
不同生长光强下8种红树植物幼苗叶片的光响应特征
2024-02-02 06:19:22朱一民李婷孙当歌叶万辉沈浩
热带亚热带植物学报 2024年1期
朱一民, 李婷, 孙当歌, 叶万辉,4, 沈浩,4*
不同生长光强下8种红树植物幼苗叶片的光响应特征
朱一民1,2,3, 李婷1,2,3, 孙当歌1,2,3, 叶万辉1,2,3,4, 沈浩1,2,3,4*
(1. 中国科学院华南植物园,中国科学院退化生态系统植被恢复与管理重点实验室,广东省应用植物学重点实验室,广州 510650;2. 华南国家植物园,广州 510650;3. 中国科学院大学,北京 1000498;4.南方海洋科学与工程广东省实验室(广州),广州 51145)
为探讨红树植物光适应的生理生态策略,对6种真红树植物[无瓣海桑()、秋茄()、木榄()、桐花树()、老鼠簕()、卤蕨()]和2种半红树植物[银叶树()、黄槿()]的1 a生幼苗在不同生长光强(自然光强的100%、45%、30%、10%)下的光合光响应特征进行了研究。结果表明,不同生长光强对红树植物光响应特征的影响因物种而异,遮荫显著提高了秋茄和木榄的最大净光合速率(Pmax),而对其他红树植物的Pmax没有显著影响;秋茄在45%光强下具有较高的Pmax,木榄的Pmax则在45%和30%光强下显著高于其他2个处理。随着生长光强的下降,秋茄幼苗叶片的光饱和点显著上升,木榄、老鼠簕和卤蕨的光补偿点呈下降趋势,木榄和卤蕨的表观量子效率升高的同时暗呼吸速率下降。木榄、老鼠簕和卤蕨具有较强的耐荫性,适宜种植在光强较弱的林下;无瓣海桑、秋茄、桐花树、银叶树和黄槿则适宜作为中上层树种或在郁闭度较低的林下种植。
红树植物;遮荫;光合作用;光强
红树植物是生长在热带、亚热带海岸潮间带的乔木、灌木或草本植物,根据其分布特征可分为真红树植物和半红树植物[1–2]。红树林因其抵挡海啸、飓风及台风的能力远胜于人类工程而素有“海岸卫士”之称,同时红树林还具有提供生物栖息地、保护生物多样性、原料供给以及净化环境等生态功能[3]。自20世纪50年代以来,由于大面积围垦和海岸带开发以及过度砍伐等人类活动的影响,红树林发生了急剧退化[4]。全球35%的红树林已经消失,我国红树林总面积从20世纪50年代到21世纪初减少近60%[5–6]。随着人们对红树林生态价值的认识进一步加深,全球开始日益重视红树林的生态恢复与重建研究[7]。近年来我国华南沿海大范围选用无瓣海桑()等速生树种进行红树林恢复和重建,这种高大、生长迅速的树种降低了林下环境的光照水平,不可避免的会对林下本土红树树种的生长、繁殖和扩散造成一定影响,导致红树林生态系统稳定性降低。当前亟需对无瓣海桑人工林的群落结构进一步改造和优化,而首先要理解红树植物光适应的生理生态策略,为此,开展了外来种无瓣海桑与7种本土红树植物在不同光强条件下的生长和生物量分配[8]以及生理生态特性的系统研究,旨在为红树林重建和改造过程中的物种优化配置提供科学指导。
光是影响植物生长发育和生存及群落结构的最重要环境因子之一[9–10],植物对于光照条件变化的响应一直都是植物生理生态学所关注的重点问题[11–12]。当环境光照不足时,可能会影响植物的固碳功能,而当光照过强时则可能会导致光合系统受损[13];在光照条件改变时,植物会启动一系列生理生态响应,特别是光合生理响应,以适应当前的光照条件[14–15]。植物净光合速率对光强变化的响应反映了环境改变对于植物光合生理、生化代谢过程的影响,其拟合参数反映不同类型植物生理适应性的差异[16],其中,叶片的最大光合速率(maximum net photosynthetic rate, Pmax)反映了植物叶片的最大光合能力,而表观量子效率(apparent quantum yield,)是光合作用中光能转化效率的指标之一,反映植物在对光能吸收、转换和利用的能力,值高说明植物叶片有较强的光能捕获能力和光能转化效率[17–18]。光饱和点(light saturation point, LSP)、光补偿点(light compensation point, LCP)和暗呼吸速率(dark respi- ration rate, Rd)则反映了植物对于光能的利用效率以及植物适应不同光照强度的能力。
红树林作为生长在陆海之间的潮间带的重要植被类型,在其生长发育过程中同时受到光照、盐度和周期性水淹等因子的作用。而目前对于红树林生理的研究多集中在红树植物对盐度、水淹的生理响应方面,关于光照条件变化对红树植物幼苗生理特征的影响的研究相对较少[19–20]。为此,本研究选择我国珠三角地区常见的8种红树植物,包括6种真红树植物: 无瓣海桑、秋茄()、木榄()、桐花树()、老鼠簕()、卤蕨()和2种半红树植物: 银叶树()、黄槿()为研究对象,比较这8种红树植物幼苗在不同生长光强条件下的光合光响应特征的差异,旨在揭示红树植物光适应的生理生态策略,为人工红树林群落的优化配置和红树林林分改造提供理论指导,为红树林的保护和恢复实践提供科学参考。
1 材料和方法
1.1 试验区和种源区自然概况
遮荫试验于2013年10月至2014年10月在位于广东省广州市的中国科学院华南植物园大型实验生态学综合试验场进行。地理位置为23°10′42.79″ N, 113°21′25.28″ E,海拔40 m。该地区属南亚热带海洋季风气候类型,年平均气温20 ℃~22 ℃,年平均相对湿度77%,年均降雨量为1 982.7 mm。
试验红树幼苗购自广东珠海淇澳红树林自然保护区附近的苗圃,该保护区位于22°23′40″~22° 27′38″ N,113°36′40″~113°39′15″ E,属南亚热带海洋性季风气候,区内年均温为22.2 ℃,基本无霜期,年降雨量为1 875.7 mm,4月—10月降雨量占全年雨量的84%左右[21]。保护区受降雨、江河径流和潮汐的影响,海水盐度在3.31‰~7.05‰之间,海域潮汐属不正规半日潮[22]。
1.2 材料
2013年8月,在珠海淇澳岛红树林自然保护区附近的苗圃内选择苗龄1 a生、长势均一的8种红树植物幼苗[8]:无瓣海桑()、秋茄()、木榄()、桐花树()、老鼠簕()、卤蕨()、银叶树()和黄槿(),其中,无瓣海桑为外来红树植物,其他7种均为本土种。无瓣海桑、秋茄、木榄、桐花树、老鼠簕和卤蕨6种为真红树,银叶树和黄槿2种为半红树植物。
1.3 试验设计
本研究共设置4个光强梯度,分别为自然光强的100% (T0,对照)、45% (T1)、30% (T2)和10% (T3)[23],用不同透光度的黑色尼龙网遮荫达到不同光强。将栽植好的幼苗标号并分别置于4个光强处理区中,每处理每种10~12株。幼苗栽种于13.4 L花盆中(上口内径30 cm, 盆底内径21 cm, 高26 cm), 每盆1株,花盆放置于含有人工配置海水的塑料盆(内径27.5 cm,高10.5 cm)中,进行遮荫处理1 a。栽培基质使用珠海淇澳岛的海泥,其盐度、含水量、全氮含量和有机碳含量分别为8.3%、42.9%、1.61%和1.85%。人工海水以粗海盐和自来水调配而成,含盐量模拟淇澳岛近岸海水平均盐度,约为6‰。试验期间监测盐度,适时补充人工海水。
1.4 光响应曲线的测定
不同光强下每树种选取3~5棵幼苗,每棵幼苗随机选取6~10片成熟叶片,于2014年8月18日至9月25日上午8:00-11:00,使用LI-6400便携式光合作用测定系统(LI-COR Inc., Lincoln, NE, USA)进行光响应曲线的测定。设定叶温为30 ℃,CO2浓度为360mol/mol,用连体叶片分别在0~2 000mol photons/(m2·s)(内置光源)的光照强度(光量子通量密度, photosynthetic photon flux density, PPFD)范围内测定光合作用光响应动态。在设置最大和最小等待时间后,由仪器自动记录数据。测定前将待测幼苗移到遮荫棚外适应1~2 d。
1.5 光响应参数

1.6 数据分析
所有数据用Microsoft Excel 2013软件整理,采用SPSS 13.0软件(SPSS Inc., Chicago, IL, USA)进行统计分析,以<0.05表示有显著差异。各处理光强间光响应特征参数的差异使用单因素方差分析(One-Way ANOVA)方法进行显著性检验,并以Tukeycomparisons法进行多重比较。
2 结果和分析
当PPFD为0~500mol photons/(m2·s)时,8种红树幼苗叶片的Pn均呈快速上升趋势(图1)。当PPFD大于500mol photons/(m2·s)后,Pn的变化趋于平缓。相比于遮荫处理,全光照下秋茄、木榄、卤蕨、银叶树和黄槿的叶片Pn增长幅度较小。除老鼠簕外,其他红树植物的Pn在遮荫下均比100%光强更高。无瓣海桑的Pn在100%、45%和30%光照下的变化趋势相近,其增长幅度在10%光照下有明显下降。
生长光强对红树植物的光响应曲线参数的影响因物种而异(表1)。生长光强对秋茄和木榄幼苗的叶片最大净光合速率(Pmax)有显著影响,而无瓣海桑、桐花树、老鼠簕、卤蕨、银叶树和黄槿幼苗无显著差异(图2)。秋茄和木榄幼苗的叶片Pmax均随着生长光强的降低呈现先上升后下降的趋势,在45%光强下达到最大值,并且3个遮荫处理下Pmax均显著高于对照。相比对照,遮荫显著增加了木榄、银叶树和黄槿等3种红树植物幼苗叶片的表观量子效率(Φ),而对无瓣海桑、秋茄、桐花树、老鼠簕和卤蕨幼苗的Φ无明显影响。木榄、桐花树和卤蕨幼苗的叶片暗呼吸速率(Rd)随着生长光强的下降而显著降低,不同生长光强下无瓣海桑、秋茄、老鼠簕、银叶树和黄槿幼苗的Rd则均无显著差异。相比于本土红树种,无瓣海桑具有较高的Pmax和Rd。
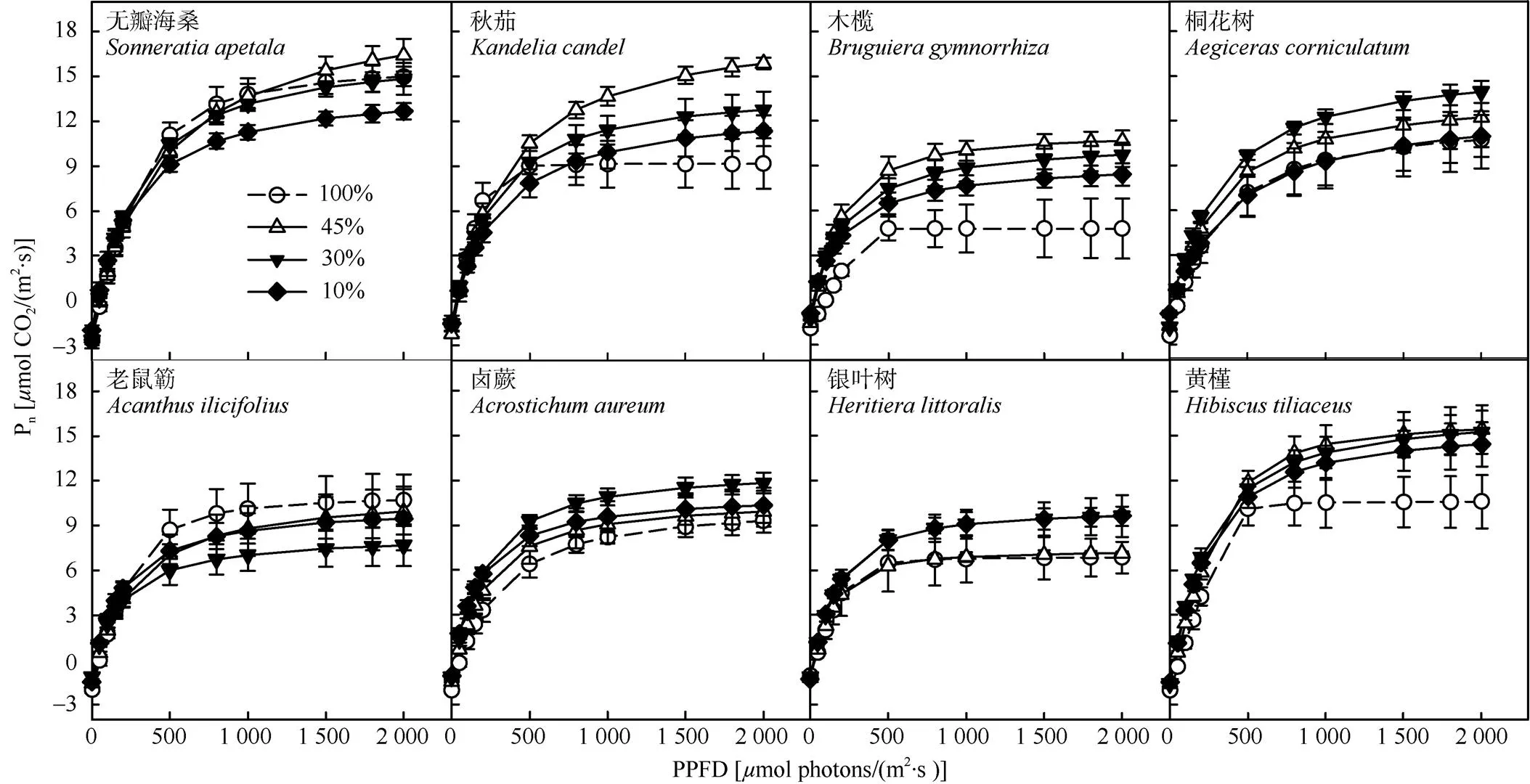
图1 8种红树幼苗叶片的光响应曲线。n=3~5; Pn: 净光合速率; PPFD: 光量子通量密度。
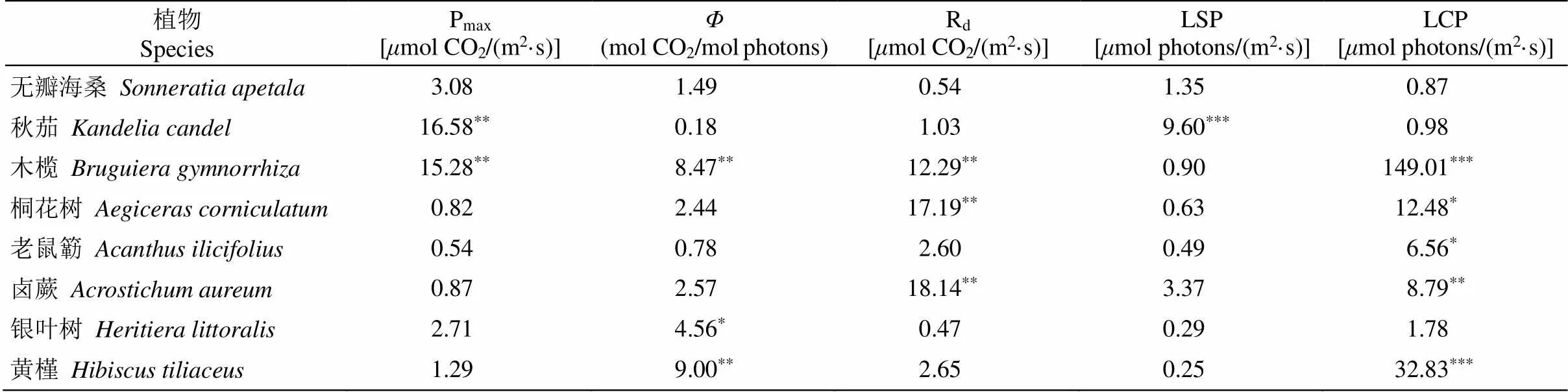
表1 生长光强对8种红树幼苗叶片光响应参数影响的单因素方差分析(F值)
*:<0.05; **:<0.01; ***:<0.001。Pmax: 最大净光合速率;: 表观量子效率; Rd: 暗呼吸速率; LSP: 光饱和点; LCP: 光补偿点。下同
*:<0.05; **:<0.01; ***:<0.001. Pmax: Maximum net photosynthetic rate;: Apparent quantum yield; Rd: Dark respiration rate; LSP: Light saturation point; LCP: Light compensation point. The same below
从图3可见,遮荫显著增加了秋茄幼苗叶片的LSP,45%、30%和10%光强下的LSP分别是对照的1.81、1.60和1.74倍,且遮荫处理间不存在显著差异。遮荫对其它红树植物幼苗的LSP均无显著影响。对于LCP而言,遮荫导致木榄、桐花树、老鼠簕、卤蕨和黄槿幼苗叶片的LCP显著降低,而对于无瓣海桑、秋茄和银叶树幼苗则无显著影响。
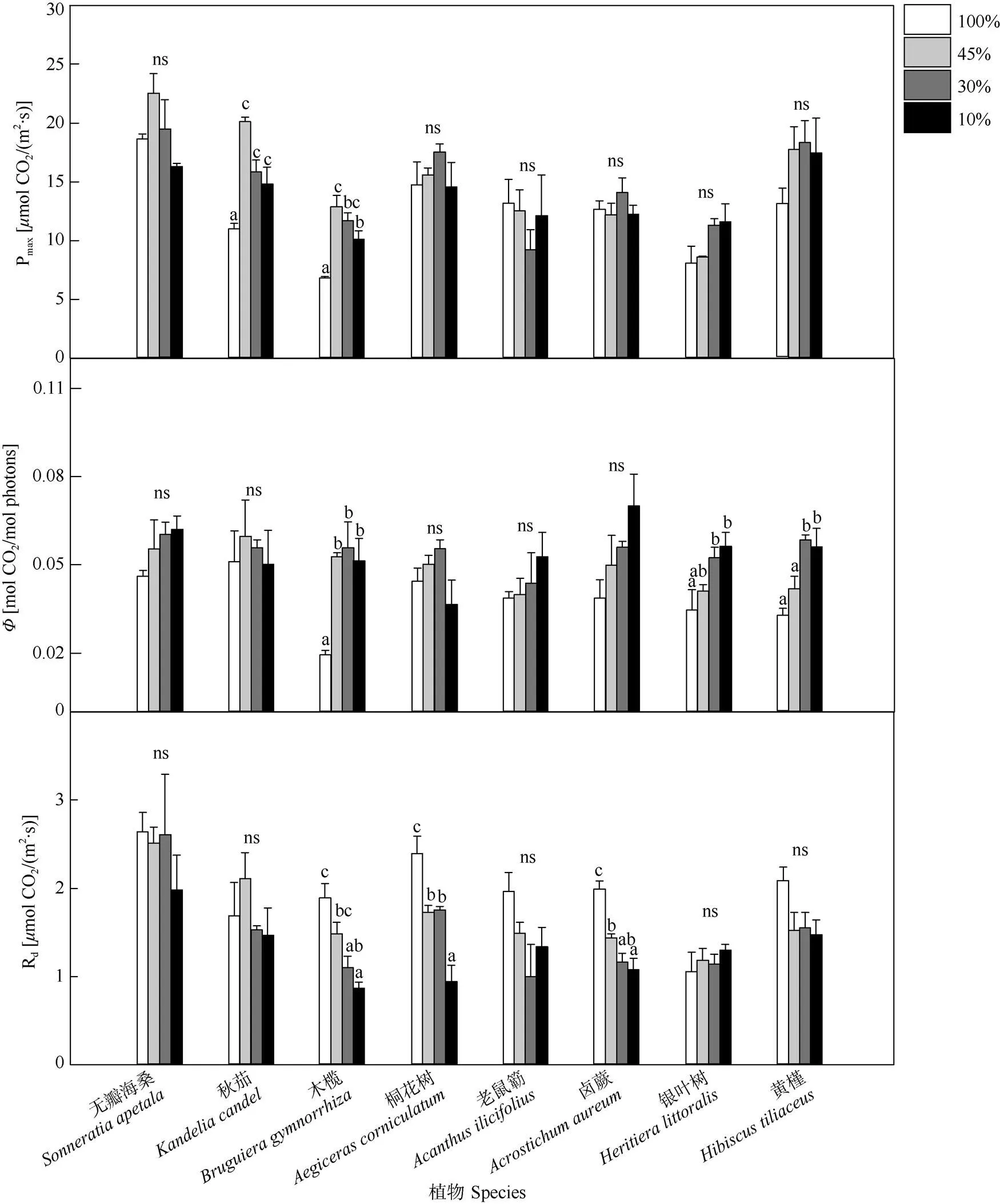
图2 不同光照条件下8种红树幼苗叶片的Pmax、Φ和Rd。n=3~5;同一物种柱上不同字母表示差异显著(P<0.05); ns: 不显著。下同
3 结论和讨论
植物叶片的光合响应特征是评价植物的光合能力及光能利用效率的有效途径[26]。表征植物利用光能的效率,其值越高表明植物叶片光能利用能力越强[18]。崔波等[27]报道白及()在遮荫下拥有更大的,表明其在遮荫条件下拥有更高的光能利用效率。本研究表明,除无瓣海桑、秋茄和桐花树外,其他5种红树植物幼苗在遮荫下的均表现出升高趋势,说明这5种红树植物幼苗对低光量子胁迫的耐性强于无瓣海桑、秋茄和桐花树, 能够在有限的光照条件下最大程度的利用光能,满足植物的生长需求。木榄、桐花树和卤蕨的幼苗Rd在遮荫下呈现出显著下降趋势,表明木榄、桐花树和卤蕨在遮荫条件下降低了能量的消耗,表现出较强的光合生理调节能力。这与前人对七子花()[10]和红毛五加()[28]的研究结果一致。
秋茄、木榄、卤蕨和2种半红树植物在遮荫下均表现出了更高的Pmax,表明其在遮荫下的光合能力更高,因此在自然生境中的强光照可能导致其发生光抑制。崔波等[27]报道白及在50%和70%的遮荫下拥有更大的Pmax,表明其在遮光下拥有更高的光合潜能。另有研究表明,金花茶()[12]、桃儿七()[29]、赤皮青冈()[15]及七子花[10]在轻度遮荫下的Pmax值升高。本研究表明,秋茄在45%的光照强度下Pmax值达到最大,木榄在45%和30%光照强度下的Pmax值显著大于其他两个处理,表明其在遮荫条件下具有更强的光合潜力,同时也可能说明木榄相比秋茄更加能适应弱光环境。彭逸生等[30]的研究表明,无瓣海桑相较于其他红树植物如桐花树表现出较弱的耐荫能力,刁俊明等[31]的研究表明无瓣海桑在荫蔽初期,弱光环境对幼苗的茎高、叶面积和叶片长宽比的增加有促进作用,而对长期生长在20%光照强度下的无瓣海桑幼苗的生长产生抑制作用,不利于幼苗发育。无瓣海桑在45%光照强度处理下净光合速率最高,随着生长光强降低,其Pmax呈下降趋势,说明弱光不利于无瓣海桑的光合作用,最终影响无瓣海桑的生长。
一般来说,阳生植物的LSP随着光照强度的增加而增加,阴生植物则与之相反[10]。秋茄在遮荫下具有更高的LSP,与金花茶的变化趋势一致[12],表明秋茄具有较好的耐荫能力。LCP的变化通常被认为是植物对于弱光环境的一种适应机制。有研究表明,遮荫下拥有更小LCP的植物可以减少消耗以便于更好地维持植物体稳态[12,32]。Zhang等[10]和Chai等[12]研究表明在遮荫条件下金花茶和七子花的LCP和Rd表现为下降趋势,这种调整可以使植物在弱光条件下减少光合产物的消耗并提高自身对弱光的利用能力。木榄、桐花树、老鼠簕、卤蕨和黄槿在遮荫下的LCP显著低于100%自然光照条件下的,表明这5种红树植物在遮荫下可以更好的提高对弱光的利用效率。同时, 10%光照强度下木榄和卤蕨的LCP也比其他红树植物低,表明其更加适应弱光环境。另外,与桐花树、无瓣海桑相比,老鼠簕的光饱和点和光补偿点都相对较低[33–34],表明老鼠簕比桐花树和上层树种更适应林下的荫生环境,这与本研究结果相似。
综上,从光合响应曲线及其特征参数来看,不同生长光强对红树植物光响应特征的影响因物种而异。秋茄、木榄、卤蕨和2种半红树植物在遮荫下均具有一定的利用弱光能力,特别是木榄和卤蕨对弱光环境表现出良好的适应性,因此适于林下种植。秋茄在遮荫下难以维持自身的物质生产与消耗的平衡,不适宜在郁闭度高的林下种植。老鼠簕通常作为红树林生态系统林下更新的主要树种,这说明其具有较好的耐荫性。我国红树林造林大多采用生长快、郁闭能力强的无瓣海桑,在对其进行科学的林分改造时,需要充分考虑改造树种在不同光照条件下的生长和生理生态特征。
[1] TANSLEY A G, FRITSCH F E. Sketches of vegetation at home and abroad: I. The flora of the Ceylon littoral [J]. New Phytol, 1905, 4(2/3): 27–55. doi: 10.1111/j.1469-8137.1905.tb05879.x.
[2] LIN P. Distribution of mangrove species [J]. Sci Silv Sin, 1987, 23(4): 481–490. [林鹏. 红树林的种类及其分布 [J]. 林业科学, 1987, 23(4): 481–490.]
[3] DASGUPTA R, SHAW R. Participatory Mangrove Management in a Changing Climate [M]. Tokyo: Springer, 2017: 2–3.
[4] MENG X W, XIA P, LI Z, et al. Mangrove degradation and response to anthropogenic disturbance in the Maowei Sea (SW China) since 1926 AD: Mangrove-derived OM and pollen [J]. Org Geochem, 2016, 98: 166–175. doi: 10.1016/j.orggeochem.2016.06.001.
[5] LIAO B W, ZHANG Q M. Area, distribution and species composition of mangroves in China [J]. Wetland Sci, 2014, 12(4): 435–440. [廖宝文, 张乔民. 中国红树林的分布、面积和树种组成 [J]. 湿地科学, 2014, 12(4): 435–440. doi: 10.13248/j.cnki.wetlandsci.2014.04.005.]
[6] LU Y P, XU W H, ZHANG Z M, et al. Gap analysis of mangrove ecosystem conservation in China [J]. Acta Ecol Sin, 2019, 39(2): 684– 691. [卢元平, 徐卫华, 张志明, 等. 中国红树林生态系统保护空缺分析 [J]. 生态学报, 2019, 39(2): 684–691. doi: 10.5846/stxb2018040 80794.]
[7] DUKE N C, MEYNECKE J O, DITTMANN S, et al. A world without mangroves? [J]. Science, 2007, 317(5834): 41–42. doi: 10.1126/science. 317.5834.41b.
[8] TAN S J, LI T, YU S R, et al. Effects of light intensity on growth and biomass allocation of seedlings of the eight mangrove species [J]. Ecol Sci, 2020, 39(3): 139–146. [谭淑娟, 李婷, 余素睿, 等. 光照强度对8种红树植物幼苗生长和生物量分配的影响 [J]. 生态科学, 2020, 39 (3): 139–146. doi: 10.14108/j.cnki.1008-8873.2020.03.019.]
[9] CHEN Z C, LIU X J, LIU C, et al. Responses of growth, photosynthesis and nonstructural carbohydrate ofvar.seedlings to shading and simulated sunfleck [J]. Chin J Ecol, 2017, 36 (4): 935–943. [陈志成, 刘晓静, 刘畅, 等. 锐齿栎幼苗生长、光合作用和非结构性碳对遮阴和模拟光斑的响应 [J]. 生态学杂志, 2017, 36(4): 935–943. doi: 10.13292/j.1000-4890.201704.010.]
[10] ZHANG Y F, CHEN C, JIN Z X, et al. Leaf anatomy, photosynthesis, and chloroplast ultrastructure ofseedlings reveal adaptation to light environment [J]. Environ Exp Bot, 2022, 195: 104780. doi: 10.1016/j.envexpbot.2022.104780.
[11] ZHANG W F, FAN D Y, XIE Z Q, et al. The seasonal photosynthetic responses of seedlings of the endangered plantto different growth light environments [J]. Biod Sci, 2005, 13(5): 387– 397. [张旺锋, 樊大勇, 谢宗强, 等. 濒危植物银杉幼树对生长光强的季节性光合响应 [J]. 生物多样性, 2005, 13(5): 387–397. doi: 10. 3321/j.issn:1005-0094.2005.05.003.]
[12] CHAI S F, TANG J M, MALLIK A, et al. Eco-physiological basis of shade adaptation of, a rare and endangered forest understory plant of Southeast Asia [J]. BMC Ecol, 2018, 18(1): 5. doi: 10.1186/s12898-018-0159-y.
[13] ZHANG J J, ZHU L, ZHANG X, et al. Photosynthetic performance and growth responses of(Decne.) L.H. Bailey (Aspara- gaceae) planted within poplar forests having different canopy densities [J]. BMC Ecol, 2020, 20(1): 25. doi: 10.1186/s12898-020-00294-7.
[14] WANG Y, TONG Y F, CHU H L, et al. Effects of different light qualities on seedling growth and chlorophyll fluorescence parameters of[J]. Biologia, 2017, 72(7): 735–744. doi: 10. 1515/biolog-2017-0081.
[15] XIA C, LI H, WANG P L, et al. Effects of shading on the photo- synthetic characteristics ofseedlings [J]. J CS Univ For Technol, 2021, 41(7): 72–79. [夏婵, 李何, 王佩兰, 等. 不同光照强度对赤皮青冈幼苗光合特性的影响 [J]. 中南林业科技大学学报, 2021, 41(7): 72–79. doi: 10.14067/j.cnki.1673-923x.2021.07. 009.]
[16] WU W M, LI Z J, LUO Q H, et al. Effects of soil water stress on light response curves of photosynthesis ofand[J]. Sci Silv Sin, 2007, 43(5): 30–35. [伍维模, 李志军, 罗青红, 等. 土壤水分胁迫对胡杨、灰叶胡杨光合作用-光响应特性的影响 [J]. 林业科学, 2007, 43(5): 30–35. doi: 10.3321/j.issn:1001-7488. 2007.05.005.]
[17] LONG S P, HUMPHRIES S, FALKOWSKI P G. Photoinhibition of photosynthesis in nature [J]. Annu Rev Plant Physiol Plant Mol Biol, 1994, 45(1): 633–662. doi: 10.1146/annurev.pp.45.060194.003221.
[18] YANG Z Q, HAN D, WANG X L, et al. Changes in photosynthetic parameters and antioxidant enzymatic activity of four tea varieties during a cold wave [J]. Acta Ecol Sin, 2016, 36(3): 629–641. [杨再强, 韩冬, 王学林, 等. 寒潮过程中4个茶树品种光合特性和保护酶活性变化及品种间差异 [J]. 生态学报, 2016, 36(3): 629–641. doi: 10. 5846/stxb201405130981.]
[19] LIAO B W, QIU F Y, TAM N F Y, et al. Study on the adaptability of mangroveseedlings to simulated tidal inundation [J]. J S China Agric Univ, 2009, 30(3): 49–54. [廖宝文, 邱凤英, 谭凤仪, 等. 红树植物秋茄幼苗对模拟潮汐淹浸时间的适应性研究 [J]. 华南农业大学学报, 2009, 30(3): 49–54. doi: 10.3969/j.issn.1001-411X. 2009.03.012.]
[20] LIAO B W, QIU F Y, ZHANG L E, et al. Effect of salinity on the growth and eco-physiological characteristics ofvar.seedlings [J]. Acta Ecol Sin, 2010, 30(23): 6363–6371. [廖宝文, 邱凤英, 张留恩, 等. 盐度对尖瓣海莲幼苗生长及其生理生态特性的影响 [J]. 生态学报, 2010, 30(23): 6363–6371.]
[21] LIAO B W, TIAN G H, YANG X B, et al. The analysis of natural regeneration and diffusion of the seedling ofin the Qi’ao Island, Zhuhai [J]. Ecol Sci, 2006, 25(6): 485–488. [廖宝文, 田广红, 杨雄邦, 等. 珠海淇澳岛无瓣海桑种苗天然更新与扩散分析 [J]. 生态科学, 2006, 25(6): 485–488. doi: 10.3969/j.issn.1008-8873. 2006.06.002.]
[22] CAI S H, LI T, ZHOU G X et al. Gas exchange characteristics in the mangrove associate[J]. Guihaia, 2016, 36(4): 397– 404. [蔡水花, 李婷, 周光霞, 等. 半红树植物黄槿的气体交换特性 [J]. 广西植物, 2016, 36(4): 397–404. doi: 10.11931/guihaia.gxzw 201403011.]
[23] HUANG L, ZHAN C A. Analysis on introduction and trial of man- groveon the seashore of east Guangdong [J]. For Sci Technol, 2003(5): 7–8. [黄陵, 詹潮安. 粤东沿海引种无瓣海桑试验研究 [J]. 林业实用技术, 2003(5): 7–8. doi: 10.13456/j.cnki. lykt.2003.05.002.]
[24] LAMBERS H, CHAPIN III F S, PONS T L. Plant Physiological Ecology [M]. 2nd ed. New York: Springer, 2008: 27–29.
[25] WALKER D A. Automated measurement of leaf photosynthetic O2evolution as a function of photon flux density [J]. Phil Trans Roy Soc B: Biol Sci, 1989, 323(1216): 313–326. doi: 10.1098/rstb.1989.0013.
[26] LIU H Y, YU H Y, XIONG K N, et al. Response characteristics of photosynthesis to light intensity of three non-wood forests tree species in karst habitat [J]. J S Agric, 2021, 52(9): 2507–2515. [刘海燕,喻阳华, 熊康宁, 等. 喀斯特生境3种经济林树种光合作用对光强的响应特征[J]. 南方农业学报, 2021, 52(9): 2507–2515. doi: 10.3969/j.issn.2095- 1191.2021.09.021.]
[27] CUI B, ZHOU Y R, WANG X M, et al. Study on the photosynthetic and physiological characteristics ofunder different light intensity [J]. J Henan Agric Univ, 2020, 54(2): 276–284. [崔波, 周一冉, 王喜蒙, 等. 不同光照强度下白及光合生理特性的研究 [J]. 河南农业大学学报, 2020, 54(2): 276–284. doi: 10.16445/j.cnki.1000- 2340.20200403.003.]
[28] CHEN Y F, HUANG X F, GU R, et al. Study on photosynthesis and physiological characteristics ofon different light intensity [J]. China J Chin Mat Med, 2018, 43(5): 926–933. [陈玉锋, 黄旭峰, 古锐, 等. 不同光照强度下红毛五加光合及生理特性研究 [J]. 中国中药杂志, 2018, 43(5): 926–933. doi: 10.19540/j.cnki. cjcmm.2018.0025.]
[29] GUO Q Q, LI H E, GAO C, et al. Leaf traits and photosynthetic characteristics of endangered(Royle) Ying under different light regimes in southeastern Tibet Plateau [J]. Photosynthetica, 2019, 57(2): 548–555. doi: 10.32615/ps.2019.080.
[30] PENG Y S, DIAO J M, CHEN G Z, et al. Ecophysiological response of three mangroves’ seedlings under the shade canopies of mangrove plantations [C]// Proceedings of the 17th Symposium of Guangdong Botanical Society. Shenzhen: Guangdong Botanical Society, 2008: 3. [彭逸生, 刁俊明, 陈桂珠. 3种红树植物幼苗对人工林下遮光条件的生理生态响应研究[C]//广东省植物学会第十七期学术研讨会论文集. 深圳: 广东省植物学会, 2008: 3.]
[31] DIAO J M, CHEN G Z. Effect of light intensity on growth and photo- synthetic properties ofseedling [J]. For Res, 2008, 21(4): 486–492. [刁俊明, 陈桂珠. 光强对无瓣海桑幼苗的生长和光合特性的影响 [J]. 林业科学研究, 2008, 21(4): 486–492. doi: 10. 3321/j.issn:1001-1498.2008.04.010.]
[32] MIAO Z W, XU M, LATHROP R G JR, et al. Comparison of the-ccurve fitting methods in determining maximum ribulose 1,5-bisphosphate carboxylase/oxygenase carboxylation rate, potential light saturated electron transport rate and leaf dark respiration [J]. Plant Cell Environ, 2009, 32(2): 109–122. doi: 10.1111/j.1365-3040.2008.01900.x.
[33] CHEN L Z, TAM N F Y, HUANG J H, et al. Comparison of ecophy- siological characteristics between introduced and indigenous mangrove species in China [J]. Estuar Coast Shelf Sci, 2008, 79(4): 644–652. doi: 10.1016/j.ecss.2008.06.003.
[34] HAN S M, LI N Y, HE P, et al. Photosynthetic characteristics of introduced and indigenous mangrove seedlings in China [J]. Acta Bot Boreali-Occid Sin, 2010, 30(8): 1667–1674. [韩淑梅, 李妮亚, 何平, 等. 引种红树与中国乡土红树幼苗光合特性研究 [J]. 西北植物学报, 2010, 30(8): 1667–1674.]
Light Response Characteristics of Seedlings Leaves of Eight Mangrove Species Under Different Light Intensities
ZHU Yimin1,2,3, LI Ting1,2,3, SUN Dangge1,2,3, YE Wanhui1,2,3,4, SHEN Hao1,2,3,4*
(1. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems, Guangdong Provincial Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China; 2. South China National Botanical Garden, Guangzhou 510650, China; 3.University of Chinese Academy of Sciences,Beijing 100049, China; 4. Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511458, China)
Mangrove species are a group of trees, shrubs or herbs that grow in intertidal zones of tropical and subtropical coasts. They are generally categorized into true mangrove and semi-mangrove species according to their distribution. Understanding the eco-physiological strategies of light adaptation of mangrove species is of important theoretical significance to revealing the community assembly mechanisms of mangrove forests, and can provide scientific guidance for the reconstruction and transformation of mangrove forests. In order to reveal the physiological and ecological strategies to light conditions of mangrove plant species and provide theoretical guidance for optimal assembly of mangrove community structure, the light response characteristics in leaves of eight mangrove seedlings, such as,,,,,,and, grown under different light intensities, including 100%, 45%, 30%, and 10% of natural sunlight, were studied using shading control experiment. The results showed thatthe effects of growth light intensities on the photosynthetic light response characteristics were different among mangrove species.Shading significantly increased the maximum net photosynthetic rate (Pmax) ofand, but had no significant effects on Pmaxof other mangrove species.had a higher Pmaxunder 45% light intensity treatment, whilehad higher Pmaxunder 45% and 30% light intensities. With the decrease of growth light intensity, the light saturation point of leaves ofseedlings significantly increased, the light compensation points of,andappeared a decreasing trend, the apparent quantum yield ofandshowed an increasing pattern, and the dark respiration rate ofanddecreased. Therefore, it was suggested that,andare suitable for planting under forest with high canopy density, while,,,andare suitable to be planted as upper layer species in mangrove forests or planted under forest with low canopy density.
Mangrove species; Shading; Photosynthesis; Light intensity
10.11926/jtsb.4677
2022-05-22
2022-07-01
广东省林业科技创新项目(2017KJCX036, 2019KJCX015);南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0408)资助
This work was supported by the Project for Forestry Science and Technology Innovation in Guangdong (Grant No. 2017KJCX036, 2019KJCX015), and the Key Special Project for Introduced Talents Team of Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou) (Grant No. GML2019ZD0408).
朱一民(1998年生),男,硕士研究生,研究方向为植物生理生态学。E-mail: zhuyimin@scbg.ac.cn
* 通讯作者 Corresponding author. E-mail: shenhao@scbg.ac.cn
猜你喜欢
热带林业(2023年3期)2023-10-30 10:38:34
小猕猴学习画刊(2022年4期)2022-05-05 01:52:59
广东第二课堂·小学(2021年10期)2021-12-03 04:35:50
世界科学技术-中医药现代化(2021年12期)2021-04-19 12:32:10
海洋通报(2020年5期)2021-01-14 09:27:04
天然产物研究与开发(2016年6期)2016-06-05 10:29:31
艺术评鉴(2016年13期)2016-05-30 16:19:05
发明与创新(2015年6期)2015-12-26 11:17:18
发明与创新·中学生(2015年6期)2015-06-01 00:40:24
发明与创新(2015年22期)2015-04-17 23:22:59
