
Hypoglycemic effect and the mechanism of action of a polysaccharide from sweet corncob in a high-fat diet and streptozotocin-induced diabetic mice
2024-01-24 01:11XinWangWeiyeXiuYeHanZhiliWangYuLuoYongqiangMa
食品科学与人类健康(英文) 2024年3期
Xin Wang, Weiye Xiu, Ye Han, Zhili Wang, Yu Luo, Yongqiang Ma
Heilongjiang Provincial Key Laboratory of Cereals and Comprehensive Processing of Cereal Resources, College of Food Engineering, Harbin University of Commerce, Harbin 150028, China
Keywords: Sweet corn cob polysaccharide Type 2 diabetes Signal pathway 5’-AMP-activated protein kinase (AMPK)
ABSTRACT Type 2 diabetes mellitus (T2DM) is a metabolic disease caused by a glycolipid metabolism disorder and islet β-cell dysfunction. SCP-80-I is a biologically active water-soluble polysaccharide isolated from sweet corncob, an agricultural byproduct. The hypoglycemic effects of SCP-80-I on T2DM mice and its mechanisms were investigated in this study. SCP-80-I was found to signif icantly reduce blood glucose and lipid deposition levels in T2DM mice, as well as decrease serum leptin and increase adiponectin secretion. Interestingly, real time-polymerase chain reaction(RT-PCR) and Western blotting results revealed that SCP-80-I could regulate the expression of several glycolipid metabolisms and insulin secretion genes and proteins, including 5’-AMP-activated protein kinase (AMPK),carnitine palmitoyltransferase I (CPT1), and acetyl coenzyme A carboxylase (ACC) in the liver and AMPK, sirtuin 1 (Sirt1), peroxisome proliferator-activated receptorycoactivator-1 (PGC-1α), and uncoupling protein 2 (UCP2)in the pancreas. To have a hypoglycemic effect, SCP-80-I regulated glycolipid metabolism and islet cell function in the liver by regulating the AMPK/ACC/CPT1 signaling pathway and the AMPK/Sirt1/PGC-1α and AMPK/Sirt1/UCP2 signaling pathways. These f indings improve our understanding of polysaccharides derived from sweet corncob and the use of SCP-80-I in the production of hypoglycemic foods.
1. Introduction
Type 2 diabetes mellitus (T2DM) is a metabolic disease characterized by islet β-cell dysfunction and insulin resistance syndrome[1]. The typical symptom of T2DM is hyperglycemia. In recent years, erratic working and living habits have considerably increased cases of T2DM[2]. T2DM is a metabolic disorder that affects people the most. T2DM treatment mainly involves controlling blood glucose and blood lipids, reducing metabolic disorders, and enhancing glycolipid metabolism[3]. T2DM is primarily treated with insulin injections and oral insulinotropic drugs to increase insulin levels in the body. To lower blood glucose levels, many drugs, including pioglitazone, acarbose, repaglinide, and sitagliptin,promote insulin sensitivity and regulate signaling pathways related to insulin secretion[4]. These medications, however, have negative effects on the gastrointestinal tract and the nervous system, causing dry cough, angioedema, hyperpotassemia, and headache[5-6]. Hence, many researchers have focused on natural products with hypoglycemic effects.
Polysaccharides are polymeric carbohydrates formed by repeating sugar units. The basic components of a polysaccharide are monosaccharides, which are connected by glycosidic bonds[7-8].Polysaccharides have hypoglycemic effects[9-10]. A polysaccharide from theGynostemma pentaphyllumherb exhibits hypoglycemic effects by inducing antioxidation and anti-inflammation[11]. In T2DM mice, a polysaccharide extracted fromCordyceps militarisdemonstrated antioxidant and hypoglycemic effects[12]. Moreover,polysaccharides exert hypoglycemic effects by regulating the expression of related genes or pathways. A polysaccharide fromCyclocarya paliuruscan relieve oxidative stress damage associated with diabetes by regulating the genes and pathways related to mitochondria and fatty acid metabolism[13]. Some studies have used a similar method to show that proteoglycans fromGanoderma lucidumand polysaccharides fromSiraitia grosvenoriihave antihyperglycemic effects[14-15].
Owing to its sweet and crisp flavor, sweet corn (Zea maysL.saccharataSturt) is popular among consumers. It has been accompanied by the production of numerous agricultural byproducts,including sweet corn cobs. Currently, a large amount of sweet corn cobs are discarded, burned, or used as agricultural fertilizer, all of which are underutilized. In a previous study, we successfully extracted and purified a water-soluble polysaccharide (SCP-80-I) from sweet corn cobs. SCP-80-I molecular weight, monosaccharide components,and structure have also been determined[16].
We investigated whether SCP-80-I improves glycolipid metabolism and islet β-cell function by regulating the target gene pathway. We looked at how SCP-80-I affected blood glucose levels, body weight, tissue morphology, and insulin release in T2DM mice. We also looked at the target genes and signaling pathways involved in liver glycolipid metabolism and pancreatic mitochondrial function in T2DM mice after SCP-80-I administration. In addition, we discovered a link between the liver and pancreatic target gene pathways and the hypoglycemic effect of SCP-80-I. We aimed to determine the mechanism by which SCP-80-I exerts an anti-hyperglycemic effect. To this end,morphological, physiological, histological, genetic (gene and protein level), and metabolic approaches were used.
2. Materials and methods
2.1 Materials and reagents
Sweet corncob (Z. maysL.saccharataSturt) was obtained from HaoWei Agriculture Co., Ltd. (Harbin, China). Streptozotocin(STZ) was purchased from Sigma-Aldrich Chemical Co. (St. Louis,MO, USA). Jiancheng Bioengineering Institute provided the insulin ELISA, leptin ELISA, adiponectin, total cholesterol (TC), total glycerol (TG), low-density lipoprotein cholesterol (LDL-C), highdensity lipoprotein cholesterol (HDL-C), liver glycogen, superoxide dismutase (SOD), catalase (CAT), glutathione peroxidase (GSH-Px),and malondialdehyde (MDA) assays were purchased from Jiancheng Bioengineering Institute (Nanjing, China). Metformin was purchased from Shengjitang Pharmaceutical Co., Ltd. (Guiyang, China). The TRIpure total RNA extraction reagent, EntiLink™ 1stStrand cDNA synthesis kit, and the EnTurbo™ SYBR green PCR SuperMix were purchased from ELK Biotechnology Co., Ltd. (Wuhan, China). The primary and secondary antibodies for p-AMPK, AMPK, ACC, CPT1,Sirt1, PGC-1, and UCP2 were purchased from ASPEN Biotechnology Co., Ltd. (Wuhan, China). The other chemical reagents were of analytical grade.
2.2 Preparation of the polysaccharides
The polysaccharides were prepared as described in our previous study. SCP-80-I was obtained from the degreased parts of sweet corn cobs and stored at –20 °C. The purity of SCP-80-I was 97.3% with the composition of glucose, and the molecular weight was 173.780 kDa.The structural characterization of SCP-80-I is shown in Fig. S1[17].
2.3 Animals and experimental design
The ICR male mice (n= 60; weight = (20 ± 2) g) were provided by Changchun Yisi Experimental Animal Center (Production license number: SCXK(Ji)-2018–0007, Certificate Number: 202000034164).The mice were housed in a controlled environment at a temperature of(23 ± 1) °C and humidity of (60 ± 5)%, following a 12 h/12 h light-dark cycle. All mice were provided a normal diet for acclimation for 7 days.
The mice were divided into 2 groups after 7 days of adaptive breeding. To obtain the high-fat model mice, the normal control group(n= 10) was given a normal diet for 28 days, while the other group(n= 50) was given a high-fat diet (Boaigang Biological Technology Co., Ltd., Beijing, China). Second, STZ (0.1 mol/L, 100 mg/kg bw)was used to induce the high-fat model mice, while physiological saline was used to induce the normal mice. The fasting blood glucose was determined after induction with STZ for 3 and 7 days. The mice with blood glucose above 11.1 mmol/L were considered the T2DM model mice and were used in subsequent experiments[18]. All T2DM model mice were divided into 5 groups; the administration is shown in Fig. 1.

Fig. 1 Animals and the experimental design.
After intragastric administration for 21 days, all mice were starved overnight and sacrificed after extracting blood samples by an orbital puncture. The tissues from the liver and pancreas were collected.
2.4 Oral glucose tolerance test (OGTT)
All groups of mice underwent the OGTT after 12 h of fasting on day 21. The mice were administered 2.5 g/kg glucose intragastrically.Blood was collected from the tail vein of the mice for analyzing glucose levels at 0, 30, 60, and 120 min after glucose administration.The area under the curve (AUC) was calculated as follows:
whereA,B,C, andDrepresent the blood glucose values determined at 0, 30, 60, and 120 min, respectively[19].
2.5 Biochemical analysis
The levels of TC, TG, LDL-C, HDL-C, leptin, and adiponectin in the serum, the level of TC and TG, the content of hepatic glycogen in the liver, the activity of SOD, CAT, and GSH-Px. Additionally,the fasting blood glucose and insulin content of all groups of mice were measured to calculate homeostasis model assessment-insulin resistance (HOMA-IR). The formula was as follows:
2.6 Morphological analysis
2.6.1 Histopathological examination
The morphological changes in the liver and pancreas were observed by hematoxylin and eosin (H&E) staining. Liver and pancreatic tissues were removed from mice in each group, fixed in 4% paraformaldehyde for 24 h, embedded in paraffin, and cut into thin sections (4 μm). After H&E staining, the tissue sections were examined by light microscopy (CX-21, OLYMPUS, Japan).
2.6.2 TUNEL staining assay
The terminal deoxynucleotidyl transferase end labeling(TUNEL) assay was performed in the pancreas using a TUNEL kit(In SituCell Death Detection Kit, Roche, Madrid, Spain), and the pancreatic tissue was embedded in paraffin after being fixed with 4%paraformaldehyde overnight. The sample was then examined under a light microscope (CX-21, OLYMPUS, Japan) to calculate the ratio of TUNEL positive rate of protein expression[20].
2.6.3 Immunohistochemical (IHC) assay
The amylin deposition was determined by the IHC assay. Briefly,the pancreatic tissue embedded in paraffin was cut into thin sections(50 μm), and endogenous peroxidase was blocked with H2O2(3%)after the antigen was repaired by microwave heating. After that,primary antibodies (1:400, 22305–1-AP, Proteintech Group, Inc.,USA) were added, and the tissue sections were incubated at 4 °C overnight, followed by the addition of secondary antibodies (1:200,AS-1107, ASPEN Biotechnology Co., Ltd., Wuhan, China) at 37 °C for 50 min. The sections were then stained with a diaminobenzidine(DAB) kit (ZLI-9033; Beijing Zhongshan Golden Bridge Biotechnology Co., Ltd., Beijing, China) and examined with a light microscope (CX-21, OLYMPUS, Japan) to assess the content and distribution of specific antibodies in pancreatic β-cells[21].
2.6.4 Mitochondrial morphology analysis
The mitochondrial morphological characteristics of the islet β-cells were observed by transmission electron microscopy (HT7700, Hitachi,Japan) after the pancreatic tissue was fixed in 2.5% glutaraldehyde and cut into thin sections (60 nm) using an ultramicrotome (Leica EM UC7, LEICA, Germany).
2.7 Real-time (RT)-PCR analysis
Total RNA from the liver and pancreas of mice was extracted using the TRIpure total RNA extraction reagent (ELK Biotechnology Co., Ltd., China), and cDNA was synthesized using the EntiLinkTM1ststrand cDNA synthesis kit (ELK Biotechnology, EQ003, China).RT-PCR was performed using the EnTurboTMSYBR Green PCR SuperMix (ELK Biotechnology, EQ001, China) and the QuantStudio 6 Flex PCR System (Life Technologies, USA). The reaction conditions were as follows: 95 °C for 180 s, 40 cycles for 10 s at 95 °C, 30 s at 58 °C, and 30 s at 72 °C. The relative expression of the mRNA was standardized toGAPDH. The data were measured using the 2–ΔΔCtmethod. The primer sequences are provided in Table 1[22].
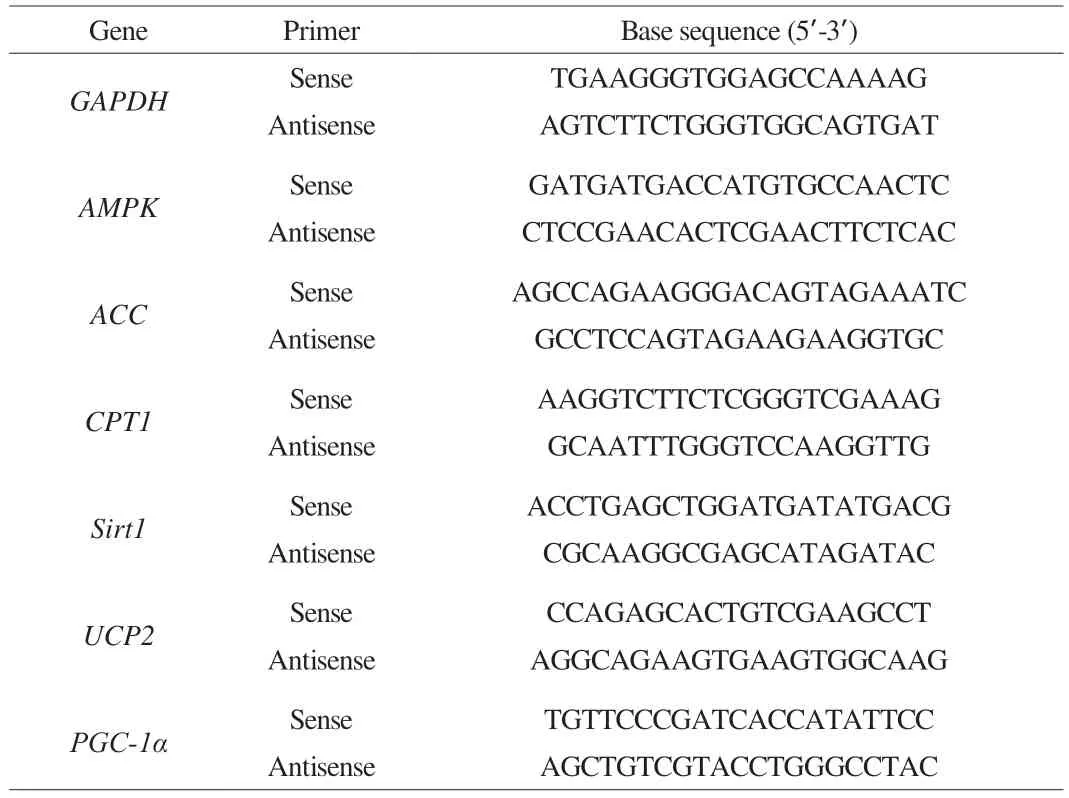
Table 1 Primer sequences for quantitative real-time PCR
2.8 Western blotting analysis
The expression of the monophosphate-activated protein kinase(AMPK), phosphorylated AMPK protein (p-AMPK), carnitine palmitoyltransferase 1 (CPT1), and acetyl-coenzyme a carboxylase(ACC) in the liver tissue and AMPK, p-AMPK, sirtuin1 (Sirt1),and proliferator-activated and receptor γ coactivator-1α (PGC-1α)levels in pancreatic tissues were determined by Western blotting.The tissues were weighed and cut into small pieces. Then, they were homogenized and placed in an ice bath for 30 min. The homogenate was centrifuged at 12 000 ×gfor 5 min at 4 °C. The supernatant was collected using a BCA kit (Jiancheng Biological Engineering Institute,Nanjing, China) to determine the protein concentration. The total protein was separated by performing SDS-PAGE and transferred onto a polyvinylidene fluoride (PVDF) membrane. The membrane was blocked for 1 h at room temperature. Then, the blocking buffer was removed, and the primary antibody (AS1061, ASPEN Biotechnology Co., Ltd., Wuhan, China) was added and incubated at 4 °C overnight.Then, after washing the membranes three times with TBS + 0.1%Tween (TBST) for 5 min each time, the secondary antibody (AS1058,ASPEN Biotechnology Co., Ltd., Wuhan, China) was added at room temperature for 30 min and washed 4 times with TBST. Finally,the electro-chemi-luminescence (ECL) reagent was added to the membranes, which were then analyzed by a chemiluminescence analyzer. The protein expression was determined by the AlphaEaseFC software system (AlphaImager System, Alpha Innotech, San Leandro,CA, USA)[23].
2.9 Statistical analysis
All data were represented as the mean ± standard deviation (SD).Significant differences among and between groups were determined by performing an analysis of variance (ANOVA) with Tukey’s test for multiple comparisons. The statistical analyses were conducted using the SPSS 17.0 (SPSS, Chicago, IL, USA) software, and the figures were plotted with Origin 8.0 (OriginLab Corp., MA, USA).
3. Results
3.1 Growth status, fasting blood glucose, and the OGTT
After intraperitoneal injection of STZ, the T2DM mice were found to have less weight, less glossy coats, and more food intake,and the fasting blood glucose levels also increased. The mice in the NC group showed stable weight and normal blood glucose levels.The weight of the mice in the SCP-H group was (36.00 ± 2.10) g after STZ administration for 21 days and (31.00 ± 2.80) g in the DC group (P< 0.05, Fig. 2A). Meanwhile, Fig. 2B depicts the food intake per mouse per day. T2DM mice had significantly higher daily food intake than the NC group, which gradually decreased after SCP-80-I treatment. The daily intake of mice in the SCP-H group was not significantly different from that of the NC group at 21 days(P> 0.05), indicating that SCP-80-I could alleviate the phenomenon of T2DM overfeeding.
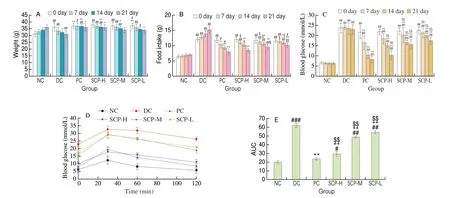
Fig. 2 Growth status, blood glucose levels, and OGTT in mice. (A) The weight of the mice. (B) The food intake per mouse per day. (C) The blood glucose in each group of mice. (D and E) The OGTT and AUC of the mice. #P < 0.05, ##P < 0.01, ###P < 0.001 compared to NC; *P < 0.05 and **P < 0.01 compared to DC;$P < 0.05 and $$P < 0.01 compared to PC. The same below.
Also, the fasting blood glucose levels decreased from(21.80 ± 1.42) mmol/L to (14.99 ± 1.17) mmol/L in the SCP-H group(Fig. 2C). The results indicated that the weight increased and fasting blood glucose levels decreased after treatment with SCP-80-I. Hence,SCP-80-I could prevent weight loss and the increase in blood glucose levels due to diabetes.
The OGTT was used to assess glucose absorption and metabolism ability. We calculated the AUC to investigate the hypoglycemic function of SCP-80-I. We discovered that the blood glucose levels in mice increased and then decreased after being given a glucose solution. We found the blood glucose levels in the mice decreased significantly (P< 0.01) 60 min after administering SCP-80-I; also,the AUC was smaller. Moreover, the decrease in blood glucose levels showed a dose-effect relationship (Figs. 2D-E). A similar result was also found for the mice in the PC group. We concluded that SCP-80-I could ameliorate glucose tolerance in diabetic mice.
3.2 Serum biochemical indices
The parameters such as TC, TG, HDL-C, and LDL-C are commonly used to evaluate the lipid circulation state in the body.The level of TC, TG, and LDL-C increased, and the level of HDL-C decreased in the T2DM mouse serum. After administering SCP-80-I,the pathological phenomenon was relieved, especially in the SCP-H group (Figs. 3A-D). Meanwhile, there was no significant difference in TC, HDL-C, and LDL-C indexes between the SCP-H and NC groups(P> 0.05), indicating that the mice were better restored regarding serum lipid metabolism. SCP-80-I significantly reduced lipid metabolism disorder in the mice (P< 0.01).
The levels of leptin, adiponectin, and HOMA-IR in the serum reflect metabolism and insulin resistance in T2DM mice[24]. The serum leptin level significantly increased, and the adiponectin level decreased in T2DM mice. This indicated some metabolic disturbance and insulin resistance in the T2DM mice. After 21 days of intragastric administration of SCP-80-I, the level of leptin decreased significantly from (0.62 ± 0.03) ng/mL in the DC group to (0.43 ± 0.03) ng/mL in the SCP-H group (Fig. 3E). We also discovered no significant difference between the SCP-M, SCP-H, and NC groups (P> 0.05).The level of adiponectin increased from (124.27 ± 9.80) ng/mL in the DC group to (162.55 ± 8.03) ng/mL in the SCP-H group (P< 0.01,Fig. 3F). Moreover, HOMA-IR in the serum decreased significantly in the SCP-H group compared to that in the DC group (6.10 ± 1.05vs. 25.20 ± 2.85,P< 0.01, Fig. 3G). These results were similar to those of the PC group. Thus, the metabolic disturbance and insulin resistance decreased in T2DM mice after treatment with SCP-80-I.
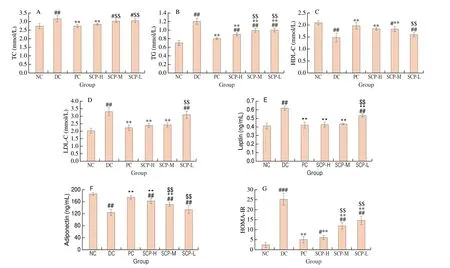
Fig. 3 Biochemical indices in the serum of mice. (A) TC, (B) TG, (C) HDL-C, (D) LDL-C, (E) Leptin, (F) Adiponectin; (G) HOMA-IR.
3.3 Hepatic glycolipid metabolism
The TC and TG levels and the hepatic glycogen content were used to evaluate the hepatic glycolipid metabolism in T2DM mice. The TC and TG levels were (0.23 ± 0.02) and (0.11 ± 0.01) mmol/L in the SCP-H group, while they were (0.33 ± 0.02) and (0.18 ± 0.01) mmol/L in the DC group (Figs. 4A and B). The results showed that the level of TC and TG in the T2DM mice decreased significantly after treatment with SCP-80-I for 21 days (P< 0.05), and there was no significant difference in the TG level in SCP-H and NC (P> 0.05). The content of hepatic glycogen increased from (4.02 ± 0.35) mg/g in the DC group to (7.77 ± 0.23) mg/g in the SCP-H group (P< 0.05, Fig. 4C).The results for the SCP-H group were similar to that for the PC group(P> 0.05). This indicated that SCP-80-I could reduce blood glucose levels by enhancing glycolipid metabolism in the liver.
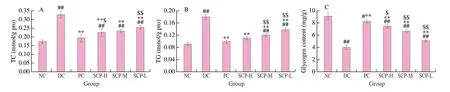
Fig. 4 Lipid deposition and glycogen content in the mouse liver. (A) TC level, (B) TG level, (C) glycogen content.
3.4 Oxidative stress in the pancreas
The activity of SOD, CAT, and GSH-Px and the contents of MDA were determined to evaluate the oxidative stress in T2DM mice. We found that the level of SOD, CAT, and GSH-Px in pancreatic tissues in T2DM mice decreased, while the level of MDA increased significantly (P< 0.01). Moreover, there was no remarkable difference in NC and SCP-H groups in the GSH-Px level (P> 0.05).This pathological phenomenon was relieved significantly 21 days after the intragastric administration of SCP-80-I (P< 0.05), indicating the positive influence of SCP-80-I in the pancreas of T2DM mice (Fig. 5).
3.5 Morphological analysis
We compared the morphology of the liver and pancreas of the mice in each group. The hepatic cells in the DC group were partly disordered, swollen, deformed, and fatty degenerated, while there were no abnormalities in the hepatic cells of the NC group. After treatment with SCP-80-I for 21 days, the structure became clearer,and lipid accumulation decreased in the hepatic cells. The effect of the treatment with SCP-80-I was similar to that after the treatment with metformin in the PC group (Fig. 6A).
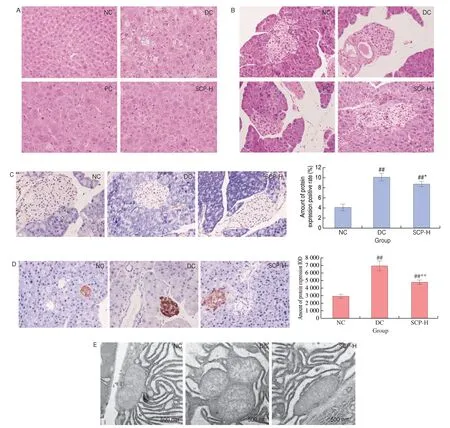
Fig. 6 Morphological analysis. (A) Histopathological examination of the livers in T2DM mice was conducted by H&E staining (400×). (B) Histopathological examination of the pancreas in T2DM mice was conducted by H&E staining (400×). (C) The pancreatic TUNEL staining assay of mice (400×) and the positive rate of protein expression. (D) The results of Amylin-IHC staining of the pancreas (400×) and positive rate of protein expression in mice. (E) Transmission electron microscopy of mitochondria in the islet β-cells. The scale bar represented a length of 500 nm.
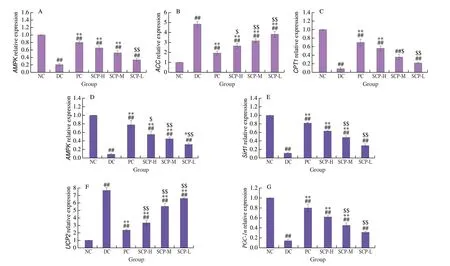
Fig. 7 Relative expression of the hepatic and pancreatic genes in mice. The expression of the (A) AMPK, (B) ACC, and (C) CPT1 genes in the liver and the(D) AMPK, (E) Sirt1, (F) UCP2, and (G) PGC-1α genes in the pancreas.
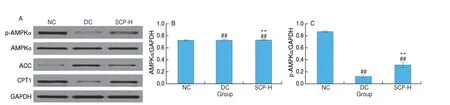
Fig. 8 Relative expression of hepatic and pancreatic proteins in mice. The expression levels of the proteins in the (A) liver and (G) pancreas; the relative density of (B) AMPKα, (C and D) p-AMPKα, (E) ACC, and (F) CPT1 in the liver and (H) AMPKα, (I and J) p-AMPKα, (K) Sirt1, and (L) PGC-1α in the pancreas. The relative density is expressed as a ratio shown in the figure.
We discovered that the islet β-cells of the NC mice had a homogeneous arrangement with compact intercellular spaces in the pancreas. The cells in the DC group atrophy show focal necrosis and are disordered. The lesion was relieved by administering metformin or SCP-80-I intragastrically. After 21 days of SCP-80-I administration,the cells were more uniformly distributed with clearer edges(Fig. 6B). These findings suggested that SCP-80-I could alleviate hepatic and pancreatic tissue damage caused by diabetes in mice.
The TUNEL assay was performed to determine the positive expression rate of apoptotic cells in the pancreas. The TUNEL assay has high sensitivity and can also detect a few apoptotic cells. The pancreatic TUNEL staining of the mice is shown in Fig. 6C. The islet β-cells in the NC group had fewer apoptotic cells (stained brown-yellow),while there were more apoptotic cells in the DC group. The apoptosis rate decreased after the administration of SCP-80-I(400 mg/(kg∙day)) for 21 days. The results of image quantification showed that the number of positive apoptotic cells in the SCP group decreased by 13.29% relative to the number of apoptotic cells in the DC group (P< 0.05).
Amylin, also known as a pancreatic amyloid peptide, builds up on the surface of islet β-cells, causing apoptosis and dysfunction[25].The IHC method was used to assess the distribution and content of specific antibodies. Fig. 6D shows the results of Amylin-IHC staining of the pancreas from mice. Amylin was distributed differently in different cells and positive apoptotic cells were found in pancreatic islets. Therefore, the protein levels expressed in the various groups differed significantly. The area with amylin deposition was lighter in the NC group than in the DC group. This pathological phenomenon was relieved after the oral administration of SCP-80-I(400 mg/(kg·day)). Moreover, the results showed that the deposition of amylin in the β-cells of the SCP-H group decreased by 30.86%compared to that in the β-cells of the DC group (P< 0.05). We found that SCP-80-I could significantly decrease the amylin expression in the islet β-cells of mice.
The mitochondria of the islet β-cells play an important role in the glucose-stimulated insulin secretion (GSIS) process. Damage to the GSIS process in T2DM is mainly related to mitochondrial dysfunction. We observed the mitochondria of the islet β-cells under a microscope. The mitochondria in the islet β-cells of the NC group were intact, granular, and had no pathological dilation in the extracellular compartment; additionally, the mitochondrial cristae were clear. The endoplasmic reticulum was dense and orderly,and there was no edema or aggregation. In the DC group, some edemas were observed in the mitochondria; the mitochondrial cristae were shorter and showed displacement. The mitochondrial cristae were randomly arranged and were fewer. The swelling in the mitochondria decreased after the intragastric administration of SCP-80-I. Also, their order and structure improved (Fig. 6E).The results indicated that SCP-80-I could control experimentallyinduced hyperglycemia and might function by relieving the shape and function of the islet cells.
3.6 RT-PCR analysis
SCP-80-I affected the expression of theAMPK,CPT1, andACCgenes in the liver and theAMPK,Sirt1,PGC-1α, andUCP2genes in the pancreas of diabetic mice.
In the liver, the expression ofAMPKandCPT1genes decreased,while that of theACCgene increased in the DC group. After treatment with different doses of SCP-80-I, the expression of theAMPKandCPT1genes was upregulated, and the expression of theACCgene was downregulated significantly (P< 0.01, Figs. 7A-C).
In the pancreas, the expression of theAMPK,Sirt1, andPGC-1αgenes decreased, while the expression of theUCP2gene increased in the DC group. Thus, theAMPK,Sirt1, andPGC-1genes were upregulated, and theUCP2gene was downregulated significantly after the treatment with SCP-80-I in T2DM mice (P< 0.01). The results also showed that SCP-80-I, like metformin, could relieve T2DM at the gene level (Figs. 7D-G). Thus, SCP-80-I can regulate the AMPK/ACC/CPT1 signaling pathway in the liver. Moreover,SCP-80-I can also regulate the AMPK/Sirt1/PGC-1 and AMPK/Sirt1/UCP2 signaling pathways in the pancreas.
3.7 Western blot assay
After 21 days of administration, SCP-80-I significantly increased p-AMPK and CPT1 protein expression while decreasing ACC protein expression in the liver. The protein p-AMPK is a phosphorylated and activated form of AMPK. The increase in AMPK activity in the liver was attributed to the upregulation of p-AMPK expression(Figs. 8A-F). Therefore, SCP-80-I may regulate the AMPK/ACC/CPT1 signaling pathway and fasten the fatty acidβ-oxidation. The Western blotting assay yielded similar results to the RT-PCR assay.
In the pancreas, the protein expression of Sirt1 and PGC-1α in the DC group decreased significantly compared to their expression in the NC group. After 21 days of administration of SCP-80-I,the expression of Sirt1 and PGC-1α was significantly upregulated(P< 0.05). Some differences were found between the results of the Western blotting assay and those of the PCR. That might be due to the differences between transcription and translation of the same target(Figs. 8G-L). These results showed that SCP-80-I can regulate and control the transcription of the genes associated with the AMPK/Sirt1/PGC-1α signaling pathway to enhance mitochondrial function and the GSIS pathway in the pancreas.
4. Discussion
Our previous study showed that SCP-80-I could relieve hyperglycemia in T2DM rats. However, the mechanism was undetermined. In humans, the liver plays an important role in energy metabolism, and lipometabolism in the liver is regulated by fatty acid oxidation. However, morphological changes, abnormal lipid metabolism, and lipid deposition in the liver are prominent pathological features in T2DM mice. This study found that SCP-80-I can relieve liver injuries in T2DM mice and has a hypoglycemic effect. This is regulated and controlled by some related genes and signaling pathways.
Abnormal glycolipid metabolism and mitochondrial damage are the most important factors promoting T2DM[26-27]. AMPK, an indirect target of metformin, is closely associated with glycolipid metabolism and mitochondrial functions[28-29]. Abnormal lipid metabolism causes liver inflammation, and the activation of AMPK can modulate mitochondrial function in the liver[30-31].Theβ-oxidation of fatty acids occurs in the mitochondria and is regulated by AMPK. Some plant extracts can modulate histopathological changes, cell apoptosis, and mitochondrial function in the liver[32-33]. We found lipid accumulation and hepatocyte swelling in T2DM mice, and these pathological changes were minimized after treatment with SCP-80-I. Furthermore, due to liver inflammation, the transfer of long-chain acyl-CoA (LC-CoA)from the fatty acidβ-oxidation pathway to the biosynthase pathway reduces insulin sensitivity. These histopathologic changes influence glycolysis and gluconeogenesis and impair glucose levels. Regulating AMPK expression or its phosphorylationin vivocan regulate downstream targets of the two processes. The expression of theAMPKgene is below the normal level in T2DM. Polysaccharides can increase the level of AMPK and regulate the signaling pathway to ameliorate hyperglycemia[34]. A polysaccharide fromHolothuria leucospilota(HLP) could relieve liver injury in diabetic rats by targeting theAMPKgene and its related signaling pathway[35].
ACC, acetyl-CoA carboxylase, can be inhibited by AMPK phosphorylation. Polysaccharides can suppress ACC expression and its signaling pathways[36]. A sulfated polysaccharide fromEnteromorpha proliferawas beneficial against glycolipid metabolism disorder, and it downregulated the mRNA and protein expression of ACC. CPT1, carnitine palmitoyltransferase 1, is a downstream target gene of ACC and is influenced by the expression of ACC[37].Plant extracts, including polysaccharides, can relieve cellular and mitochondrial injury in the liver by restoring CPT1 activity[38-40]. A lower expression of AMPK leads to a higher expression of ACC, and that reduces the expression of CPT1 in T2DM[41-42]. This reduces fatty acidβ-oxidation in the mitochondria. Hence, the AMPK/ACC/CPT1 signaling pathway is important for diabetes patients, and it can reduce abnormal liver function caused due to diabetes by promoting fatty acidβ-oxidation.
This study found that SCP-80-I could lower TC, TG, and LDL-C levels while increasing HDL-C levels in the livers of T2DM model mice, implying that SCP-80-I could regulate hepatic lipid metabolism.Meanwhile, the content of liver glycogen was increased, which increased glucose conversion and utilization by the liver and was beneficial to blood glucose reduction. We discovered that SCP-80-I could regulate the AMPK/ACC/CPT1 signaling pathway, resulting in increased expression ofAMPKandCPT1genes and decreased expression ofACCgenes, thereby enhancing the fatty acidβ-oxidation pathway. Our results were similar to those of a study by Li et al.[43],where Hakka stir-fried tea was found to relieve obesity and inflammationin vivothrough activation of the AMPK/ACC/CPT1 signaling pathway.
The pancreas is also an important organ, and GSIS is one of the major endocrine functions of the pancreas. GSIS and basal insulin secretion work together to regulate blood glucose levels. The factors responsible for the failure of islet β-cells in T2DM include oxidative stress, endoplasmic reticulum stress, and harmful amyloid fiber deposition. Pancreatic amyloid fibers, also called amylin,accumulate on the cell surface and cause apoptosis. We found SCP-80-I could relieve oxidative stress in the pancreas in T2DM mice and reduce the deposition of pancreatic amylin in islet cells;thus, the islet β-cell apoptosis was relieved. This showed that SCP-80-I could reduce blood glucose by increasing GSIS and basal insulin secretion. Our results were similar to those of some previous studies. Polysaccharides fromAngelica sinensiscan relieve hyperglycemia by inhibiting pancreatic β-cell apoptosis[44]. Moreover,polysaccharides extracted fromG. lucidumcan also improve islet β-cell function and have a hypoglycemic effect[45].
Islet β-cells are sensitive to oxidative stress, and this process is regulated by many genes and proteins[46]. AMPK is also related to islet β-cell function, and the regulation of the expression of AMPK and its downstream genes can have a hypoglycemic effect[47-48].Sirt1,a downstream target gene of AMPK, plays an important role in many physiological pathways in mammals[49].Sirt1is a potential target for treating chronic diseases; it inhibits cancer development and protects the nervous system[50]. The activation ofSirt1can promote GSIS and inhibit key factors that cause diabetes[51-52].
UCP2, a downstream target gene regulated bySirt1, is expressed in the adipose tissue, skeletal muscle, and pancreas[53].UCP2can transport protons and increase proton conduction in the mitochondria. GSIS is triggered by glucose. Then, ATP produced in the tricarboxylic acid cycle (TCA) acts on the sulfonylurea receptor(SUR) and ATP-sensitive potassium (KATP) channels[54], resulting in plasma membrane depolarization and insulin release. Thus, ATP production is important for glycolipid metabolismin vivo. However,the activation ofUCP2decreases the ATP level produced during mitochondrial respiration. This might makeUCP2harmful to the GSIS process[55].UCP2knockout mice were better protected from an injury of the islet β-cells caused by palmitate[56]. Furthermore, those mice had a larger pancreas and more pancreatic islet β-cells during embryonic development. TheUCP2promoter can be combined withSirt1, which inhibitsUCP2transcription[51]. Thus,UCP2is associated withSirt1activation and pancreatic islet function[57-58].
PGC-1α, another downstream target gene ofSirt1, is expressed in the liver, skeletal muscle, and pancreas[59].PGC-1αcan regulate mitochondrial biogenesis and control energy metabolism[60-61].Mitochondria can affect energy supply, fatty acid metabolism, and ROS generation in islet β-cells. Hence,PGC-1αplays an important role in each stage of GSIS. Low levels ofPGC-1αin vivomay cause insulin resistance[62]. The activatedPGC-1αcan revert mitochondrial morphological changes caused by hyperglycemia and enhance insulin secretion. Based on the discussion above, we conclude that the AMPK/Sirt1/PGC-1α and AMPK/Sirt1/UCP2 signaling pathways are related to mitochondrial function and the GSIS process.
We discovered that SCP-80-I could boost the activity of antioxidant enzymes in the pancreas, reducing oxidative stress. It also improved pancreatic β-cell quality by lowering tryptic deposition and apoptosis. Furthermore, SCP-80-I improved the pathological morphology of mitochondria in T2DM model mice pancreatic cells,which had a positive effect on pancreatic mitochondrial function. And the reason for these findings could be regulated by signaling pathways such as AMPK/Sirt1/PGC-1α and AMPK/Sirt1/UCP2. After 21 days of SCP-80-I administration, we discovered that theAMPK,Sirt1, andPGC-1genes were upregulated while theUCP2gene was downregulated. Several reports have shown that polysaccharides can regulate theAMPKgene and the AMPK-related signaling pathways in vivo to ameliorate hyperglycemia. For example, a polysaccharide extracted from a traditional Chinese herb,Vaccinium bracteatumThunb.(VBTLP), could alleviate hepatic gluconeogenesis by regulating AMPK and its upstream genes[63]. AMPK participates in complex metabolic processes in cells and regulates many downstream targets in the liver and pancreas. Some of those downstream targets are related to glycolipid metabolism and insulin function. Astragalus polysaccharide (APS) can enhance glucose uptake to improve insulin sensitivity through AMPK activation[64].
We investigated three signaling pathways related to AMPK,including the AMPK/ACC/CPT1, AMPK/Sirt1/PGC-1α, and AMPK/Sirt1/UCP2 signaling pathways. Based on the results, we inferred that SCP-80-I can regulate AMPK and its downstream targets and has a hypoglycemic effect. We concluded that SCP-80-I could relieve T2DM by regulating glycolipid metabolism and islet β-cell function through the AMPK/ACC/CPT1, AMPK/Sirt1/PGC-1α, and AMPK/Sirt1/UCP2 signaling pathways (Fig. 9). We also found that SCP-80-I can enhance the expression of some genes in the liver and pancreas in T2DM to relieve the symptoms of diabetes. There might be some connection between the target gene pathways in the liver and pancreas, associated with the hypoglycemic effect of SCP-80-I.SCP-80-I can relieve abnormal liver functions caused by lipid deposition and alleviate islet dysfunction induced by lipid toxicity. In the future, we hope to uncover the mechanism of the hypoglycemic action of SCP-80-I in other metabolic organs (i.e., the kidney and intestine), as well as some other target genes and signaling pathways.Furthermore, analysis at the metabolomic level is required to complete the study of SCP-80-I in mitigating T2DM mechanisms.
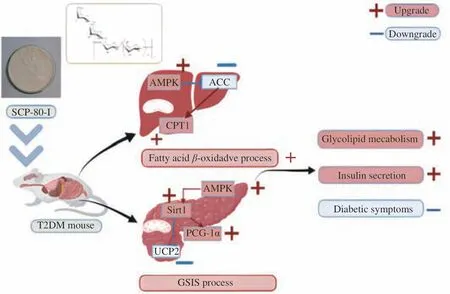
Fig. 9 Mechanism of the hypoglycemic effect of SCP-80-I on type 2 diabetic mice.
5. Conclusion
In this study, we looked at a polysaccharide (SCP-80-I) derived from sweet corn cobs that have hypoglycemic properties. SCP-80-I inhibits T2DM progression via AMPK,a key molecule in regulating bioenergetics metabolism. SCP-80-I, in particular, could increase liver glycolipid metabolism via the AMPK/ACC/CPT1 signaling pathway and increase pancreatic insulin secretion via the AMPK/Sirt1/PGC-1α and AMPK/Sirt1/UCP2 signaling pathways. These findings could provide a theoretical foundation for developing hypoglycemic functional foods based on SCP-80-I. In the future, we will investigate the mechanism of SCP-80-I to alleviate the process of T2DM from a metabolomics standpoint and provide a foundation for high-value agricultural byproduct utilization.
Conflicts of interest
All authors state that they have no personal or financial conflicts of interest.
Acknowledgment
This research was financially supported by the Doctoral Scientific Research Start-up Foundation of the Harbin University of Commerce(2019DS098), the Young Innovation Talents Project from the Harbin University of Commerce (2019CX31), and the Graduate Innovation Fund from the Harbin University of Commerce (YJSCX2019–615HSD). The authors are grateful for their support. The authors also would like to thank BioRender (biorender.com) for flowchart material and potential mechanism figure.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, in the online version, at http://doi.org/ 10.26599/FSHW.2022.9250130.

- 食品科学与人类健康(英文)的其它文章
- Betalains protect various body organs through antioxidant and anti-inf lammatory pathways
- Effects of Maillard reaction and its product AGEs on aging and age-related diseases
- Characterization of physicochemical and immunogenic properties of allergenic proteins altered by food processing: a review
- Polyphenol components in black chokeberry (Aronia melanocarpa)as clinically proven diseases control factors—an overview
- Food-derived protein hydrolysates and peptides: anxiolytic and antidepressant activities, characteristics, and mechanisms
- Recent advances in the study of epitopes, allergens and immunologic cross-reactivity of edible mango