
油茶根腐病与丛枝菌根真菌群落的相关性分析
2024-01-22 05:05杨娅琳吴峰婧琳陈健鑫武自强张东华马焕成伍建榕
西南农业学报 2023年11期
杨娅琳,吴峰婧琳,陈健鑫,武自强,刘 丽,张东华,马焕成,伍建榕,
(1.西南林业大学生物多样性保护学院/云南省高校森林灾害预警控制重点实验室,昆明 650224;2.西南林业大学林学院/西南地区生物多样性保育国家林业局重点实验室,昆明 650224)
【研究意义】油茶(CamelliaoleiferaAbel.)又称茶子树,属于山茶科山茶属,是中国南方特有的木本油料树种[1],它与油棕、油橄榄和椰子一起被称为世界四大木本油料植物。油茶具有环境适应性强、耐贫瘠、果期长、树龄长等优点,因而成为中国南方推广种植的经济林木。油茶主要分布在中国南方的大部分省份(如江西、湖南、云南等),种植历史长达千年,云南具备高原特色油茶产业得天独厚的自然条件,以野生型油茶为主[2]。油茶压榨出的茶油是一种优质的植物油,不饱和脂肪酸含量高达90%以上,还富含油酸、亚油酸、硬脂酸和茶多酚等活性成分,具有抗氧化、抗炎和抗菌等功效[3]。国家林业和草原局公布了《林草产业发展规划(2021—2025年)》,根据规划,油茶种植面积将达6.00×106hm2,油茶年产量将达2.00×106t[4]。丛枝菌根真菌(Arbuscular mycorrhizal fungi, AMF)是林地土壤中最普遍、最重要的真菌类群,定殖于植物根系后能与植物建立起良好的共生关系,研究发现,AMF能够与80%以上的陆生植物形成共生关系,形成互惠共生体[5-7]。植物分泌信号诱导AMF孢子的萌发并侵入根系,在根系皮层内形成特殊结构,如丛枝(Arbuscule)、泡囊(Vesicle)和胞间菌丝等[8]。【前人研究进展】油茶是能与AMF共生的植物之一,林宇岚等[1]通过高通量测序技术发现不同品种油茶根际土壤共有AMF 12个属,说明油茶根际土壤内有丰富的AMF种类。AMF能够提高植物的抗旱和抗盐碱能力[9-10],缓解重金属对宿主植物的影响[11],提高宿主植物对病虫害的抵御能力[12],促进植物对矿质营养元素的吸收[13],增强宿主植物抗污染物毒害的能力[14],还能够调节植物的种间关系[15]。Safir[16]研究发现摩西球囊霉(Glomusmosseae)定殖能够减少洋葱(Alliumcepa)根部病原菌红根腐菌(Pyrenochaetaterestris)的侵染,提高洋葱的抗病性。周昱[17]研究发现,接种丛枝菌根真菌可提升对云杉根腐病的防治效果。如今,已有越来越多的研究关注AMF对植物病原菌的防控作用。【本研究切入点】油茶根腐病在苗期是一种毁灭性的土传疾病。油茶患根腐病后地上部分表现为植株发育不良、矮小,叶片黄化脱落,甚至整株干枯死亡;受影响植株地下部分根部在初期变成褐色至深黑色,后期根部表皮腐烂脱落,延伸至地面根茎交界处,影响水分和养分运输,植株的生长发育受阻,严重影响油茶成林及后续产量[5]。【拟解决的关键问题】德宏州油茶集约化种植以及管理方式等原因导致油茶根腐病大面积发生,油茶品质下降。本研究以德宏州梁河县健康植株和根腐病发病植株根系和根际土壤为研究材料,探究根腐病的发生与AMF群落的关系,为充分挖掘和利用AMF生防、促生功能提供依据。
1 材料与方法
1.1 材料采集
样品于2021年8月27日采自德宏州梁河县九保乡丙界村(98°19′40″ E,24°48′59″ N,海拔1451 m)。德宏州梁河县是一个典型的农业县,境内森林覆盖率60%,具有丰富的森林资源,属南亚热带季风气候,四季不分明,雨量充沛,立体气候明显,日照时数长。
研究对象为10年生的白花油茶,品种为德林油B1。根据油茶地上部分黄化、叶片脱落等症状选择对象,共选取15棵目标树。在距离主干约30~50 cm的东南西北4个方向须根丰富处,采集油茶根系,轻晃掉附着在根系表面不紧密的土壤,收集剩余附着在根系的土壤,充分混匀,为根际土壤样品(S);抖落土壤后的二级侧根,为根系样品(R)。将样本用干冰保存运回实验室,放入-80 ℃的冰箱备用。根系分为2份,1份用于分子实验,1份用于AMF侵染率测定;土壤以200目筛网过筛,分为2份,1份用于分子实验;1份用于理化参数测定。
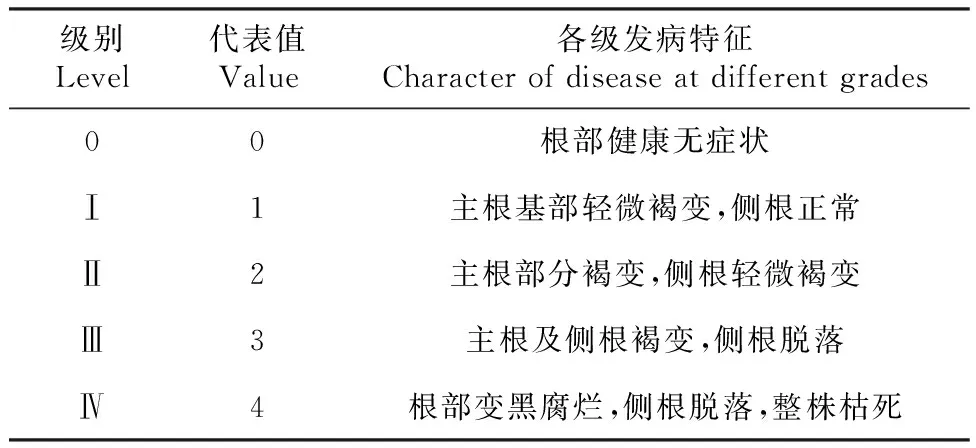
表1 病害严重度分级标准
病情指数参考方中达[18]制定的分级标准(表1)计算。每个病害等级下采集3份不同油茶植株的根系及根际土壤,充分混合为1个样本,最后得到5个根系样本和5个根际土壤样本,0、Ⅰ、Ⅱ、Ⅲ、Ⅳ级下根系分别命名为HR0、DR1、DR2、DR3、DR4,根际土壤分别命名为HS0、DS1、DS2、DS3、DS4。对采集的根系进行表面消毒,用75%乙醇浸泡消毒1 min,再用无菌水冲洗1次,然后用5%次氯酸钠溶液浸泡消毒5 min,最后用无菌水冲洗3次,即完成表面消毒[19]。
1.2 油茶根系中AMF的定殖情况观察及根际土壤中AMF孢子密度检测
计算AMF的孢子密度(Spore density),采用湿筛沉淀法分离根际土壤中的AMF孢子,将分离得到的孢子置于立体显微镜下观察并计数,以每20 g风干土样中含有的孢子数表示。
计算AMF定殖率,采用AMF定殖染色法:先将根样用无菌水冲洗4~5次;再用90 ℃的10% KOH水浴解离2 h;待自然冷却后,蒸馏水冲洗4~5次;再放入2% HCl溶液内常温浸泡5~10 min;用蒸馏水将根系清洗干净,加入0.05%苯胺蓝溶液室温染色1 h;随后用脱色液[m(甘油)∶m(乳酸)∶m(蒸馏水)=1∶1∶1]脱色2~3 d后制成玻片。在光学显微镜下观察,每个重复至少检测30个根段观察菌根定殖情况,记录其定殖率。
菌根真菌定殖率(%)=∑(0×根段数+10%×根段数+20%×根段数+……+100%×根段数)/观察总根段数
1.3 高通量基因组测序方法
根系表面经过洗净消毒后,采用液氮研磨的方法将样品磨成粉末状以便提取,采用E.Z.N.A Mag-Bind soil DNA Kit提取试剂盒(OMEGA)进行土壤和植物根系DNA提取,2%琼脂糖凝胶检测DNA完整性,DNA样本浓度采用Qubit3.0荧光定量仪检测。采用AMV4.5NF(5’-AAGCTCGTAGTTGAATTCG-3’)/AMDGR(5’-CCCAACTATCCCTATTAATCAT-3’)引物对AMF真菌rDNA进行PCR扩增,采用30 μL扩增体系,PCR反应条件:94 ℃,3 min→(94 ℃,30 s→45 ℃,20 s→65 ℃,30 s)5次循环→(94 ℃,20 s→55 ℃,20 s→72 ℃,30 s)20次循环→72 ℃,5 min→10 ℃,∞。第二轮扩增引入Illumina桥式PCR兼容引物,采用30 μL扩增体系,PCR反应条件:95 ℃,3 min→(94 ℃,20 s→55 ℃,20 s→72 ℃,30 s)5次循环→72 ℃,5 min→10 ℃,∞。利用Illumina 高通量测序仪测序平台进行测序。PCR扩增及测序工作由生物工程(上海)股份有限公司进行。
1.4 土壤养分检测
1.5 生物学信息分析
将二代测序得到的PE读数首先按照Overlap关系进行拼接,区分样本后进行序列质量质控和筛选,然后进行OTU聚类(ASV去噪)分析和物种分类学分析。所有序列都可以根据不同的相似性等级划分OTUs,通常对97%相似等级的OTUs进行生物信息统计分析。根据不同的扩增类型,通过相应的数据库对OTU代表序列进行分类和分析。最后,在每个分类级别上统计每个样本的群落组成:界(Kingdom),门(Phylum),纲(Class),目(Order),科(Family),属(Genus),种(Species)。
1.6 统计学数据分析
根据PE读数之间的Overlap关系,PEAR用于将成对的读数合并为一个序列;使用PRINSEQ在读取尾部切割质量值小于20的碱基,并设置10 bp窗口,如果窗口中的平均质量值低于20,则从窗口中切掉后端碱基,筛选出质量控制后的含N序列和短序列,最后筛掉低复杂度序列。选取具有97% 相似性的OTU分类等级,利用 Mothur 计算不同随机抽样下的α多样性指数,利用Excel 2003软件统计各分类单元的数据;Qiime(v1.9.1) 用于计算β多样性距离矩阵;用R 语言工具绘制香农指数、维恩图和PCA分析的稀释曲线。直方图绘图由GraphPad绘制。

表2 不同病情等级根腐病油茶土壤理化性质测定
2 结果与分析
2.1 土壤理化性质
由表2可知,随病害等级增加,土壤pH呈升高趋势,且发病等级Ⅲ、Ⅳ级与0、Ⅰ、Ⅱ级油茶土壤间的pH存在显著差异;发病等级为0级和Ⅱ、Ⅲ、Ⅳ级的油茶土壤有机质存在显著差异,且病害等级随有机质含量的增加而升高;发病等级为Ⅲ级和0、Ⅰ、Ⅱ、Ⅳ级的油茶土壤速效磷有显著差异;发病等级为Ⅳ级和0、Ⅰ、Ⅱ、Ⅲ级的油茶土壤速效钾有显著差异;油茶根腐病病害等级随全氮含量的增加而升高;发病等级为Ⅲ、Ⅳ和0、Ⅰ、Ⅱ级的油茶土壤全钾存在显著差异,且病害等级随全钾含量的增加而降低;发病等级0、Ⅰ与Ⅱ、Ⅲ、Ⅳ级间的油茶土壤硝态氮存在显著差异;发病等级Ⅰ、Ⅱ与0、Ⅲ、Ⅳ级间的油茶土壤氨态氮存在显著差异。

**表示差异显著(P<0.05)。下同。** indicates significant differences at the 0.05 level. The same as below.图1 不同发病等级油茶的AMF定殖率Fig.1 Total AMF infection rate of C.oleifera with different disease grades
2.2 油茶根系的AMF定殖率及根际土壤孢子密度
在梁河县5个发病等级的油茶根系和根际土壤中均检测到AMF的存在,不同发病等级油茶AMF的定殖率和孢子密度存在差异。由图1可知,病害等级为0级的油茶AMF的定殖率最高,病害等级为Ⅳ级的油茶AMF定殖率最低。由图2可知,在每20 g病害等级不同的油茶根际风干土壤中,AMF孢子密度均在30个以上,病害等级为0级的油茶AMF的孢子密度最高,病害等级为Ⅳ级的油茶AMF的孢子密度最低,这与油茶根系 AMF 定殖状况结果相一致。
2.3 油茶根系以及根际土壤AMF分子多样性分析
2.3.1 不同病害等级油茶根系及根际土壤AMF测序数据统计分析 对病害等级不同的油茶根系以及根际土壤进行高通量分子测序(表3~4),在病健植株的根系中共获得有效序列212 150个,HR0、DR1、DR2、DR3和DR4的有效序列分别为45 621、43 551、40 267、39 534和43 177,平均长度为216~218 bp。在油茶植株样本的根际土壤中共获得有效序列241 631个,HS0、DS1、DS2、DS3和DS4的有效序列分别是49 595、52 158、49 093、41 334和49 451,平均长度为218~224 bp。所有有效序列被划分成203个OTU,其中根系的OTU数量为112,HR0、DR1、DR2、DR3和DR4的OTU数量分别为47、61、30、21和31;根际土壤中OTU数量为186,HS0、DS1、DS2、DS3和DS4的OTU数量分别为99、75、69、69和62。
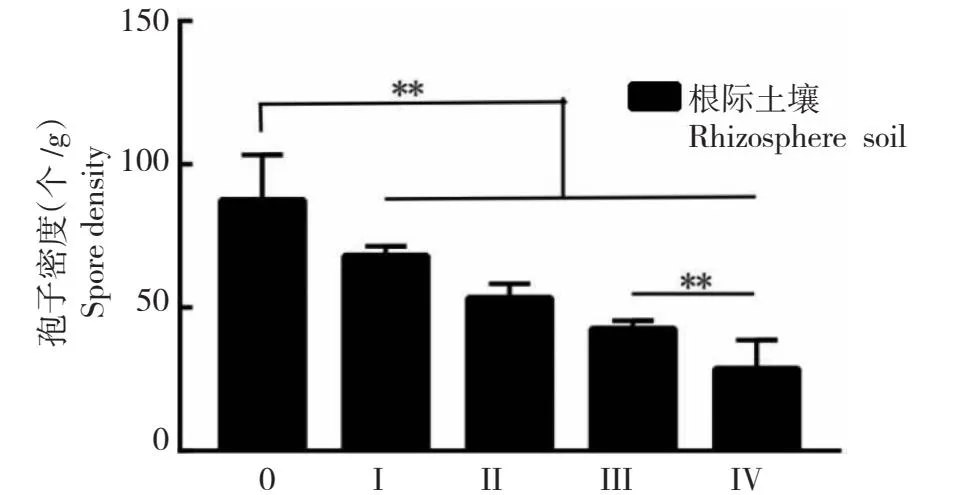
图2 不同发病等级油茶的AMF孢子密度Fig.2 AMF spore density of C.oleifera with different disease grades
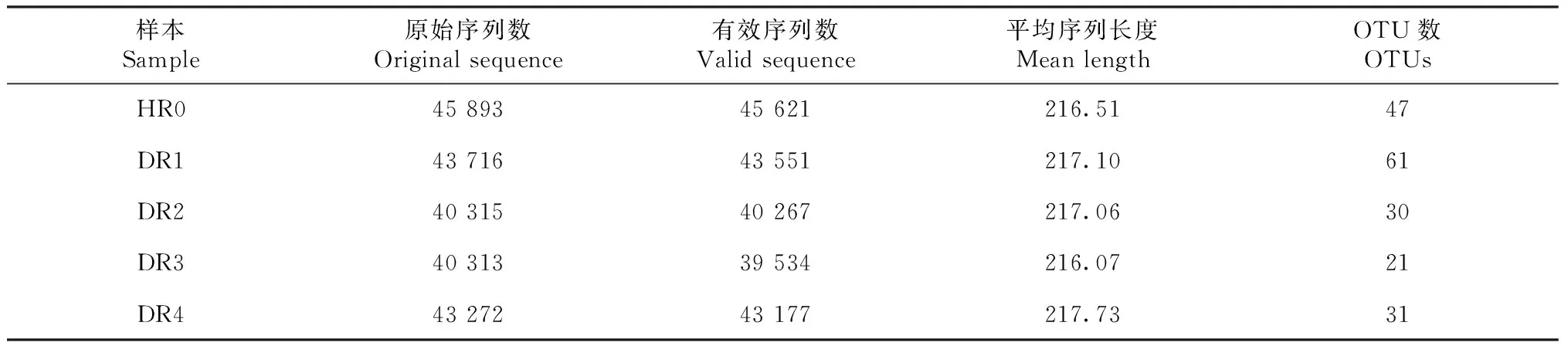
表3 植株样本根系AMF高通量测序数据
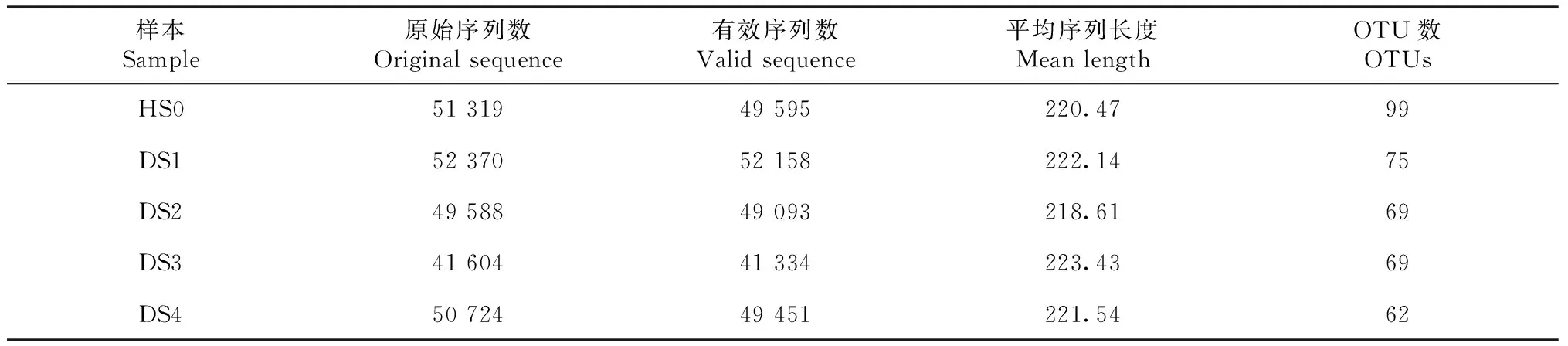
表4 植株样本根际土壤AMF高通量测序数据
2.3.2 不同病害等级油茶根系及根系土壤AMF稀释曲线比较分析 稀释曲线可用于比较不同测序样品中物种的丰富度,也可用于解释样品中测序数据的数量是否足够以及是否需要添加测试数据。由图3可知,采样点检测样品的稀释曲线最终趋于稳定,表明测序数据量足够大,能够客观反映群落的真实情况,可以进行数据分析。由图3-A可知,不同病害等级的油茶根系内AMF的Shannon指数HR0>DR1>DR2>DR3>DR4,说明不同病害程度的油茶根系中AMF的多样性由高到低依次为HR0、DR1、DR2、DR3和DR4,即油茶越健康,根系内AMF的多样性越丰富(Pearson相关系数为0.896),且呈显著相关性(P<0.05);由图3-B可知,不同病害等级的油茶根际土壤中AMF的Shannon指数HS0>DS4>DS2>DS3>DS1,说明不同病害程度的油茶根际土壤中AMF的多样性由高到低依次为HS0、DS4、DS2、DS3和DS1,即油茶的发病程度与土壤中AMF的多样性无关(Pearson相关系数为0.059),这与根系中AMF的多样性指数状况不一致。
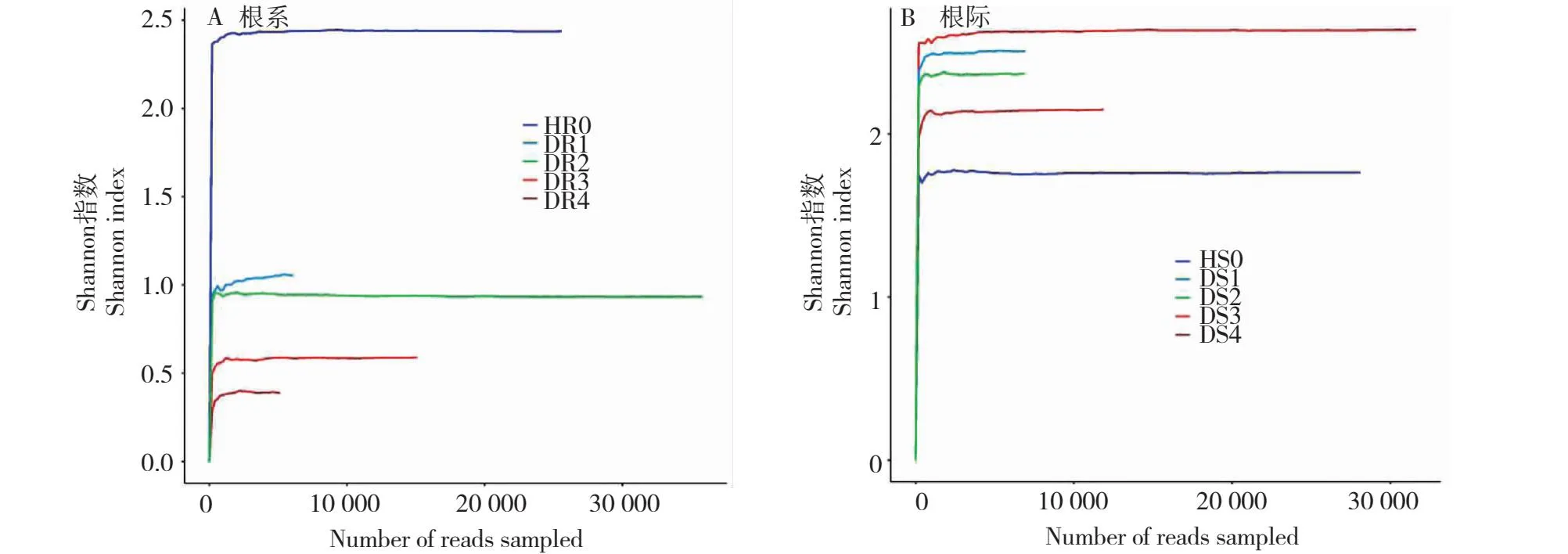
图3 不同病害程度的油茶根系和根际土壤Shannon指数稀释曲线Fig.3 Shannon index dilution curves of C.oleifera roots and rhizosphere soil with different disease degrees
2.3.3 不同病害等级油茶根系以及根际土壤 AMF 多样性比较分析 α多样性可用来反映物种丰富度及多样性。由表5可知,油茶根系中:不同病害等级的油茶根系内AMF的OTUs、均匀度指数Ace和丰富度指数Chao1均是DR1最大,其次是HR0,说明健康和发病程度为Ⅰ级的油茶根系中AMF的丰度较高。病害等级为Ⅳ(DR4)的Shannon指数最低,同时Simpson指数最高,说明在根系中连续抽到相同 AMF 物种的概率较高,其 AMF 多样性较低。而健康油茶(HR0)的Shannon指数最高,同时Simpson指数又是最低,说明在健康根系中连续抽到相同 AMF 物种的概率较低,其 AMF 多样性较高,HR0多样性指数是DR4的6.26倍。由表6可知,油茶根际土壤中:不同病害等级油茶的根际土壤AMF的OTU、均匀度指数Ace、丰富度指数Chao1以及Shannon指数均是健康油茶(HS0)的最高,说明HS0中AMF的丰富度以及多样性最高。可能病害发生后,会不同程度地降低油茶根系及根际土壤中AMF的丰富度和多样性。
2.3.4 不同病害等级油茶根系以及根际土壤AMF的OTU分类 韦恩图可以直观地展示不同病害等级下油茶根系和根际土壤中 AMF 群落间 OTUs物种组成的共有及差异情况。油茶根系中:5个病害等级不同的油茶共有112个OTUs,共享OTU数4个,HR0、DR1、DR2、DR3和DR4分别有47、61、30、29和31个OTUs,HR0、DR1、DR2、DR3和DR4特有的OTUs分别为21、18、11、2和7,HR0特有的OTU数最多,占所有OTUs的18.75%(图4-A)。油茶根际土壤中:5个病害等级不同的油茶共有186个OTUs,共享OTU数14个,HS0、DS1、DS2、DS3和DS4分别有99、75、69、69和62个OTUs,HS0、DS1、DS2、DS3和DS4特有的OTU数分别为23、30、17、10和7,DS1特有的OTU数最多,占所有OTUs的16.13%(图4-B)。5个不同病害等级的根际土壤中共同包含的OTU数比根系中的多10个。5个不同病害等级的根系和根际土壤中共享2个OTUs,HR0、DR1、DR2、DR4、HS0、DS1、DS2、DS3和DS4特有OTUs分别为1、4、4、5、15、21、12、8和3个,DR3没有(图4-C)。
2.3.5 油茶根系以及根际土壤AMF的群落组成 在油茶的根系以及根际土壤中共获得203个AMF-OTUs,排除不可归类的,总共将其划分为1门1纲4目7科(表7)。在门和纲水平上分别是球囊菌门 (Glomeromycota)和球囊菌纲(Glomeromycetes);在目水平共鉴定4个目:球囊霉目Glomerales,83.62%)、类球囊霉目(Paraglomerales,10.45%)、多样孢囊霉目(Diversisporales,3.68%)、原囊霉目(Archaeosporales,1.30%);在科水平,共鉴定出7个科:球囊霉科(Glomeraceae,83.38%)、近明球囊霉科(Claroideoglomeraceae,0.25%)、巨孢囊霉科(Gigasporaceae,3.00%)、无梗囊霉科(Acaulosporaceae,0.68%)、类球囊霉科(Paraglomerales,0.11%)、双型囊霉科(Ambisporaceae,0.92%)、原囊霉科(Archaeosporaceae,0.38%);未被注释到的归为“Others”。

表5 不同病害等级油茶根系AMF-α多样性指数
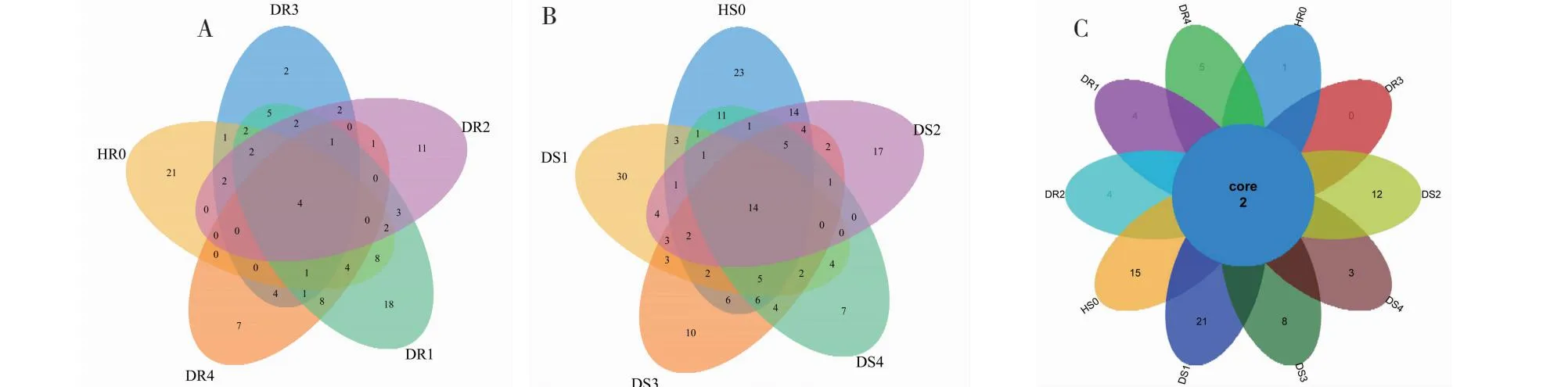
A:根系;B:根际土壤;C:根系和根际土壤。A: Fibrous roots; B: Rhizosphere soil; C: Root system and rhizosphere soil.图4 AMF-OTU比较分析Fig.4 Comparison of AMF-OTU
2.3.6 不同病害等级油茶的根系以及根际土壤AMF在属水平比较分析 将未定名和未分类的AMF归入“Others”,不同病害等级下油茶根系AMF群落由根孢囊霉属Rhizophagus、球囊霉属(Glomus)、双型囊霉属(Ambispora)和巨孢囊霉属(Gigaspor) 4个属组成;根际土壤AMF群落由根孢囊霉属、球囊霉属、双型囊霉属、巨孢囊霉属和类球囊霉属(Paraglomus)5个属组成。由图5-A可知,油茶根系中,发病等级不同的5个样本HR0、DR1、DR2、DR3和DR4中分别检测到4、4、1、1和3个属,病害等级为0、Ⅰ的油茶根系中AMF优势属(丰度>1%)均为根孢囊霉属(1.35%~11.44%);与健康油茶相比,Ⅰ、Ⅱ、Ⅲ和Ⅳ级球囊霉属丰度分别降低95.9%、98.6%、100%和97.1%,根孢囊霉属丰度别降低88.2%、100%、100%和100%。由图5-B可知,油茶根际土壤中:发病等级不同的5个样本HS0、DS1、DS2、DS3和DS4中分别检测到5、5、4、5和4个属,HS0的优势属为球囊霉属(1.40%),DS1的优势属为球囊霉属(12.40%)和巨孢囊霉属(3.65%),DS3的优势属为球囊霉属(16.05%)和双型囊霉属(3.78%),DS4的优势属为根孢囊霉属(44.13%)、球囊霉属(3.38%)、双型囊霉属(2.59%)和类球囊霉属(1.14%);发病程度最严重的油茶根际土壤(DS4)内根孢囊霉属(Rhizophagus)明显多于其他病害等级内的。此外类球囊霉属(Paraglomus)只出现在根际土壤中。
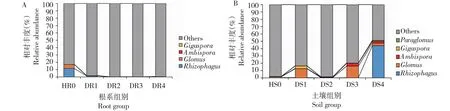
图5 不同病害等级油茶根系以及根际土壤 AMF 在属水平上聚类树Fig.5 Cluster tree of AMF in C.oleifera roots and rhizosphere soil of different disease grades at genus level
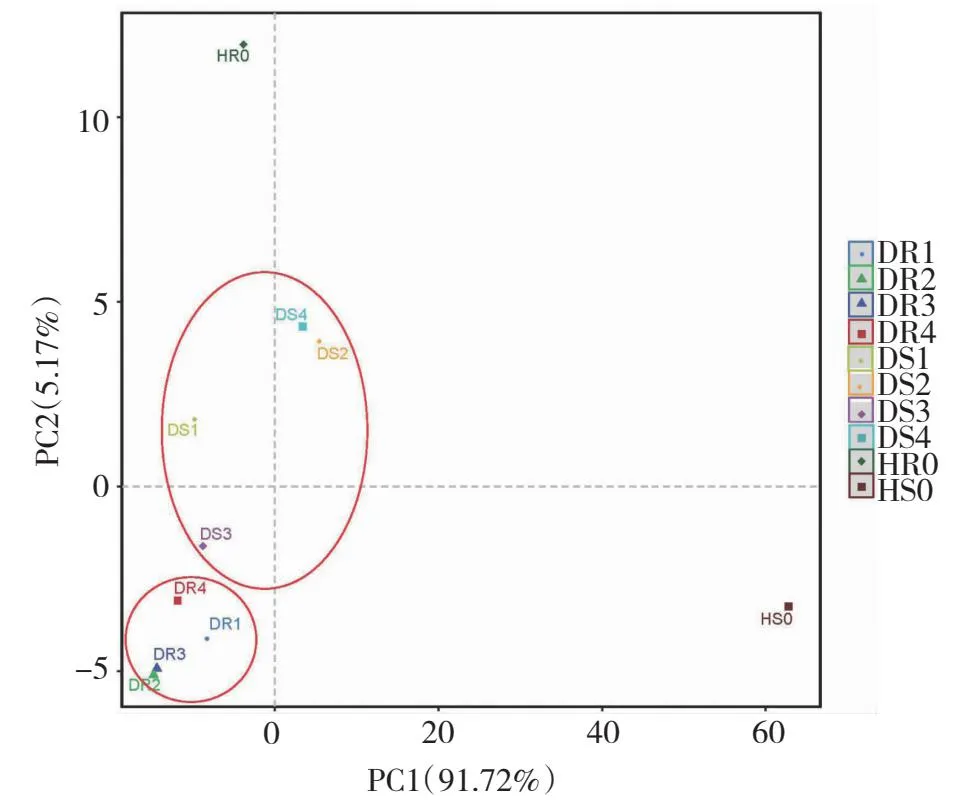
图6 不同病害等级油茶根系和根际土壤中AMF的β多样性的主成分分析(PCA)Fig.6 Principal component analysis (PCA) of AMF β diversity in C.oleifera roots and rhizosphere soil with different disease grades
2.3.7 不同病害等级油茶根系以及根际土壤AMF群落PCA分析 根系和根际土壤主成分分析结果表明PC1和PC2的累计贡献率可达96.89%。由图6可知,不同病害等级的根系和根际土壤存在着一定差异,油茶根系及根际土壤中患病样品(DR1、DR2、DR3、DR4、DS1、DS2、DS3、DS4)间距离较近,AMF组成较相似,而油茶根系及根际土壤中健康样本(HR0、HS0)间距离较远,AMF的组成则存在较大差异,说明病害等级为0级时AMF的结构与病害等级为Ⅰ、Ⅱ、Ⅲ、Ⅳ的差异较大,患病能改变油茶根际AMF结构,使AMF的结构趋于相似。
2.4 AMF群落组成与土壤因子RDA分析
从图7可知,土壤因子对AMF群落变异的解释量超过96.89%。第一排序轴解释群落变化的60.34%,而第二排序轴解释群落变化的36.55%。对AMF群落影响从大到小的环境因子是有机质>pH>速效钾>有效磷。AMF和环境因子相关性分析(表8)表明,有机质与均匀度指数呈显著负相关,与AMF定殖率和孢子密度呈极显著负相关;全氮与AMF定殖率呈显著负相关,与孢子密度呈极显著负相关;孢子密度与全钾呈极显著正相关,与硝态氮呈显著负相关,其余土壤理化性质与Ace指数、AMF定殖率和孢子密度无显著相关。所有土壤理化性质与Shannon指数 和Chao指数无显著相关性。
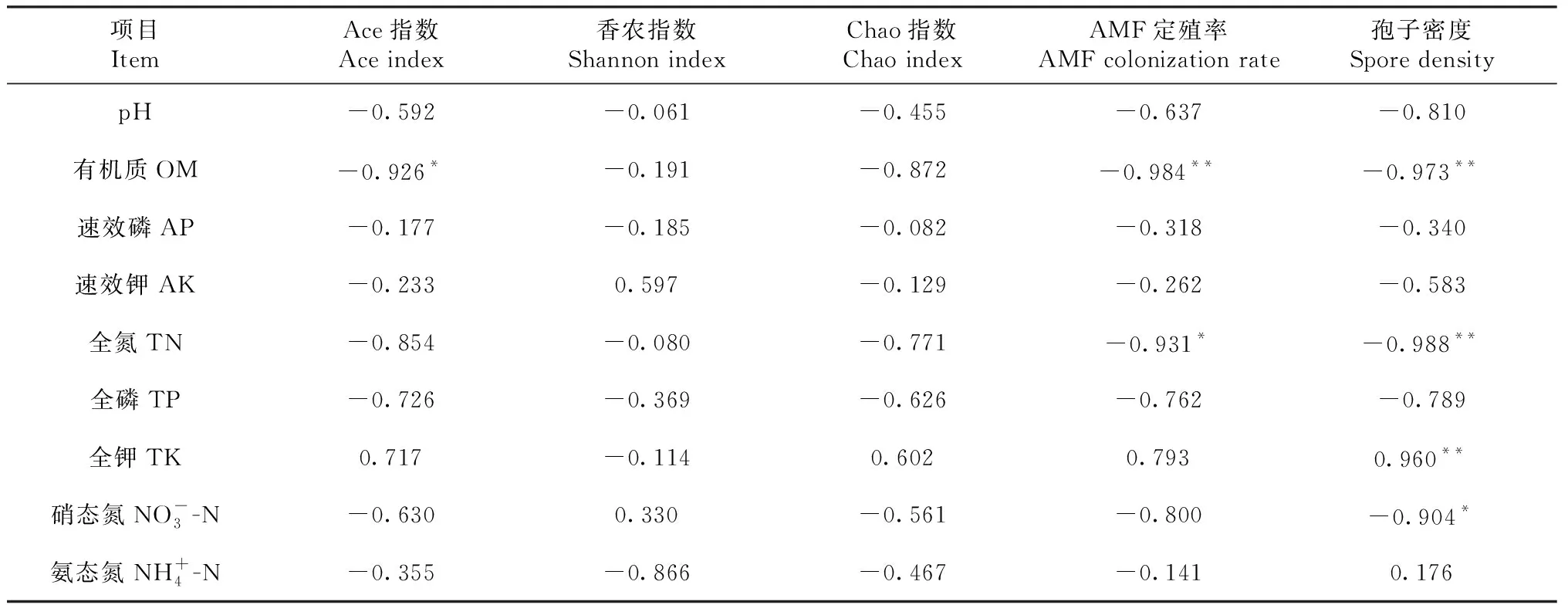
表8 AMF物种多样性和环境因子相关性分析
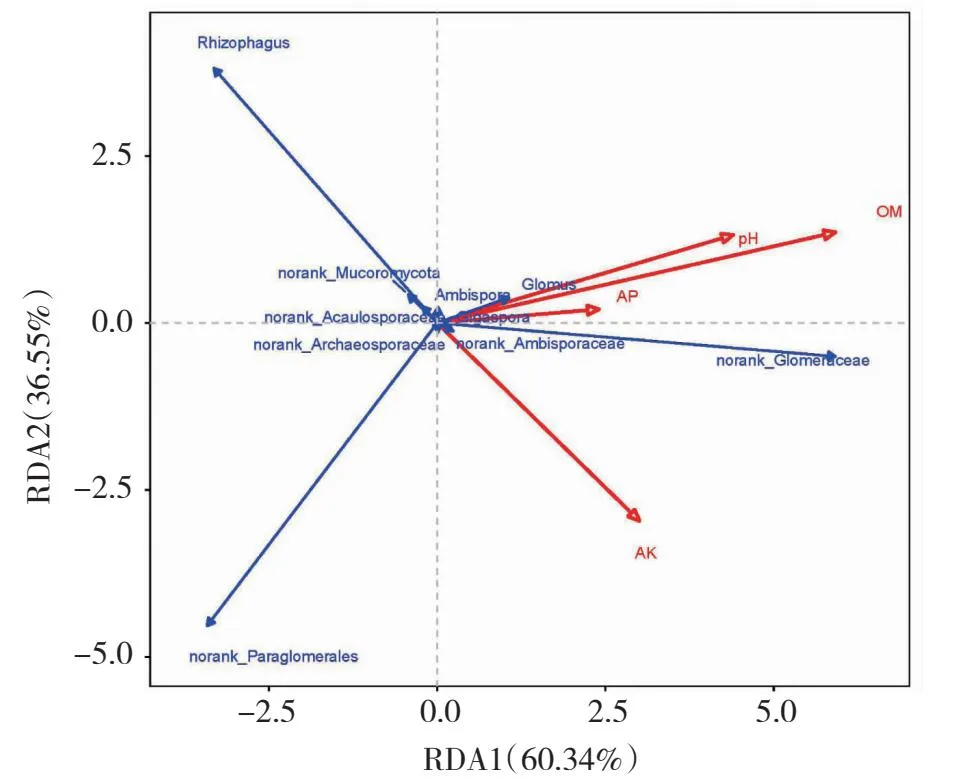
图7 不同病害等级油茶根际土壤AMF属水平群落及土壤理化因子的RDA分析Fig.7 RDA analysis of AMF horizontal community and soil physical and chemical factors in rhizosphere soil of C.oleifera at different disease levels
3 讨 论
AMF作为一类有益真菌,在陆地土壤微生物生态系统中扮演着重要的角色。它通过与寄主植物建立稳定的互惠菌根共生体,来促进植物生长发育,改善植株生理特性,从而提高植物对非生物胁迫和生物如病虫害的抗性等[20]。AMF在显微镜下难以鉴定区分,其缺乏明显的物种特征,因而研究AMF多样性存在一定的困难[21],同时,传统分离方法分离AMF孢子的效率低,不能在短时间内获得全面、大量的孢子信息。而Illumina高通量测序具有高通量和高精度的优点,可以克服这些方法的局限性,扩大对油茶AMF类群的了解。油茶是能与AMF建立共生关系的作物之一,对油茶根际土壤中AMF群落的研究已有开展,先前的研究表明在油茶根际土壤内存在着丰富的AMF资源,如邓小军等[22]采用湿筛倾析-蔗糖离心法在湖南省油茶林样地共分离到3属8种AMF;林宇岚等[1]采用Illumina Miseq高通量测序技术在江西省的5个油茶品种中获得4目10科12属的AMF。本研究利用Illumina高通量测序技术,在健康和患病的油茶根系以及根际土壤中共检测到203个AMF-OTUs,属于4目7科5属(属水平将未定名和未分类的AMF归入“Others”)。
AMF分布受到根际土壤和环境的影响,本研究发现,无论是健康植株还是患病植株,根际土壤AMF多样性和丰富度指数普遍高于根系,这与王化秋等[23]的研究结果一致。 AMF 定殖率和孢子密度与植物病害的发病率、病情指数呈显著负相关[24],当植物受到病原菌侵染时可能会影响孢子的密度以及定殖率,本研究发现,随着病害等级的上升,AMF的定殖率和孢子密度显著下降,即油茶发病的情况越严重AMF的定殖率和孢子密度越低,裴妍等[21]也同样证实健康烟株AMF的定殖率和孢子密度显著大于患黑胫病的烟株。本研究发现,油茶根系中,较健康(HR0和DR1)的AMF总OTU数和特有OTUs大于患病较严重(DR2、DR3和DR4),说明感根腐病会降低油茶根系内的OTUs。较为健康(HR0和DR1)的油茶,根系内AMF的 Ace、Chao 指数更高;Shannon指数随病害等级的升高而降低,即感病会降低油茶根系 AMF 的多样性;宋放等[25]同样也证实柑橘受黄龙病病原侵染后根系中AMF的物种组成和相对丰度会发生改变。油茶根际土壤中,随着病害程度的加重,AMF类群的OTUs呈现出逐级递减的趋势,且健康的油茶根际土壤内AMF的 Ace、Chao 指数最高,即患根腐病会降低根际土壤中AMF的丰富度;但Shannon指数与发病程度的关系不明显,即患病对油茶根际土壤内AMF的多样性影响较弱;吕燕等[26]研究发现,宁夏枸杞根区土壤 AMF多样性不受根腐病菌影响,AMF 多样性变化不明显,这与本研究结果一致。综合上述结果可知油茶患病能够降低根系内AMF的多样性,改变AMF的群落结构;油茶患病能够降低根际土壤内AMF的丰度,改变AMF的群落结构,但不影响其AMF群落的多样性。由此推测,根腐病病原菌和AMF在油茶根内存在明显的竞争作用,一方面,AMF菌丝能通过竞争生态位点,来减少病原菌的侵占位点,一定程度上抑制病原菌对植物根系的侵染[27];另一方面,AMF通过与病原菌直接的营养竞争,提升寄主抵御病原菌的能力[28]。
AMF孢子可以在没有寄主的情况下萌发和生长,但它们的菌丝生长非常有限,必须依赖宿主植物才能完成其生活史[29]。本研究发现,根系中,油茶患根腐病后,球囊霉属Glomus和根孢囊霉属Rhizophagus的丰度降低。刘先良等[30]发现接种根内球囊霉(G.intraradices)和摩西球囊霉(G.mosseae)的烟草菌根苗对青枯病的抗性明显增强,李柄霖等[31]发现接种根内根孢囊霉(R.intraradices)可以降低根腐病的发病率和病害指数,增加相关防御酶的活性以增强植物对根腐病的抗性,减少根腐病的发生和危害。表明球囊霉属和根孢囊霉属可能会提高植物的抗性,但这两类AMF类群能否提高油茶对根腐病的抗性还需进一步研究。根系招募土壤真菌是具有选择性的[32],本研究发现,在健康油茶根系内丰度最高的根孢囊霉属,在发病等级最严重的根际土壤中丰度最高,由此推测,油茶可能在受到病原菌侵染后会向环境中分泌相关物质,招募有益菌。另外,类球囊霉属(Paraglomus)只在根际土中出现,这可能是因为AMF对寄主植物的侵染有一定的偏好性。PCA结果表明,患病会导致AMF的群落发生改变,根系及根际土壤中的AMF群落组成在受到病原菌侵染后,结构趋于相似,发病后的油茶AMF群落与健康油茶存在较大差异,这与裴妍等[21]研究结果一致。
本研究表明,土壤有机质与AMF的Ace指数、AMF定殖率、孢子密度呈显著负相关,盛敏等[33]研究发现AMF种的丰度和多样性指数随有机质的增加而降低,但刘敏等[34]研究发现,土壤有机质与孢子密度呈正相关,这可能是土壤有机质含量对AMF分布的影响有一个阈值,当土壤有机质含量太高时,肥力过剩,会抑制菌根和AMF生长,导致AMF丰度下降[35]。土壤全氮含量与AMF定殖率和孢子密度呈显著负相关性,低浓度下的氮会促进AMF生长,与前人研究结果一致[36]。此外,有研究表明钾元素与AMF定殖呈正相关,也有研究发现根系全钾含量与AMF群落的多样性和丰富度呈显著正相关[23],本研究发现,全钾与孢子密度呈极强正相关,这可能因为AMF的外生菌丝可以延伸到钾的亏缺区,吸收利用空间上对植物根系无效的那部分钾元素使得孢子密度增加。在土壤中,有效养分是植物可以直接吸收和利用的养分形式,与AMF密切相关。本研究发现,硝态氮与孢子密度呈显著负相关。裴妍等[21]研究发现速效氮与根系AMF多样性指数间呈负相关,均证实了土壤速效养分与AMF的负相关性。尽管速效磷通常是调控AMF定殖植物的主要因素,但是本研究中并没有发现速效磷与AMF的相关性,推测在一定范围内随着土壤磷含量的增加,将会促进AMF的定殖及生长发育[37];当超过或低于一定范围时,不利于AMF的生长发育,AMF的定殖率和多样性就会减少[38]。
4 结 论
从不同病害等级的油茶根系和根际土壤来看,根系中AMF多样性和丰富度指数低于根际土壤。随着病害程度的加重,AMF的定殖率与孢子密度下降,即油茶病害的严重程度与定殖率以及孢子密度呈负相关关系。根系内,随着油茶病害程度的加重,AMF的群落结构发生改变,丰富度以及多样性下降;根际土壤内,随着油茶病害程度的加重,AMF的群落结构发生改变,丰富度下降,但多样性变化不明显。土壤理化性质影响AMF定殖与孢子密度,但不影响AMF的多样性。本研究为筛选具有拮抗作用的 AMF 菌剂防治油茶病害奠定了理论基础。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2017年21期)2018-01-03
现代园艺(2017年21期)2018-01-03
中国西部(2017年4期)2017-04-26
湖南农业(2017年1期)2017-03-20
中国蔬菜(2016年8期)2017-01-15
渔业致富指南(2016年12期)2016-11-11
科学大众(中学)(2015年9期)2015-10-12
少儿科学周刊·少年版(2015年3期)2015-07-07
