
国际近视研究院关于轴性近视患者非病理性眼组织变化的报告
2024-01-24 02:21:44著者国际近视研究院
中华实验眼科杂志 2023年11期
著者:国际近视研究院
译者:章歆梓1 王菁菁1 何鲜桂1,2 许迅1,2
1上海市眼病防治中心 上海市眼科医院 上海市视觉健康中心,上海 200050;2上海交通大学医学院附属第一人民医院 国家眼部疾病临床医学研究中心,上海 200080
轴性近视与眼部各种非病理性和病理性组织学及肉眼变化有关[1-2]。其病因尚未完全清楚,但当眼轴过长导致图像聚焦在视网膜前面时就会发生轴性近视。在轴性近视眼中,眼部结构变化是眼轴增长的结果,主要影响位于锯齿缘后方的组织。本研究描述了与轴性近视相关的非病理性眼部组织变化,并以屈光度数为-6.0~-8.0 D和眼轴长度为26 mm作为临界值,区分轻至中度近视与高度近视之间的变化[3]。
1 眼眶和眼球形状
正视眼通常呈扁球形或球形。近视的眼轴增长导致眼球形状变为长椭圆形[4-11]。从几何角度解释这种变化的最简单方式是赤道部球壁的矢状扩张。最近研究表明,光感受器和视网膜色素上皮(retinal pigment epithelium,RPE)细胞的密度以及视网膜总厚度的减少与眼轴长度有关,尤其是在赤道与后极之间的中点,其次是赤道区域,这提示赤道后部是近视眼球壁扩张的中心[12-14]。如果眼球壁扩张的中心恰好位于赤道,则更有可能发生纯粹的眼轴增长。然而,由于眼球壁扩张的中心位于赤道后方,因此推测除了眼轴增长,眼球的水平和垂直直径也会小幅度增大,眼球壁赤道前部的区域同样会轻度扩张。在一项实验中,研究者对摘除的眼球进行了观察,证实近视眼球壁扩张的主要位置在赤道后部:在眼轴≤24 mm的眼球中,眼轴长度每增加1 mm,眼球的水平和垂直直径分别增加0.44 mm和0.51 mm;在眼轴>24 mm的眼球中,眼轴长度每增加1 mm,眼球的水平和垂直直径分别增加0.19 mm和0.21 mm[7]。这一发现也可以解释为什么眼轴增长会导致Bruch膜开口(Bruch membrane opening,BMO)在视神经乳头(optic nerve head,ONH)处扩大,因为眼轴增长引起的眼球冠状直径增加可能会增加眼后部BM的张力,BM内部张力增加可能首先导致BMO的增大,随后在黄斑区出现BM的继发性扩张缺陷[15-17]。 一些实验和临床研究结果发现,调节眼轴增长的反馈机制的感觉部分存在于眼球后部的中周边区域,这与近视眼球壁的扩张中心位于赤道后部一致[18-21]。眼球壁在赤道后部与赤道部扩张的观点同样也与临床上观察到的BMO在中心凹方向上后移相符[15-17]。BMO在中心凹方向上的移动进一步解释了轴性近视的观察结果,包括BM在视盘鼻侧边界突出至视乳头内区(例如筛板前方被视乳头周围脉络膜边缘组织包围),中度近视眼视盘形状椭圆化,以及BM在颞侧视乳头旁区(例如视乳头旁gamma区)的代偿性缺失(图1~3)[15,17,22-23]。相应地,这些具有颞侧gamma区的视盘至中心凹距离也会延长。[24]
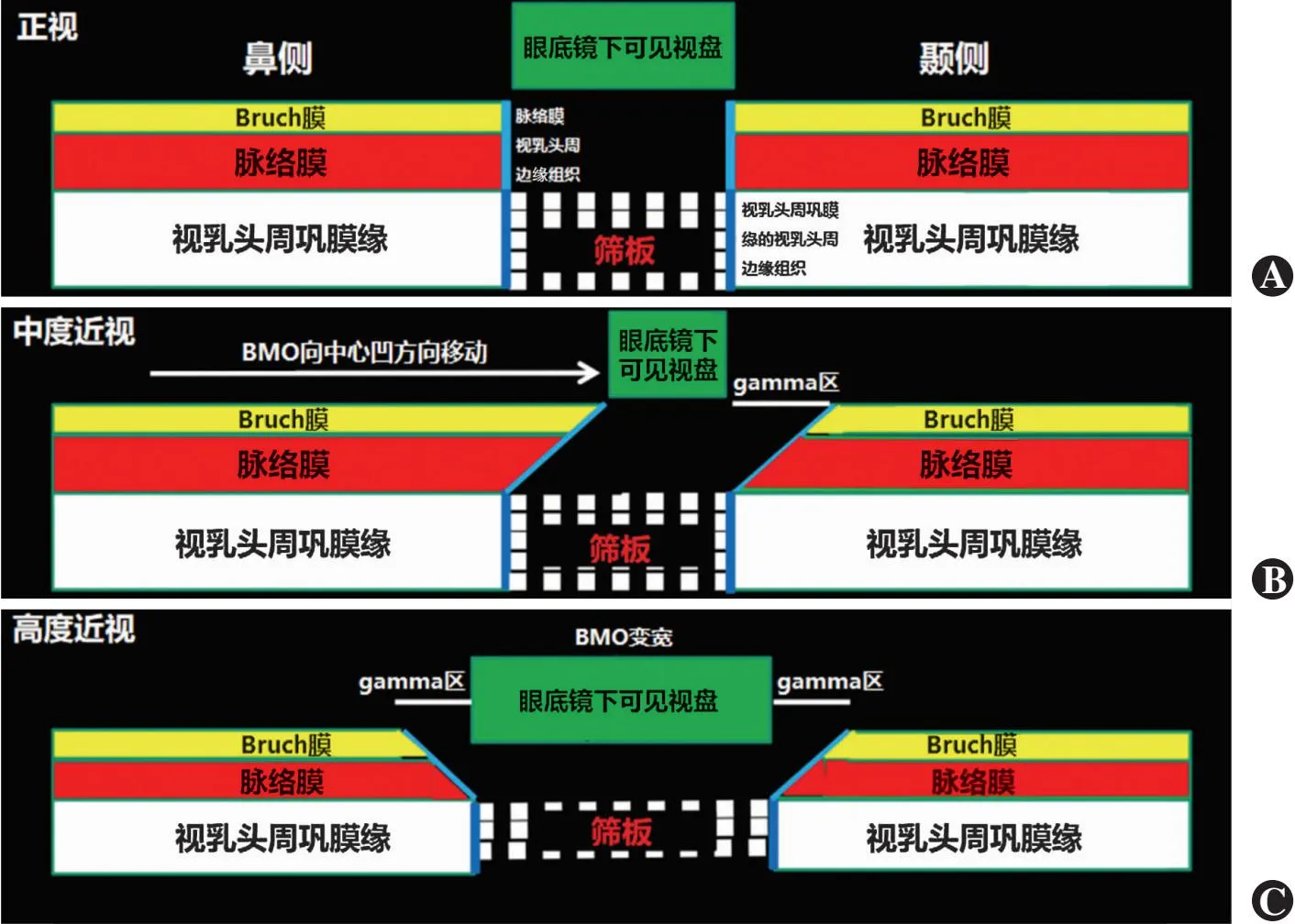
图1 ONH管壁三层结构(BMO、脉络膜开口和视乳头周巩膜缘开口)在不同程度近视眼中的排列[2] A:在正视眼中的排列 B:在中度近视眼中BMO向颞侧方向移动 C:在高度近视眼中BMO变宽
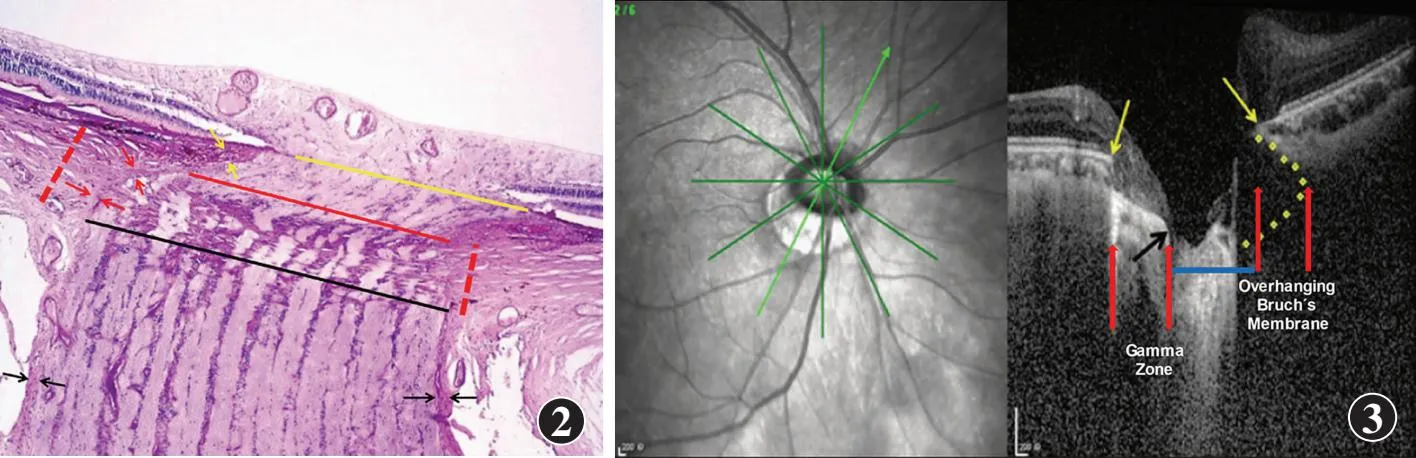
图2 中度近视眼的ONH组织学图片 ONH管的三层结构:BMO(黄线)、脉络膜开口(黄线和红线之间)、由脉络膜视乳头周边界组织(“Jacoby”)划定(黄色箭头),以及被筛板覆盖的视乳头周巩膜缘开口(红线和黑线之间)、由视乳头周巩膜缘的视乳头周边界组织(“Elschnig”)划定(红色箭头) 红色虚线:视乳头周巩膜缘;黑色箭头:视神经软膜 图3 中度近视眼ONH的光学相干断层扫描图像 BM在鼻侧突入到视乳头内,相应地在颞下方缺失(例如gamma区),以及由于突出的BM而在眼底镜下直径相对减小的视盘(蓝线) 黄色箭头:BM末端;黑色箭头:gamma区的中央端
2 ONH
ONH由ONH管和视乳头旁区组成[17]。ONH管是视网膜神经节细胞轴突(例如视网膜神经纤维)和视网膜中央静脉的出口,以及视网膜中央动脉的入口。ONH管壁由三层结构组成:内层为BMO,中层为脉络膜开口,外层是在视乳头周巩膜缘穿透开口的筛板(图1~3)[2,17]。ONH管包含约120万根神经纤维,穿过约160~300个筛板孔[25-26]。视网膜中央血管主干位于筛板的中央区域,通常轻微偏向鼻上方[27]。
3 视盘
视盘可以被视为眼底镜下神经视网膜边缘和视杯的可见部分[28]。视盘在正视眼中基本呈圆形,到中度近视眼中变为椭圆形(通常为垂直椭圆形),且通常尺寸较小[22,28-31]。BMO向颞侧的移位导致BM在鼻侧突出到视乳头内区(图1~3),这可能是视盘由圆形变为椭圆形的原因[2,16,22-23]。BM突出的部分使眼底镜下视杯鼻侧区域不可见,因此如果将视盘定义为眼底镜下神经视网膜边缘和视杯的可见部分,此时视盘就会变小并呈现竖直的椭圆形[16-17,22]。眼底镜检查时视盘呈垂直椭圆形的另一个次要原因可能是透视伪影,随着眼轴增长,眼底镜观察视盘的视角从几乎垂直到倾斜角度[32]。由于这种透视,视盘的水平直径会显得相对缩短。
尽管中度近视眼具有相对较小的视盘,高度近视眼的视盘和ONH管却往往比中度近视眼和正视眼都要大[33-34]。高度近视眼视盘的增大与筛板的延长和变薄有关[35]。筛板厚度的减小缩短了眼内腔(受眼压作用)和球后腔(受眼眶脑脊液压力作用)之间的距离[36],导致跨筛板压力梯度的增加和明显的筛板形态学变化,以及潜在的对筛板孔产生的剪切力,可能进而增加高度近视眼中类青光眼和/或青光眼样视神经病变的发病率[37-41]。
在高度近视眼中,筛板的拉伸和乳头旁gamma区、delta区发展引起的视乳头旁组织变平导致视杯变平,因为神经视网膜边缘的高度和视杯深度之间的空间差异减小[33-34]。这一因素使得神经视网膜边缘和视杯难以区分,是增加高度近视眼视神经损伤检测难度的原因之一[42]。在高度近视眼中,视盘扩大伴随着BMO的扩大,这导致鼻侧突出的BM回缩至视乳头旁区,从而形成圆形的视乳头旁gamma区[16-17,42-44]。高度近视眼的视盘形状表现出很高的个体差异性[33-34],其长轴可以是垂直的、倾斜的,有时也可以是水平的。在极度近视眼中,视盘可能受到视神经向后牵拉的影响,使眼轴明显增长的眼球内收受限[45-46]。由于视神经起源于眼眶的稍偏鼻上方,视神经硬膜在巩膜后部(例如视乳头周巩膜缘的外周端)插入引起的视神经牵拉在ONH颞下方达到最大,这将导致ONH的垂直旋转,使其在眼底镜下的形状变为垂直椭圆形。由于这种向后牵拉的最强作用点并不在ONH颞侧边界,而是位于ONH颞下方边界,因此还可导致ONH在矢状位上的旋转,使视盘上极转向中心凹方向,这可以解释ONH朝中心凹方向矢状旋转的现象。
4 视乳头旁gamma区和delta区
构成ONH的三层结构在出生时相互对齐,并与巩膜形成近乎垂直的角度(图1)。在中度近视的青少年和年轻人中,ONH管呈倾斜角度,BMO通常朝向中心凹,筛板位于其前方。这种方向导致视网膜神经纤维从后内侧向外前方斜向退出,然后在视神经内又向后弯曲,朝向眼眶的鼻上侧区域。正如本文其他部分所指出的那样,ONH管三层结构错位导致BM突出到视盘鼻侧的视乳头内区,并使BM在颞侧视乳头旁区缺失(图1~3)[16-17,23]。视乳头旁没有BM的区域即被定义为gamma区(图4)[17,43-44]。
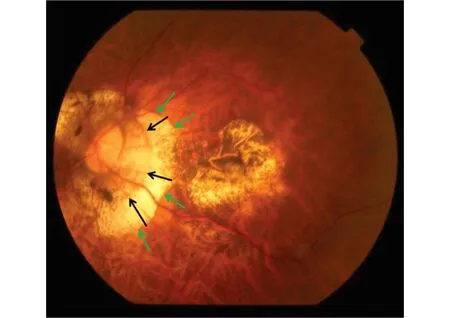
图4 伴有视乳头旁gamma区(绿色箭头)和视乳头旁delta区(黑色箭头)高度近视眼的眼底照片
导致ONH管结构错位的病因尚不清楚。Lee等[47]认为,一个潜在的机制可能是筛板向前移位,导致神经纤维从后向前斜向退出。但这种引起筛板向前移位的力量仍不明确,尤其是在眼轴增长的眼中,眼球内收时视神经是将ONH结构向后拉(如本文其他部分所述)而非向前[45-46]。另一种可能的机制是BMO向中心凹方向移位,如既往研究所讨论的[15],这种BMO向后的移位可能是由于赤道部和赤道后部的BM扩张而产生的。
除了ONH管三层结构的错位外,与眼轴增长相关的BMO扩大是可能导致gamma区扩大的第2个机制[16]。横断面研究表明,BMO扩大存在于眼轴长度大于26.0或26.5 mm(或近视屈光不正约为-8.0 D)的眼中[16]。BMO扩大导致视乳头内区突出的BM回缩,从而形成一个圆形的gamma区[44]。
视乳头旁delta区是gamma区内视乳头周巩膜缘延长和变薄的区域(图4)[17,40,43]。后部巩膜在生理上分为外部(延续成视神经硬膜)和内部(延续成视乳头周巩膜缘)[48-49],后者穿过视乳头周围边界组织与筛板相连,构成眶部脑脊液间隙的前界。视乳头旁beta区被定义为ONH周围缺乏RPE但存在BM的区域[2,17]。在具有gamma区的眼中,视乳头周巩膜缘仅被视网膜神经纤维层、脉络膜视乳头周围边界组织和内界膜覆盖[17,40,43,50]。在没有gamma区或视乳头旁beta区的眼中,视乳头周巩膜缘由脉络膜、BM、RPE以及深层和浅层视网膜层覆盖。由于视乳头周巩膜缘是筛板的生物力学锚点,高度近视相关的视乳头周巩膜缘牵拉可能对筛板的生物力学产生影响,并可能是高度近视中青光眼或类青光眼样视神经损伤增加的原因之一。有研究表明,继发性大视盘和大delta区在临床上与更高的青光眼或类青光眼样视神经损伤患病率相关[17,37,40,51-52]。
与眼轴增长相关的BMO扩大和gamma区、delta区扩大造成了视野中盲点的扩大,由此产生的暗点是绝对性的,因为这些区域缺乏视网膜光感受器。
5 Zinn-Haller动脉环
Zinn-Haller动脉环位于视神经硬膜与后部巩膜的交界处,视乳头周巩膜缘的外周端[53],为包括筛板在内的ONH提供营养。如果在眼底镜下可见,动脉环代表着delta区与gamma区的边界[17]。Delta区的延长会导致动脉环与筛板之间距离的增加,这种血供与筛板之间距离的增加可能是高度近视患者易发生青光眼或类青光眼样视神经损伤的另一个原因。
6 视乳头周围边界组织
在ONH管中,视乳头周脉络膜和视乳头周巩膜缘由两者的视乳头周边界组织和视乳头内区分开(图2)[50,54-55]。视乳头周巩膜缘的边界组织是视神经软膜的延续,并继续进入脉络膜边界组织,与BM的末端相连[17,50],由于和视乳头周巩膜缘的胶原纤维垂直交叉,并通过脉络膜边界组织与BM相连,故视乳头周巩膜缘边界组织在矢状位上为筛板提供了生物力学稳定性。脉络膜边界组织分隔了脉络膜内腔和视乳头内区,前者由于脉络膜毛细血管的腔隙,荧光素和白蛋白会渗漏到细胞外间隙中,而后者的血管没有腔隙,因此不会发生此类生理性渗漏。因此,边界组织可能位于假定的脉络膜-ONH屏障的位置。脉络膜边界组织连接着筛板的浅层外周端和BM末端,在具有gamma区和delta区的眼中,其延长量和BM末端在视盘边缘的回退量基本相同。脉络膜边界组织的延长伴随着相应的变薄,因此在gamma区扩大的眼中,脉络膜边界组织的体积并不受影响[50],但因为变薄有可能发生破裂,使BM末端不再与ONH牢固连接。这可能是具有大gamma区的近视眼中BM末端卷曲的原因,并可能导致BM的继发性皱褶,后者可在组织学和光学相干断层扫描(optical coherence tomography,OCT)图像上观察到[56]。
除了眼前节的巩膜小突和眼后节的涡静脉,脉络膜边界组织是连接眼球内部(如脉络膜复合体、BM、RPE、视网膜、睫状体、虹膜、晶状体和玻璃体)与外部(如巩膜和角膜)的唯一结构。由于巩膜直接与眼外肌相连,会经历显著的旋转加速和减速运动,而内部的球体遵循惯性定律,在加速时会滞后,在减速时会追赶,这种外部与内部球体之间运动的不一致性会对连接两者的巩膜小突和脉络膜视乳头周边界组织产生压力和牵拉,这一发现意味着脉络膜视乳头周边界组织在眼的生物力学中具有重要意义。然而目前还不清楚与眼轴增长相关的脉络膜视乳头周边界组织的伸长和变薄对ONH和黄斑可能产生哪些生物力学影响。
7 视神经
高度近视已被确认为与青光眼样神经病变以及非青光眼样视神经损伤相关的主要因素[1,37,40-41,57-59]。这种青光眼样视神经病变的发病率增加可能是由于筛板的延长和变薄,伴随着相关的筛板内组织变化和跨筛板压力梯度的增大、作为筛板生物力学锚点的视乳头周巩膜缘的延长和变薄,以及视乳头周围Zinn-Haller动脉环与筛板之间距离的增加。而高度近视眼中非青光眼样视神经损伤的发病率增加可能是由于视网膜神经节细胞体到视盘的距离增加,导致视网膜神经纤维延长和潜在牵拉[17,59]。视网膜神经节细胞体与视盘之间距离的增加是由于眼轴增长和gamma区扩大造成的视网膜延长,进而导致中心凹与视盘之间的距离增加[24,60]。目前尚不清楚中度近视是否是青光眼样视神经病变的危险因素[61-64]。中度近视也尚未被描述为非青光眼样视神经损伤的危险因素。
8 视网膜
眼轴增长导致眼球周长增加。根据最近一项针对摘除人眼进行的组织形态学研究,眼球周长的增加小部分与睫状体长度(包括睫状冠和平坦部)增加有关,而大部分与视网膜延长有关,例如从锯齿缘到视盘和黄斑的距离增加[60]。眼轴长度每增加1 mm,视网膜长度(从锯齿缘到视盘)增加0.73 mm(95%CI:0.65~0.81),睫状体长度(从巩膜小突到锯齿缘)增加0.16 mm(95%CI:0.12~0.20)[60]。视网膜长度和睫状体长度彼此相关,视网膜长度每增加1 mm,睫状体长度就增加0.12 mm(95%CI:0.07~0.17)。这些发现与先前观察到的研究结果一致,即近视眼球扩大主要影响轴向眼球直径,其赤道水平和垂直直径的增加程度明显较低[4-11]。
组织形态学结果显示,与眼轴长度相关的光感受器密度下降在赤道后部最为明显,赤道部次之,最后是锯齿缘[12]。正如本文其他部分所讨论的,这些发现表明眼轴增长主要发生在赤道后部,赤道部次之,最后是锯齿缘。相应地,赤道部和赤道后部的视网膜总厚度随着眼轴长度的增加而变薄,而黄斑区的视网膜厚度与眼轴长度无关或关联不明显[13]。既往临床研究中,在纠正了年龄、性别、视盘-中心凹距离和中心凹下脉络膜厚度等参数后,较薄的黄斑区视网膜外核层(作为光感受器密度较低的替代指标)被证明与更长的眼轴相关[12,70]。一些研究应用了自适应光学扫描激光眼底成像,结果显示中心凹视锥细胞的马赛克排列随着眼轴增长而扩张[71-75]。然而,视锥细胞的角度采样密度(以每平方度中的视锥细胞数量表示)究竟是否随眼轴长度变化而变化尚不清楚[71-75]。一般而言,我们可以假设近视者的光感受器和RPE细胞数量与非近视者相同,但在近视眼中,由于光感受器分布在与眼轴增长相关的更大的内表面上,因此它们的整体平均密度随着眼轴增长而降低。然而在中心凹区,光感受器的密度并没有随眼轴增长而明显减少,这一发现与其他中心凹的形态结构(例如BM厚度和中心凹RPE细胞密度)和眼轴长度关联不明显或根本不相关一致。
从临床角度来看,近视相关的赤道后部和赤道部光感受器密度下降可能直接导致空间分辨率的下降和感受野的扩大,后者也许与内核层和视网膜神经元层细胞密度的潜在下降相关。这些假设与临床观察到的高度近视眼的视野向心性收缩相吻合[76]。在这种情况下,还需要考虑到视轴的增长可能导致视网膜上的投影图像变大。Chui等[77]的临床研究结果显示,周边部视网膜位置的视觉分辨率与近视屈光不正大小呈线性下降。与正视眼相比,屈光度为-15.0 D眼的视网膜感受单元之间的间距是正视眼的2倍,即只具有50%的外周视觉分辨率[77]。Chui等[77]利用光栅视力数据对眼球扩张模式进行分析,发现近视眼球壁扩张的中心位于眼球体几何中心的后方。这一发现与本文其他部分所讨论的关于眼球中周部RPE细胞和光感受器的密度以及视网膜厚度的组织形态学数据相一致。
其他的与眼轴增长相关的视网膜非病理性变化包括赤道部和赤道前部的格子样变性和鹅卵石变性[78-81]。从正视到中度近视,格子样变性的患病率逐渐增加,在眼轴长度为26.0~26.9 mm(或屈光度为-6.00~-8.70 D)时达到峰值,然后随高度近视和极度近视下降。[79]鹅卵石变性的患病率通常随眼轴长度增加而增加,在高度近视眼中最高。鹅卵石变性区域由于RPE层黑色素的稀少或完全缺失而呈明亮的白色。在最近一项组织形态学研究中,鹅卵石变性区域的BM和脉络膜毛细血管层厚度较薄,而其外侧厚度较无鹅卵石变性眼的对应区域更厚[81]。鹅卵石变性区域表现出组织紊乱的视网膜与变薄的BM之间的牢固粘连,且只有少数RPE岛存在,巩膜和总脉络膜则没有明显的区域性变薄。在光学显微镜下,鹅卵石变性区域内的BM(除RPE岛外)似乎是单层的,而在鹅卵石变性区域外是双层的。恶性脉络膜黑色素瘤已被发现在空间上与肿瘤前方的鹅卵石变性相关[82],该研究提出,由于脉络膜恶性黑色素瘤以及其他肿瘤(如脉络膜痣和脉络膜转移瘤)引起的周边脉络膜血流灌注不足可能是导致鹅卵石变性的原因。
9 RPE
与光感受器相似,位于其下方的RPE在眼轴长度增加时也显示出细胞密度的降低,其中以赤道后部最为显著[14]。有趣的是,RPE细胞密度在后极部并不降低或只是轻微降低[14]。这种RPE细胞密度与眼轴长度之间的逆相关性在赤道后部最强,赤道部次之,最后是锯齿缘,而在后极部最弱,这与近视眼球壁扩张主要发生在赤道后部一致,但后极部本身,除了gamma区和delta区的形成外,似乎并未主要参与该过程[15]。尽管并不总是能观察到,但眼轴长度增加时后极部RPE细胞密度轻微降低,这和眼轴长度增加时中心凹视锥细胞密度降低一致[71-75]。因此光感受器密度/RPE细胞密度的比值可能不会受到眼轴长度的明显影响。
10 BM
针对欧洲和中国人眼以及豚鼠模型进行的组织形态学研究表明,BM厚度与眼轴长度无关[83-85]。即使在眼轴长度超过30 mm的极端情况下,中心凹BM的厚度也与较短眼轴长度眼相似。这些发现提示,BM的表面积随着眼轴长度的增加而增加,但其厚度保持不变,因此BM的体积会随着眼轴增长而增加,这与巩膜和脉络膜的体积不随眼轴伸长而增加相反[86-87]。BM能够保持其厚度的能力表明BM在眼轴增长的过程中起到了积极的作用。
需要注意的是,BM的厚度仅为2~4 mm,光学显微镜很难检测到BM厚度较小的变化。随着拉伸应力的增加,物体会发生延长,其比例由杨氏模量描述。假设BM在眼轴增长眼中并不发生重塑,那么它应该处于某种拉伸应力之下,而如泊松比所描述的,随着这种应力的增加,通常会出现部分组织变薄。这些情况使人怀疑BM可能确实有变薄,但在以往的光学显微镜检查中并未被发现[84-86]。
从中央小凹到视盘边缘的距离随着眼轴长度的增加而增加,而从中心凹到gamma区边界的距离与眼轴长度无关[24]。研究表明,与眼轴长度相关的视盘至中心凹距离增加是由于gamma区的形成和扩大,而黄斑区的BM长度则与眼轴增长无关。相应地,在没有黄斑区BM缺陷眼中,颞上动脉弓和颞下动脉弓之间的距离与眼轴长度无关,这与在无BM缺陷的眼轴增长眼中黄斑区BM不会扩大的观点相吻合[88]。
一些有明显眼轴增长眼的眼后部表现出BM缺损,并在空间上与巩膜葡萄肿相关联[89]。BM缺损被一个没有RPE细胞的较大区域包围,在缺损边缘BM的厚度并未变薄。目前还不清楚在眼轴增长眼中,特别是在高度近视眼中,BM是否同弹性假黄瘤和AMD一样表现出生化成分的变化(例如钙化程度)[90-94]。这个问题可能也与漆裂纹的形成有关,眼底镜下的漆裂纹或许即RPE层和(部分眼中)其下方的BM的线性缺损[95-98]。RPE细胞缺失造成的黄斑区RPE层和BM缺损与视野中的绝对暗点相一致。
11 中心凹-视盘距离
由于颞侧gamma区的形成和扩大,中心凹-视盘距离随着眼轴增长而增加[24]。颞上和颞下血管弓之间的距离与眼轴长度无关,因此颞侧血管弓之间的夹角随着眼轴增长而减小[88]。这个过程导致了ONH颞侧视网膜神经纤维层的拥挤以及其他区域相应变薄,特别是视网膜神经纤维层厚度曲线峰值的位置向颞侧移动[88,99-100]。这些视网膜神经纤维层厚度的变化与眼轴增长时kappa角(例如以视盘为顶点的颞上血管弓和颞下血管弓之间的夹角)的减小有关,在根据视网膜神经纤维层厚度诊断视神经图像时必须考虑到这些变化。中心凹-视盘距离的增加导致了乳头黄斑束中视网膜神经纤维的延长和潜在拉伸[101],这可能导致非青光眼样视神经损伤,并可能解释了在高度近视眼中发生的、无法用黄斑形态去解释其病因的中央旁暗点现象。视网膜神经纤维从视网膜神经节细胞体到视盘的走行原本主要呈弧形,在这种情况下,它们可能通过采取更直的路线来补偿与视盘距离的增加。相应地,在进展的高度近视眼中,视网膜血管向视盘的路线也会更直[101]。
12 脉络膜
正如临床研究所示,脉络膜厚度随着眼轴增长而变薄,以中心凹下区域最为明显[102-104]。脉络膜厚度变薄已被证明主要影响中、大脉络膜血管层,而脉络膜毛细血管层的厚度则与眼轴长度的增加无关,或仅有少量关系[101]。相应地,随着眼轴增长,大、中脉络膜血管层在脉络膜总厚度中的比例下降,毛细血管层的比例则增加[101]。
在最近一项针对非高度近视眼的临床研究中,利用OCT血管成像,在校正了年龄、眼压、血清高密度脂蛋白胆固醇浓度和图像质量评分等因素后,多因素分析结果显示脉络膜毛细血管血流缺损与眼轴长度无关[105]。其他研究也报告了类似的结果,但并未纠正与眼轴长度相关的图像放大效应[106-107]。在Jiang等[108]进行的一项研究中,脉络膜毛细血管层的整体血管密度在高度近视和非高度近视眼之间没有差异。相反地,一项研究报告称,通过OCT血管成像测量,随着眼轴增长和脉络膜变薄,脉络膜的血管密度和毛细血管层的血流灌注减少[109]。类似地在另一项应用SS-OCT血管成像的研究中,高度近视眼中较长的眼轴与中心凹周边区域更高的脉络膜毛细血管血流缺损相关,尽管在中心凹旁和中心凹区没有发现这样的关联[110]。
约17%的高度近视眼有脉络膜内视乳头旁腔(也称为脉络膜上视乳头旁腔),在最初被描述为视乳头周围脱离[68]。脉络膜内视乳头旁腔定义为位于视乳头旁区的脉络膜(与BM相连)和巩膜之间的裂隙(图5)[65-69]。此腔隙通常出现在视乳头旁区的下方至颞下方。如Spaide等[67]所描述,这些病变在眼底镜下呈现为ONH边界下方至颞下方的橙色区域,伴有巩膜向后弯曲,但其上方的BM-RPE-视网膜复合体没有发生变形。巩膜的后外弯曲可能提示腔隙形成的原因是巩膜的后方移位,而非视网膜和RPE的前移[67]。根据生理上的弹性应力-应变关系,组织抵抗形变的能力与其厚度和组成有关,与眼压相关的后巩膜壁形变在视乳头旁gamma区和delta区会比眼底其他区域更为明显,因为gamma区和delta区上方的组织要么不存在,要么异常薄[67]。如本文其他部分所述,解释脉络膜腔隙发生的另一种可能是,视神经相对较短,不足以使眼轴明显增长的眼球进行充分的内收[45-46,69]。在极度增长眼球的内收过程中,视神经(可能是视神经硬膜)在硬膜与巩膜融合线上对后巩膜(例如视乳头周巩膜缘的外周端)施加拉力,这可能导致对视乳头周巩膜的向后牵拉,以及巩膜和依附在BM上的脉络膜之间的继发性裂隙。脉络膜上视乳头旁腔不被认为是病理性的,因为它通常不伴随功能缺陷,并且尚未被报告为近视相关病变的危险因素[68]。关于近视防控治疗对脉络膜影响的详细内容,请参阅IMI-动态的脉络膜白皮书[111]。
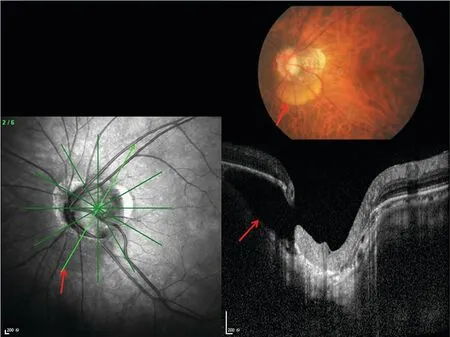
图5 视乳头旁脉络膜内腔的眼底照片和OCT图像
13 巩膜
在眼轴增长的过程中,巩膜经历了明显的重塑,其厚度随着眼轴长度增加而减少,在后极部最为显著,在锯齿缘和锯齿缘前部变化最小[4,112-114]。组织形态学研究表明,在3岁及以上的人群中,巩膜的横截面积和体积与眼轴长度和年龄大小均无关,而在2岁及以下的儿童中,巩膜的横截面积和体积随年龄增长而增加[86-87]。
巩膜重塑伴随着细胞外基质 (extracelluar matrix,ECM) 成分和巩膜成纤维细胞的变化或者说由这些变化诱导[115-135]。巩膜由致密的纤维状粘弹性结缔组织构成,其中包括不规则排列的胶原纤维以及分布在纤维之间的蛋白多糖和非胶原糖蛋白。巩膜成纤维细胞产生胶原蛋白和ECM的其他成分。研究表明,眼轴增长可能与活跃的巩膜重塑相关,造成巩膜基质的弱化[118]。作为早期巩膜重塑的证据,在树鼩眼实验性近视和恢复过程中观察到了巩膜代谢和弹性的增加[115-116,121]。从生物力学的角度来看,树鼩的研究揭示,组织延展与时间的比率 (例如巩膜蠕变率) 在近视进展的眼中增加,而在近视改善时则减少[121],这些生物力学变化过去被认为与巩膜肌成纤维细胞诱导的巩膜 ECM 变化有关。Curtin 等[135]通过电子显微镜进一步证实了近视眼巩膜形态构成的变化:与正视眼相比,近视眼的巩膜胶原纤维束主要呈层状排列,其纤维直径减小而纤维直径的范围较大,异常的星状纤维数量增多,以及直径均匀且非常细小的纤维群数量增加[135]。
组织应力和组织生长因子(包括TGF-β)会影响肌成纤维细胞。肌成纤维细胞是表达α-平滑肌肌动蛋白的分化后成纤维细胞。实验性近视树鼩眼巩膜中组织金属蛋白酶-2抑制剂的表达减少,当外源性升高组织金属蛋白酶-2抑制剂时,近视发展和眼轴增长显著减慢,后巩膜胶原降解也受到抑制[125,127]。
在一项针对高度近视患者的全基因组关联研究中,发现了4条京都基因与基因组百科全书信号通路,包括苯丙胺成瘾、ECM受体相互作用、神经活性配体受体相互作用和肌动蛋白细胞骨架调控通路,与高度近视相关,缺氧诱导因子-1α信号通路与超过-10.0 D的高度近视相关[132]。这些结果与观察到的缺氧信号、eIF2信号和哺乳动物雷帕霉素靶向信号通路在小鼠近视模型中的激活一致[133]。此外,这些结果也支持了既往其他研究中,小鼠和豚鼠的实验性近视与巩膜中缺氧诱导因子-1α上调相关的结论。类似地,人类巩膜成纤维细胞暴露于缺氧环境会诱导肌成纤维细胞转分化,Ⅰ型胶原表达下调。在另一项针对高度近视患者的临床研究中,巩膜相关基因多态性的检测结果显示TGF-β2的多态性与近视相关,因此TGF-β2可能是巩膜重塑过程中的一个重要因素[134]。
目前尚不清楚究竟是眼轴增长相关的巩膜变化导致眼轴增长,还是眼内组织的延长在前而巩膜改变在后。有假设认为参与近视巩膜重塑的信号通路包括多巴胺、视黄酸和腺苷。具体请参阅Summers等(2021)[136]和Brown等(2022)[137]的综述。
14 玻璃体
随着眼轴增长,玻璃体的大分子发生改变,玻璃体黏度降低,玻璃体后脱离的发生率增加[138-142]。虽然大分子仅占玻璃体总量的2%,但其对于玻璃体的透明度、胶体状态和生理功能非常重要[142]。早期对人眼的研究报告指出,近视眼玻璃体的蛋白质浓度、胶原含量和透明质酸浓度较正视眼降低[143]。在最新的小鼠近视模型研究中发现,近视眼玻璃体中的钾、钠和氯含量降低[144]。随着眼轴增长和年龄增长,玻璃体内胶原纤维变性,纤维-透明质酸结合的胶体液化,玻璃体发生混浊,并最终造成玻璃体后方与视网膜和ONH内界膜的脱离。在某些情况下,玻璃体后脱离后在ONH处发生凝结和粘连,形成Martegiani环,被视为较大的漂浮物,如果临床上很明显,就会引起视觉质量的降低,这种情况被称作“视觉退行性飞蚊症”[142]。在一些高度近视眼中,可能发生玻璃体不完全后脱离,玻璃体皮质残留在黄斑区[139]。
15 眼前节
与眼后节相比,近视眼轴增长所引起的眼前节变化较小,角膜厚度和直径与眼轴长度基本无关[145-147]。角膜前表面曲率半径在中度近视眼中随眼轴增长略微减小,但在高度近视眼中与眼轴长度无关。前房深度和前房角随眼轴增长增加。相应地,较长的眼轴与更低的原发性开角型青光眼患病率、发病率相关。中度近视眼中,晶状体厚度随眼轴长度增加而减小,而在高度近视眼中晶状体厚度与眼轴长度无关。
16 结论
近视眼轴增长相关的非病理性变化主要发生在眼球的后半部分。随着眼球形状从球体变为长椭圆体,光感受器和RPE细胞密度以及视网膜总厚度下降,其中以赤道后部最显著,其次为赤道部,而在后极部最不明显。脉络膜和巩膜在后极变薄最明显,在锯齿缘最不明显。此外,在近视发生早期巩膜即发生重塑,脉络膜和巩膜厚度发生变化的同时体积并未改变;BM的厚度与眼轴长度无关,但其体积随眼轴增长而增加。在中度近视中,BMO通常朝中心凹的方向移动,导致BM向鼻侧视乳头内区突出,颞侧BM缺失(例如视乳头旁gamma区),眼底镜下可见视盘水平直径缩短呈椭圆形,进而致检眼镜下观察视盘面积减小,中心凹-视盘距离增大(由于gamma区形成,但黄斑部BM不延长),kappa角减小,乳头黄斑束视网膜血管和视网膜神经纤维变直和拉伸。高度近视眼的解剖特征包括BMO增大,视盘增大,筛板、视乳头周巩膜缘(例如视乳头旁delta区)和视乳头周脉络膜边界组织延长和变薄,以及圆形gamma区和delta区的形成。全面了解非病理性近视的眼部变化特征对于更好地理解近视眼轴增长的机制、病理性结构变化以及近视导致的心理生理后遗症对视功能的影响具有重要意义。
利益冲突国际近视研究院报告的出版费用由Brien Holden Vision Institute、Carl Zeiss Vision、CooperVision、EssilorLuxottica、Hoya、Thea、Alcon和Oculus的捐赠支持;本文所有作者均声明不存在利益冲突
志谢感谢国际近视研究院的支持和帮助。感谢Rebecca Weng对本文翻译进行校对
声明本文著者并未参与本次翻译过程
参考文献(略)
猜你喜欢
军事文摘(2024年2期)2024-01-10 01:58:44
国际眼科杂志(2023年3期)2023-04-15 15:59:29
中国科技纵横(2021年7期)2021-07-22 02:55:56
临床眼科杂志(2021年2期)2021-05-26 03:26:38
青少年科技博览(中学版)(2020年1期)2020-04-21 08:57:40
奥秘(创新大赛)(2019年4期)2019-04-15 07:13:08
中医眼耳鼻喉杂志(2018年1期)2018-04-10 02:54:56
奥秘(2015年3期)2015-09-10 07:22:44
纯碱工业(2014年6期)2014-03-11 15:09:25
山西大同大学学报(自然科学版)(2014年5期)2014-01-23 01:58:01
