
青海云杉不同演替阶段林下草本多样性特征及其环境解释
2024-01-17 03:02邹星晨王欣苗左亚凡张泽鑫贺康宁
生态学报 2023年24期
邹星晨,王欣苗,左亚凡,张泽鑫,贺康宁,*
1 北京林业大学水土保持学院,水土保持国家林业局重点实验室,北京 100083 2 北京市水土保持工程技术研究中心,北京 100083 3 水利部综合事业局,北京 100053
新世纪以来,全球变暖、环境恶化和人类活动的加剧导致了生物多样性的锐减[1]。生物多样性的保护已成为森林生态领域的研究热点。林下植被作为森林生态系统一个重要组成成分,在维持生态系统物种多样性方面有不可忽视的作用[2]。有研究表明:林下草本虽然生物量占比不高,但其物种多样性却能占据森林生态系统的90%[3]。影响草本多样性的因素众多,近年来,有关草本多样性的研究多集中在分析气候、林分结构和土壤因子对草本多样性的影响,并比较不同影响因素下草本多样性的差异。这种仅仅分析单个因子对草本多样性的研究,难以揭示影响因子之间的耦合关系,难以全面反映影响草本多样性的机理。有研究表明:林下草本丰富度,不仅受到林分结构等生物因子的影响,还与土壤肥力等非生物因子密切相关[4]。前人研究表明:树高、胸径、林分密度、郁闭度等林分结构因子和土壤养分、水分会显著影响林下草本多样性[4-6]。探究影响林下草本多样性的主要因素对维持森林生态系统稳定具有重要意义。
青海云杉(Piceacrassifolia)是祁连山地区的主要森林类型,对于维系该地区的物种多样性、改善区域小气候、涵养水源和维持生态平衡等方面均有着至关重要的作用[7-8]。随着天然林保护工程的实施,经过20年的封禁治理,林区内现存大量白桦(Betulaplatyphylla)、红桦(Betulaalbosinensis)、山杨(Populusdavidiana)等天然林,形成了以青海云杉为顶级群落的完整演替序列,是研究青海云杉演替过程的良好场所[9]。以往对青海云杉的研究多集中在土壤质量[10]、生态化学计量[11-12]、林分更新[13]、水源涵养[14]等方面,而关于青海云杉林下草本影响因素和演替过程中的草本多样性变化的研究却鲜有报道。
因此,本研究以阔叶纯林(白桦纯林、红桦纯林)→阔叶混交林(白桦×山杨、红桦×山杨)→针阔混交林(白桦×云杉、红桦×云杉)→常绿针叶林(青海云杉纯林)完整的演替序列,以空间代替时间的方法,通过物种重要值、物种丰富度、Simpson指数、Shannon-Wiener指数和Pielou指数等多样性指数对比分析,比较不同演替阶段的草本多样性特征,探究草本多样性的影响因子,以期为祁连山地区物种多样性保护和生态修复提供理论依据。
1 研究区概况
研究区位于青海省海东市互助县东北部,地理坐标为102°6′-102°43′E, 36°42′-37°60′N。该区处于青藏和黄土高原的过渡地带,属于温带大陆性季风气候,年平均气温5.8℃,年平均降水量477.4mm。林区内生物资源丰富,主要乔木树种有青海云杉、白桦、红桦、山杨等;灌木主要有小檗(Berberisamurensis)、金露梅(Dasiphorafruticosa)、银露梅(Dasiphoraglabra)等,草本主要有:野草莓(Fragariavesca)、黑麦草(Loliumperenne)、披碱草(Elymusdahuricus)、拳参(Bistortaofficinalis)。
2 研究方法
2.1 样地设置与调查
2022年7月,在青海互助北山林场,采用典型抽样法,按照青海云杉演替序列,选择阔叶纯林(演替早期阶段)、阔叶混交林(演替早中期阶段)、针阔混交林(演替中后期阶段)和青海云杉纯林(顶级群落阶段),各6个共24个20m×20m的样地。记录样地的经纬度、海拔、坡度和坡向等基本信息,样地基本信息如表1所示。

表1 样地基本信息表Table 1 Basic information table of sample plots
2.2 生物与非生物指标测定
在各样方内对每株乔木进行每木检尺,测高仪测定每株树高、胸径尺测量胸径,使用鱼眼相机(WinSCANOPYTM 2010a) 获取冠层照片, 运用XLScanopy软件处理照片,计算叶面积指数和郁闭度。统计样地内主要树种的数量,推算各样地的林分密度信息。采用五点取样法,在各样地取0-20cm的土层混合土样带回实验室,运用环刀法测定土壤水分、采用重铬酸钾稀释热测定土壤有机质[15]、靛酚蓝比色法测定碱解氮、NaHCO3浸提-火焰光度法测定速效磷、醋酸铵浸提-火焰光度计法测定速效钾、电位法测定土壤pH[16]。林分因子代表生物变量、土壤养分代表非生物变量,基础数据如表2所示。
2.3 林下草本多样性调查
按五点取样法在各样地布设5个1m×1m的草本调查样方,参考中国在线植物志,记录植物拉丁文、科、属、种和生活习性等基本信息。统计计算各物种的数量、高度、盖度,并计算各物种的重要值。采用草本的重要值(IV)、物种丰富度(S)、Shannon-Wiener多样性指数(H)、Simpson多样性指数(D)和Pielou均匀度指数(J)来分析林下植被的物种多样性[8]。各指标计算公式如下:
J=-∑Pi×lnPi/lnS;
式中:RA表示物种的相对多度、RC表示相对盖度、RH表示物种的相对高度;Pi表示第i种的个数占样方内总物种的个数;S代表样方内的总物种数。
2.4 数据分析
首先利用Excel 2019和SPSS 22.0对数据进行统计分析,采用单因素方法分析(One-way ANOVA)中的LSD(最小二乘法)和邓肯检验对不同演替类型下的林下草本多样性进行差异性检验。再用Spearman相关系数分析林下草本多样性与影响因子的相关性,最后采用CANOCO 5.0中的RDA冗余分析来探讨影响草本多样性的主要影响因子。采用Origin 2021绘图,Excel制表。
3 结果与分析
3.1 林下草本组成与重要值
林下草本组成如图1所示,本研究共发现草本31科64属81种,其中阔叶纯林(Ⅰ)草本植物共21科33属42种;阔叶混交林(Ⅱ)共22科40属50种;针阔混交林(Ⅲ)共22科42属51种;云杉纯林(Ⅳ)共15科21属23种。针阔混交林(Ⅲ)下的草本种类最丰富,而云杉纯林(Ⅳ)下的草本种类最少。各林分类型下主要草本的重要值如表3所示:各林分类型之间的优势草种存在部分重叠,黑麦草、山尖子、野草莓在四种林分类型下均有分布。其中,黑麦草在林分Ⅰ、Ⅲ中的重要值最大,其重要值为:23%、24%;野草莓在林分Ⅱ的重要值最大,其值为26%;冷蕨在林分Ⅳ的重要值最大,其值为33%。
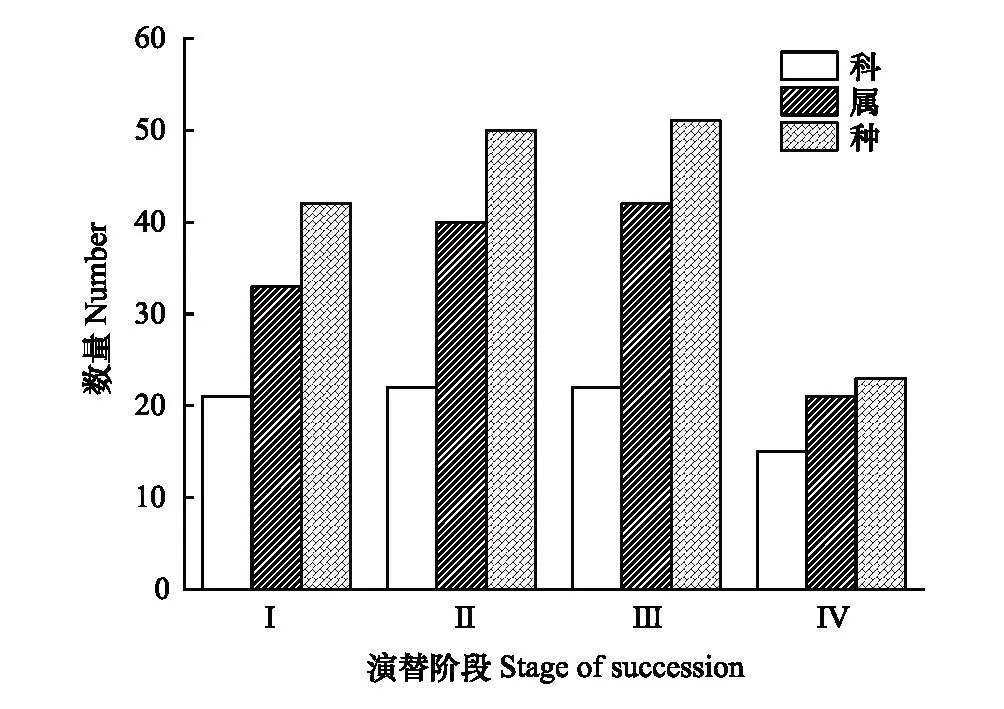
图1 不同演替阶段林下草本组成Fig.1 Understory herbaceous composition at different succession stages

表3 草本主要物种组成及其重要值Table 3 Main species composition of herbs and their important values
3.2 不同演替阶段林下草本多样性特征
不同演替阶段草本物种多样性如图2所示,阶段Ⅰ的物种丰富度均值为11.33,阶段Ⅱ丰富度的均值为16.83,阶段Ⅲ为19.29,阶段Ⅳ为8;物种丰富度和物种多样性指数在不同演替阶段之间存在显著差异,各指标变化趋势一致,随着青海云杉演替正方向呈现先增加后减小的趋势,各多样性指标均在Ⅲ最大,在Ⅳ最小。

图2 不同演替阶段草本多样性Fig.2 Herbaceous diversity at different succession stages不同字母表示差异显著
3.3 草本多样性与影响因素的相关分析
草本多样性指标与生物和非生物因素的相关分析如表4所示:生物因子中,郁闭度与草本丰富度呈显著正相关(P<0.05);林分密度与Simpson指数和Pielou均匀度指数显著负相关;平均树高与物种丰富度呈极显著负相关(P<0.01),与Shannon-Wiener指数显著负相关;平均胸径与物种丰富度、Shannon-Wiener指数极显著负相关,与Simpson指数显著负相关;LAI与4个多样性指标均显著负相关。非生物因子中,土壤水分与Simpson指数、Shannon-Wiener指数、Pielou均匀度指数呈极显著负相关;碱解氮、速效磷与物种丰富度显著正相关;速效钾与物种丰富度呈显著正相关,与Shannon-Wiener指数呈显著正相关;土壤有机质与丰富度、Simpson指数和Shannon-Wiener指数显著正相关;土壤pH均大于7且与多样性指标无显著相关性。

表4 草本多样性与影响因素Spearman相关分析表Table 4 Spearman correlation analysis of herbaceous diversity and influencing factors
3.4 草本多样性的驱动因子分析
对各个环境因子与草本多样性指数进行RDA冗余分析(图3)结果表明:第一主轴解释变量为74.95%,第二主轴解释变量为7.24%,两轴共解释草本多样性与环境关系的82.19%,能较好的反映排序的大部分信息。实心箭头代表草本多样性指标,空心箭头代表影响因子。影响因子的连线长度代表该因子与草本多样性指标的影响程度,连线越长表示该因子影响越大。箭头之间夹角的余弦值表示相关系数[17]。其中,林分密度、LAI、胸径、树高与草本多样性指标S,D,H,J的夹角为钝角且LAI连线最长,这表明:与草本多样性负相关的指标中,LAI具有较强的解释能力。土壤水分和养分等指标与草本多样性指标的夹角为锐角,土壤水分、速效钾和土壤有机质的连线较长,这表明:速效钾、土壤有机质和土壤水分对草本多样性具有较强的解释能力。采用蒙特卡洛置换检验对11个影响因子进行筛选排序(表5)结果表明:速效钾变量解释度为27%、土壤有机质解释度为28.5%、LAI变量解释度为13.2%、土壤有机碳、速效钾和LAI对草本多样性指数具有显著影响(P<0.05),是影响草本多样性的主要因素。

图3 林下草本多样性影响因子RDA分析结果图Fig.3 RDA analysis results of understory herb diversityS:物种丰富度 Species richness index;D:Simpson多样性指数 Simpson index; H:Shannon-Wienerd多样性指数 Shannon-Wiener index; J: Pielou均匀度指数 Pielou evenness index;CD:郁闭度 Canopy density;SD:林分密度 Stand density; MTH:平均树高 Mean tree height; DBH:平均胸径 Mean diameter at breast height; LAI:叶面积指数 Leaf area index; SWC:土壤含水量 Soil water content; AN:碱解氮 Alkali-hydrolyzable nitrogen;AP:速效磷 Available phosphorus; AK:速效钾Available potassium; SOM:土壤有机质 Soil organic matter; pH:土壤酸碱度 Soil pH
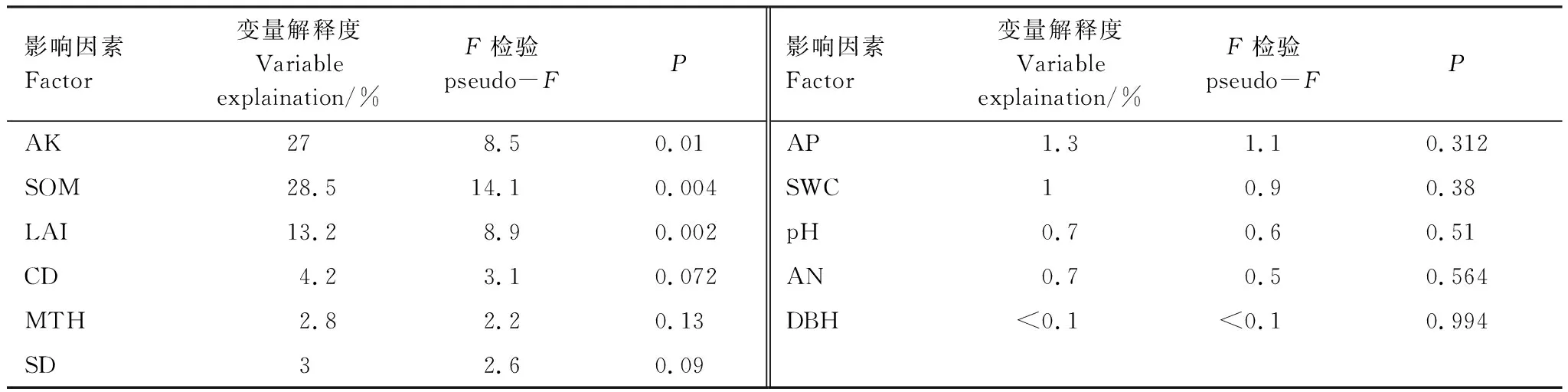
表5 生物非生物指标解释度与蒙特卡洛检验表Table 5 Biometric Interpretation and Monte Carlo Test
4 讨论
4.1 群落演替与草本多样性的关系
本文采用了空间代替时间的方法研究不同演替阶段下的草本多样性差异,此方法虽简单可行,但缺少时间维度的长期观测。今后可进行多年连续检测观察不同演替阶段的草本多样性的变化。研究表明:祁连山东部地区森林在不同演替阶段的林下草本组成和多样性存在显著差异。研究发现:黑麦草、野草莓是不同演替阶段的优势草种。这可能是黑麦草因其根系发达、分蘖能力强、繁殖力高,能适应各种林下环境[18];而野草莓通过匍匐茎能占据更多的生境空间,获取更多的养分和资源[19]。不同演替阶段下,林下草本优势种以阳生草本为主,而阴生草本随着演替的进展其重要值在不断增加(表3)。在青海云杉纯林阶段,冷蕨等阴生草本占据绝对优势,这主要是所调查的青海云杉纯林密度较大(表2),林内形成了偏阴生的环境且调查的样方多在阴坡(表1),光照时间短,水分较好,适合耐阴喜湿的草本生长[20]。在针阔混交林阶段,草本多样性指标达到峰值,这可能是一方面:针阔混交增加了生境异质性,为不同生活型草本的生长提供条件[21],另一方面,针阔混交林通过增加枯落物种类,促进林内养分循环,改良土壤养分状况[22],进而促进了林下草本的生长。本研究发现,在阔叶纯林向青海云杉的演替序列中,林下草本多样性呈先增加而后减少的规律。可能是一方面林分演替到青海云杉纯林阶段时,林内阴暗,气温低,湿度大,林下草本难以生长[23-24];另一方面青海云杉作为浅根性树种,其根系通过化感作用,产生能够抑制种子萌发所需的关键酶类, 抑制了林下草本的生长[25]。
4.2 生物因素对草本多样性的影响
Spearman相关分析(表4)和RDA冗余分析(图3)的结果表明:生物因子中,LAI是显著影响林下草本多样性的主要林分因子,LAI与林下草本多样性呈显著负相关。这可能是LAI代表冠层叶片生长状况,反映乔木对光的获取能力,乔木冠层通过叶片对光截获,抑制林下低矮草本对光的获取[26],而光作为植物生长必要的环境因子之一[27],光的缺乏会影响林下草本生长而导致林下草本多样性偏低。林分密度是进行林分调控的重要因子,林分密度可通过控制林下光照条件和种间竞争影响林下草本的生长状况[28-29]。本研究得出:林分密度的增加会导致物种多样性的降低,这与部分学者得出的结论相同[30-31]。此外,树高和胸径能反映乔木生长状况[32],是反映林冠结构的重要指标。乔木可通过林冠结构改变林内光照强度、林分湿度的等林内环境,影响林下草本植物的生长[33-34]。本研究中:树高和胸径与草本多样性负相关,这可能是乔木的胸径越大,树高越高,其冠幅也越大,乔木对土壤养分、水分的吸收能力越强,导致林下草本土壤水分和养分的缺乏,草本多样性降低[35-36]。
4.3 非生物因素对草本多样性的影响
非生物因子中,速效钾和土壤有机质是显著影响林下草本多样性的主要土壤养分。本研究发现速效钾与物种丰富度和Shannon-Wiener指数显著正相关,这可能与当地的气候和成土母岩有关[37]。土壤中的速效钾主要取决于土壤中微生物对矿物分解的活性,虽然祁连山地区土壤发育于玄武岩、火山岩等富含钾元素的矿物,但由于研究区普遍海拔较高,土壤微生物活性较低,使得土壤中的速效钾含量不高,成为限制该地区草本多样性的主要环境因子[38]。本研究得出:土壤有机质含量越高,林下草本多样性越好,可能是土壤有机质作为土壤养分的载体[39-40],通过矿化作用分解成能被林下草本吸收的矿物养分[41]。因此,有机质含量越高,能被植被吸收利用的养分就越多,草本多样性就越高[42]。土壤水分与草本多样性正相关,这是因为土壤水分作为限制植被生长的重要环境因子[43],水分的缺乏会抑制植物对养分的吸收[44]。氮、磷元素是组成植物蛋白质的主要元素,由于在土壤中供应受限,通常被认为是限制植物生长的重要因素[45]。杨学亭通过研究祁连山不同草地类型与养分的关系得出,氮元素与草本多样性正相关,磷元素是影响草本多样的关键因素[46],这与本文研究结果部分一致。然而,在本研究中却未体现磷元素的重要性,这可能是研究区磷元素含量较少,未能体现差异性(表2)。
5 结论
在阔叶纯林→阔叶混交林→针阔混交林→云杉纯林的演替序列过程中,草本多样性总体呈先增大而后减少的规律,并在针阔混交林达到最大,云杉纯林最小。LAI是影响草本多样性的主导生物因子,速效钾,土壤有机质是影响草本多样性的主要非生物因子。今后,在草本多样性低下的青海云杉林进行林分改造时,可以适当的调整林分密度和补植桦树以营造针阔混交林。在对祁连山东部地区的森林群落多样性研究过程中,可考虑加入角尺度、竞争指数、光照条件等其他环境因子的作用,以期为祁连山地区森林生态恢复和林分改造提供理论支持。
猜你喜欢
中华建设(2023年10期)2023-10-07
现代农村科技(2020年7期)2020-12-19
少儿美术(2020年8期)2020-11-06
森林工程(2018年4期)2018-08-04
启迪与智慧·教育版(2018年12期)2018-02-26
山西林业科技(2017年4期)2018-01-23
现代园艺(2017年13期)2018-01-19
方圆(2018年23期)2018-01-07
甘肃林业(2016年2期)2016-11-07
广西林业科学(2016年4期)2016-03-16
