等[32]对非洲7种水车前属植物在未来不同CO2排放浓度路径下物种分布的研究表明,海拔和气候(气温、降水量)对水车前属植物的分布影响显著,气候变化背景下水车前属植物生境会发生衰减,重心逐渐向北移动。另外,一些学者在对桫椤()、绵刺()等种群潜在空间分布情况的研究表明,未来气候情景下,种群适宜生境总面积呈增加趋势,其中,高度和中度适宜生境减少,低适宜生境面积增加[17,33], 与本研究的结论相似;但不同于桫椤[17]、青冈()[34]、秤锤树属()[35]、梭梭()[36]等种群适宜生境重心向北部、高纬度、高海拔地区转移的情况,本研究中水菜花分布总体重心呈向南和低纬度转移趋势,与秦媛媛等[33]对绵刺的潜在地理分布研究结论相似, 但该文献中种群逐渐向高海拔地区转移的特征又与水菜花适宜生境转移格局相异。可以看出,物种间自身生物学特性的差异,形成了对不同环境因子的依赖,而环境因子空间分布的异质性,又能进一步引发物种区域演替格局的差异。
综合来看,温差、降水量、土壤类型、水环境深度、浊度以及入侵植物等,是制约水菜花种群繁育和扩增的主要关键因素,本研究仅从气候、地形和土壤变量角度模拟水菜花种群的适生区域,结果或存在一定片面性;同时,样点较少、气候环境基础数据尺度较大等问题,也进一步增加了研究结果的不确定性;在实际应用及后续研究中,需结合人为扰动、水质情况、入侵植物等多因素共同论证, 确保研究结果更具科学性和指导意义。
本研究结果表明,影响水菜花种群分布的主要自然因素有等温性、最冷季度降水量、土壤类型和年均降水量;在气候变化背景下,全新世中期-当前时期-2070年,水菜花适宜生境面积先减小后增大, 当前气候条件下水菜花适宜生境面积最小,2070年时期水菜花适宜生境得到扩增,但增加部分均为低适宜区,高适宜和中适宜区均发生衰减;未来演替中降水和温度的变化将引起水菜花生境收缩,在南部地区会出现新的适宜区,整体重心逐渐呈西南-东北-西南方向转移。
[1] YANG X B, LI D H, CHEN Y K, et al. The Colored Illustrated Flora of Hainan Province, Vol. 1-14 [M]. Beijing: Science Press, 2015. [杨小波, 李东海, 陈玉凯, 等. 海南植物图志, 第1~14卷 [M]. 北京:科学出版社, 2015.]
[2] ZHAO Z C, SUN X Z, WANG H Q. A study on the ecology of the fresh water Hydrocharidaceae in south China [J]. Acta Ecol Sin, 1984, 4(4): 354–363. [赵佐成, 孙祥钟, 王徽勤. 华南地区淡水水鳖科植物的生态特征和群落学观察 [J]. 生态学报, 1984, 4(4): 354–363.]
[3] SHEN Y C, LEI J R, SONG X Q, et al. Annual population dynamics and their influencing factors for an endangered submerged macrophyte () [J]. Front Ecol Evol, 2021, 9: 688304. doi: 10.3389/ FEVO.2021.688304.
[4] JIAN Y X, YANG G M, PENG Y H, et al. Studies on daryotypes of(L.) Pers. and(Wall.) Dandy [J]. J Hunan Coll Trad Chin Med, 1996, 16(1): 56–58. [简永兴, 杨广民, 彭映辉, 等. 水白菜与水菜花的核型分析 [J]. 湖南中医学院学报, 1996, 16(1): 56–58.]
[5] ZHANG Q F, SHEN Z X, LI F Y, et al. Complete chloroplast genome sequence of an endangeredand its phylogenetic analysis [J]. Mitochondrial DNA BResour, 2020, 5(3):2209–2210. doi: 10.1080/23802359.2020.1768921.
[6] WANG H X, GUO J L, LI Z M, et al. Characterization of the complete chloroplast genome of an endangered aquatic macrophyte,(Hydrocharitaceae) [J]. Mitochondrial DNA BResour, 2019, 4(1): 1839–1840. doi: 10.1080/23802359.2019.1612719.
[7] WANG S N, LI P P, LIAO Z Y, et al. Adaptation of inorganic carbon utilization strategies in submerged and floating leaves of heteroblastic plant[J]. Environ Exp Bot, 2022, 196: 104818. doi: 10.1016/j.envexpbot.2022.104818.
[8] HUANG W M, HAN S J, XING Z F, et al. Responses of leaf anatomy and CO2concentrating mechanisms of the aquatic plantto variable CO2[J]. Front Plant Sci, 2020, 11: 1261. doi: 10.3389/fpls. 2020.01261.
[9] FARASHI A, KABOLI M, KARAMI M. Predicting range expansion of invasive raccoons in northern Iran using ENFA model at two different scales [J]. Ecol Inform, 2013, 15: 96–102. doi: 10.1016/j.ecoinf.2013. 01.001.
[10] KONG W Y, LI X H, ZOU H F. Optimizing MaxEnt model in the prediction of species distribution [J]. Chin J Appl Ecol, 2019, 30(6): 2116–2128. [孔维尧, 李欣海, 邹红菲. 最大熵模型在物种分布预测中的优化 [J]. 应用生态学报, 2019, 30(6): 2116–2128. doi: 10.13287/ j.1001-9332.201906.029.]
[11] LIN H, WANG Y S. Analysis and research on water quality of Meishe River national wetland park in Haikou [J]. Trop For, 2020, 48(4): 42– 46. [林华, 王耀山. 海口美舍河国家湿地公园水质分析研究 [J]. 热带林业, 2020, 48(4): 42–46. doi: 10.3969/j.issn.1672-0938.2020.04.009.]
[12] YANG X B, WU Q S, LI Y L, et al. Characteristic of tropical forest composition in north of Hainan Island [J]. Sci Sil Sin, 2005, 41(3): 19– 24. [杨小波, 吴庆书, 李跃烈, 等. 海南北部地区热带雨林的组成特征 [J]. 林业科学, 2005, 41(3): 19–24. doi: 10.3321/j.issn:1001- 7488.2005.03.004.]
[13] WANG X. Study on interconnected river network system and hydro- dynamic water environment in Haikou City [D]. Guangzhou: South China University of Technology, 2018. [王欣. 海口市河湖水系连通与水动力水环境研究 [D]. 广州: 华南理工大学, 2018.]
[14] SHEN Y C, LU G, LIU S B, et al. Characteristics of plant distribution in volcanic lava wetlands in Yangshan, Haikou [J]. Wetland Sci, 2019, 17(5): 493–503. [申益春, 卢刚, 刘寿柏, 等. 海口羊山火山熔岩湿地中的植物分布特征 [J]. 湿地科学, 2019, 17(5): 493–503. doi: 10. 13248/j.cnki.wetlandsci.2019.05.001.]
[15] YANG X Q, KUSHWAHA S P S, SARAN S, et al. Maxent modeling for predicting the potential distribution of medicinal plant,L. in Lesser Himalayan foothills [J]. Ecol Eng, 2013, 51: 83– 87. doi: 10.1016/j.ecoleng.2012.12.004.
[16] ZHANG H, ZHAO H X, XU C G. The potential geographical distri- bution ofunder climate change in China [J]. Chin J Ecol, 2021, 40(4): 968–979. [张华, 赵浩翔, 徐存刚. 气候变化背景下孑遗植物桫椤在中国的潜在地理分布 [J]. 生态学杂志, 2021, 40(4): 968–979. doi: 10.13292/j.1000-4890.202104.022.]
[17] ZHANG H, ZHAO H X, WANG H. Potential geographical distribution ofin China under future climate change scenarios based on Maxent model [J]. Acta Ecol Sin, 2020, 40(18): 6552–6563. [张华, 赵浩翔, 王浩. 基于Maxent模型的未来气候变化情景下胡杨在中国的潜在地理分布 [J]. 生态学报, 2020, 40(18): 6552–6563. doi: 10.5846/stxb201906111232.]
[18] ARAÚJO M B, GUISAN A. Five (or so) challenges for species distribution modelling [J]. J Biogeogr, 2006, 33(10): 1677–1688. doi: 10.1111/j.1365-2699.2006.01584.x.
[19] PEARSON R G, RAXWORTHY C J, NAKAMURA M, et al. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar [J]. J Biogeogr, 2007, 34(1): 102–117. doi: 10.1111/j.1365-2699.2006.01594.x.
[20] NING Y, LEI J R, SONG X Q, et al. Modeling the potential suitable habitat of, a limestone-endemic plant [J]. Chin J Plant Ecol, 2018, 42(9): 946–954. [宁瑶, 雷金睿, 宋希强, 等. 石灰岩特有植物海南凤仙花潜在适宜生境分布模拟 [J]. 植物生态学报, 2018, 42(9): 946–954. doi: 10.17521/cjpe.2018.0066.]
[21] SEMENIUK C A, SEMENIUK V. The response of basin wetlands to climate changes: A review of case studies from the Swan Coastal Plain, south-western Australia [J]. Hydrobiologia, 2013, 708(1): 45–67. doi: 10.1007/s10750-012-1161-6.
[22] BARROS D F, ALBERNAZ A L M. Possible impacts of climate change on wetlands and its biota in the Brazilian Amazon [J]. Braz J Biol, 2014, 74(4): 810–820. doi: 10.1590/1519-6984.04013.
[23] ZHANG M, LIN H, LONG X R, et al. Analyzing the spatiotemporal pattern and driving factors of wetland vegetation changes using 2000—2019 time-series Landsat data [J]. Sci Total Environ, 2021, 780: 146615. doi: 10.1016/j.scitotenv.2021.146615.
[24] YU C L, WANG Z C, LIU D, et al. Evolution process and driving force analysis of natural wetlands in Xiliao River Basin based on SWAT model [J]. Trans Chin Soc Agric Eng, 2020, 36(22): 286–297. [于成龙, 王志春, 刘丹, 等. 基于SWAT模型的西辽河流域自然湿地演变过程及驱动力分析 [J]. 农业工程学报, 2020, 36(22): 286– 297. doi: 10.11975/j.issn.1002-6819.2020.22.032.]
[25] YANG D, WANG W J, WU X Q, et al. Regime shifts of Anguli wetland and its response to climate change from 1985 to 2016 [J]. Res Environ Sci, 2021, 34(12): 2954–2961. [杨丹, 王文杰, 吴秀芹, 等. 1985—2016年安固里淖湖泊湿地生态系统稳态转变及对气候变化的响应 [J]. 环境科学研究, 2021, 34(12): 2954–2961. doi: 10.13198/j. issn.1001-6929.2021.09.17.]
[26] LIU Z W, LI S N, WEI W, et al. Research progress on alpine wetland changes and driving forces in Qinghai-Tibet Plateau during the last three decades [J]. Chin J Ecol, 2019, 38(3): 856–862. [刘志伟, 李胜男,韦玮, 等. 近三十年青藏高原湿地变化及其驱动力研究进展 [J]. 生态学杂志, 2019, 38(3): 856–862. doi: 10.13292/j.1000-4890.2019 03.002.]
[27] SCHEFFER M, SZABÓ S, GRAGNANI A, et al. Floating plant dominance as a stable state [J]. Proc Natl Acad Sci USA, 2003, 100(7): 4040–4045. doi: 10.1073/pnas.0737918100.
[28] FAN Z H, HUANG Z, ZHOU X H. Optimizing and improving strategy for Haikou wetland ecological system [J]. CS For Invent Plann, 2019, 38(2): 68–72. [范志浩, 黄铮, 周湘红. 海口湿地生态系统优化提升策略 [J]. 中南林业调查规划, 2019, 38(2): 68–72. doi: 10.16166/j. cnki.cn43-1095.2019.02.016.]
[29] SHEN Y C, REN M X, LI W, et al. Landscape plant community and its application model in Yangshan Wetland [J]. Jiangsu Agric Sci, 2021, 49(11): 92–97. [申益春, 任明迅, 黎伟, 等. 羊山湿地景观植物群落与景观应用模式 [J]. 江苏农业科学, 2021, 49(11): 92–97. doi: 10. 15889/j.issn.1002-1302.2021.11.016.]
[30] THOMAS C D, CAMERON A, GREEN R E, et al. Extinction risk from climate change [J]. Nature, 2004, 427(6970): 145–148. doi: 10. 1038/nature02121.
[31] ZHONG Z Q, QIU P H, YANG X. Analysis of wetland changes and driving forces in Haikou in the last 30 years [J]. J Hainan Norm Univ (Nat Sci), 2021, 34(2): 215–226. [钟尊倩, 邱彭华, 杨星. 海口市近30年来湿地变化及其驱动力分析 [J]. 海南师范大学学报(自然科学版), 2021, 34(2): 215–226. doi: 10.12051/j.issn.1674-4942.2021.02.014.]
[32] NGAREGA B K, NZEI J M, SAINA J K, et al. Mapping the habitat suitability ofspecies in Africa [J]. Plant Divers, 2022, 44(5): 468–480. doi: 10.1016/j.pld.2021.12.006.
[33] QIN Y Y, LU K, DU Z Y, et al. Potential changes in the geographical distribution of the relict plantMaxim. in China under climate change scenarios [J]. Acta Ecol Sin, 2022, 42(11): 4473– 4484. [秦媛媛, 鲁客, 杜忠毓, 等. 气候变化情景下孑遗植物绵刺在中国的潜在地理分布 [J]. 生态学报, 2022, 42(11): 4473–4484. doi: 10.5846/stxb202106111553.]
[34] ZHANG L J, LI Y H, REN H, et al. Prediction of the suitable distribution ofand its implications for the northern boundary of subtropical zone of China [J]. Geogr Res, 2020, 39(4): 990–1001. [张立娟, 李艳红, 任涵, 等. 气候变化背景下青冈分布变化及其对中国亚热带北界的指示意义 [J]. 地理研究, 2020, 39(4): 990–1001. doi: 10.11821/dlyj020190306.]
[35] YANG T, WANG S T, WEI X Z, et al. Modeling potential distribution of an endangered genus () endemic to China [J]. Plant Sci J, 2020, 38(5): 627–635. [杨腾, 王世彤, 魏新增, 等. 中国特有属秤锤树属植物的潜在分布区预测 [J]. 植物科学学报, 2020, 38(5): 627– 635. doi: 10.11913/PSJ.2095-0837.2020.50627.]
[36] MA S M, WEI B, LI X C, et al. The impacts of climate change on the potential distribution of[J]. Chin J Ecol, 2017, 36(5): 1243–1250. [马松梅, 魏博, 李晓辰, 等. 气候变化对梭梭植物适宜分布的影响 [J]. 生态学杂志, 2017, 36(5): 1243–1250. doi: 10.13292/j.1000-4890.201705.032.]
Potential Habitat Selection and Spatial Pattern Prediction of
WU Tingtian, LEI Jinrui*, CHEN Zongzhu, CHEN Xiaohua, LI Yuanling
(Hainan Key laboratory of Monitoring and Application of Tropical Forestry Resources, Haikou Wetland Protection Engineering Technology Research and Development Center, Hainan Academy of Forestry (Hainan Academy of Mangrove), Haikou 571100, China)
, a wild plant under State second-class protection, likes to be grown in clean water, and is extremely sensitive to environmental changes. It is one of the key indicator species for testing wetland environment and climate change. In China, itisonly distributed sporadically in volcanic lava wetland in northern Hainan, the survival condition is not optimistic. The study of potential habitat selection and spatial pattern evolution ofpopulation will help strengthen the conservation of endangered species and the restoration as well as the management of wetland ecosystem. Based on GIS platform and MaxEnt model, combining with climate, topography and soil factors, the environmental limiting factors ofpopulation and the evolution pattern of potential suitable habitat in the context of climate change were studied. The results showed thatpopulation was sensitive to temperature difference and precipitation change. Besides, isothermality, precipitation in the coldest quarter, type of soil and annual precipitation were key environmental factors affecting the distribution ofpopulation. Under the background of climate change from the middle Holocene to current to 2070s, the area of suitable habitat ofdecreased first and then increased, and the distribution center showed a transfer pattern from southwest to northeast to southwest. Under the future climate scenarios, the highly and moderately suitable habitats were decrease, while the low suitable habitats will increase. The new habitats will appear in the southern region, and the suitable habitats will decrease in the northeast, northwest and southwest. Therefore, the potential habitat selection and spatial distribution characteristics ofpopulation were discussed from the perspective of climate environment, which could provide reference and theoretical basis for endangered species conservation, wetland management and biodiversity maintenance.
; Climate change; Potential habitat; Environmental factor
10.11926/jtsb.4720
2022-08-25
2022-10-21
国家自然科学基金项目(32260106);海南省省属科研院所技术开发专项(KYYS-2021-21, KYYS-2021-04)资助
This work was supported by the National Natural Science Foundation of China (Grant No. 32260106), the Project for Technology Development of Hainan Provincial Institutes (Grand No. KYYS-2021-21, KYYS-2021-04).
吴庭天(1990年生),男,硕士,助理研究员,主要从事森林生态学和植物生态学研究。E-mail: wutingtian@126.com
* 通讯作者 Corresponding author. E-mail: raykingre@163.com
猜你喜欢
《学习方法报》地理商务星球七年级(2023年13期)2023-10-11
今日农业(2022年16期)2022-11-09
落叶果树(2021年6期)2021-02-12
孩子·小学版(2020年10期)2020-12-07
启蒙(3-7岁)(2019年8期)2019-09-10
江西农业(2018年23期)2018-02-11
民族音乐(2017年6期)2017-04-19
水利科技与经济(2016年6期)2016-04-22
体育科技(2016年2期)2016-02-28
西华师范大学学报(自然科学版)(2015年3期)2015-02-27
