
塑料垃圾在南极生物体内的污染现状研究进展
2024-01-15 11:58陈宗正方超妙星王芮薄军郑榕辉李渊林龙山张静
极地研究 2023年4期
陈宗正 方超 妙星 王芮 薄军 郑榕辉 李渊 林龙山 张静
(1 集美大学水产学院, 福建 厦门 361021;2 自然资源部第三海洋研究所, 福建 厦门 361005)
0 引言
塑料因其价格低廉、重量轻、坚固耐用、耐腐蚀, 并且具有很高的隔热和绝缘性能, 长期受到人们青睐[1]。塑料消费的不断增加也不可避免地导致了塑料垃圾的增加, 其中大部分通过工业排放、垃圾和陆地径流进入海洋环境[2]。塑料在海洋垃圾中占比高达80%~85%[3]。据统计, 2020年全球塑料产量高达3.67 亿吨[4], 但仅小于5%的塑料材料被回收[5], 由于其不易降解的特征和人类持续排放等原因, 导致塑料在海洋环境中得以积累。其中, 微塑料指粒径小于5 mm 的塑料碎片、细线、纤维、微球或薄膜[6], 该概念由Thompson 等[7]于2004 年首次提出, 并在2008 年由美国国家海洋与大气管理局(NOAA)主办的首届关于海洋微塑料碎片国际研讨会上确定了微塑料粒径5 mm 的尺寸上限[8]。
目前有关微塑料的研究区域主要集中在近岸海域并涵盖太平洋、大西洋、印度洋和北冰洋[6,9-10],但涉及南极地区的研究相对匮乏。南极由于其偏僻的地理位置和极端的气候条件, 一直以来被认为是遭受人类活动负面影响最少的区域, 但是随着极地旅游业的兴起和渔业捕捞的迅猛发展, 以及日益增多的南极各类科考活动, 南极海洋生态系统面临着外来物种引入、过度捕捞和环境污染的生态风险, 其中微塑料污染问题愈发受到人们关注。微塑料主要可以通过3 种不同的途径到达南极地区: (1)来自当地的人类活动, 例如旅游业的发展、科考活动的增多, 以及生活废水处理不当或随意排放等; (2)来自南极地区以外的大气、海洋环流或生物运输; (3)来自更北的水域, 由风暴驱动的扩散和全球变暖引起[11]。
对南极地区的微塑料研究最早可追溯到20世纪80 年代, 研究人员发现南极毛皮海狮(Arctocephalus gazella)被缠死在废弃的塑料垃圾中[12]。2017 年,Munari 等[13]首次在罗斯海区域不同深度的沉积物中发现高浓度的微塑料, 在海岸线位置则出现大量的橡胶和尼龙。2018 年, Reed 等[14]检测了南极罗瑟拉研究站附近沉积物中的微塑料浓度, 最高达5 个·(10 mL)-1。2019 年, Lacerda 等[15]报道了南极半岛海洋表面的塑料碎片浓度为1794 件·km-2,其中微塑料浓度达54%。2020 年, Kelly 等[16]研究发现南极东部海冰中的微塑料浓度约为11.71 个·L-1,且成分与大多数海洋中一致。Cunningham 等[12]还发现在南极半岛、南桑威奇群岛、南乔治亚的陆上沉积物微塑料平均浓度(湿重)分别为1.3±0.51、1.09±0.22 和1.04±0.39 个·g-1。
微塑料含有不同种类的化学物质, 如在塑料生产过程中往往会添加稳定剂和染料, 一旦进入环境, 微塑料就可以释放这些物质, 或从环境中吸收或吸附一些疏水性的有机化合物和重金属[17]。目前摄入微塑料对低营养级的消费者所造成的负面影响已经逐渐明朗, 例如, 微塑料会在鱼类的鳃、肝脏和肠道中积累并且扰乱脂质和能量的代谢[18], 造成乙酰胆碱酯酶活性降低[19], 吸附了污染物的微塑料对器官的危害更大等[20]。南极生物适应极端的环境条件, 具有独特的表型特征, 与不同纬度的物种相比它们更容易受到环境变化和污染物的影响[21]。本文总结了有关南极生物摄入塑料的相关研究并进行归类, 按营养级由低到高的顺序(无脊椎动物、鱼类、鸟类、海洋哺乳类)来评述目前南极生物摄入塑料制品的研究现状,探讨今后重点研究的方向, 可为国内外学者开展南极微塑料研究提供参考。
1 南极各营养级生物摄入塑料制品现状
1.1 无脊椎动物
微塑料因其尺寸大小与浮游生物近似, 故可以被海洋无脊椎动物以不同的方式摄食, 此前已在贻贝、蚯蚓和海参体内发现微塑料[22]。南极底栖生物群落主要依靠水体和沉积物中的碎屑来获取能量和营养物质, 因此存在摄入微塑料的风险。2020 年, Sfriso 等[22]首次对南极底栖生物进行微塑料污染研究, 选择罗斯海12 种具有不同捕食习性的大型底栖生物(表1), 发现约83%的生物样本含有微塑料, 微塑料平均丰度为0.7 个·mg-1(湿软组织)和1 个·个体-1, 粒径范围为33~1000 μm。经鉴定其中86%为尼龙, 5%为聚乙烯(Polyethylene,PE), 其余聚合物为聚甲醛(Polyformaldehyde,POM)、聚苯乙烯(Polystyrene)、聚丙烯(Polypropylene,PP)、酚醛树脂等。双壳类中微塑料丰度最高, 其次是腹足类、多毛类、端足类和腔肠动物。
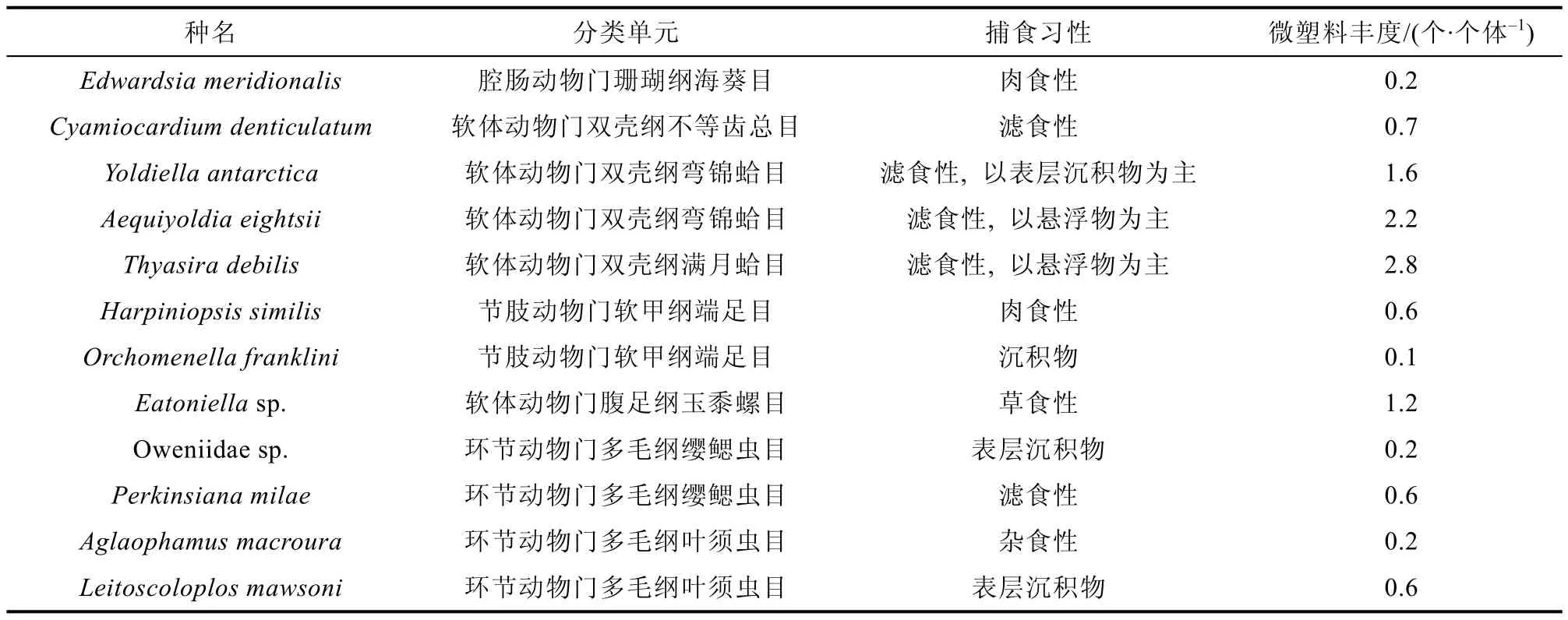
表1 12 种不同捕食习性的大型底栖生物体内微塑料丰度[22]Table 1. Microplastic content per individual in twelve macrobenthic species of different feeding strategies[22]
棘皮动物既可以直接吸收来自环境中的微塑料, 也可以通过营养转移间接吸收来自饵料生物中的微塑料[23]。其中海星隶属于棘皮动物门海星纲(Asteroidea), 是底栖生物群落中的顶级捕食者。2021 年, Cossi 等[17]在两种海星Henriciaobesa和Odontasterpenicillatus体内发现微塑料, 其中,H.obesa中平均丰度为3.34±4.13 个·g-1(湿软组织)和1.00±1.03 个·个体-1,O.penicillatus中平均丰度为1.94±2.09 个·g-1(湿软组织)和2.70±2.91 个·个体-1。微塑料以纤维状和碎片状为主, 所有微塑料呈蓝色, 其粒径范围为24~1340 μm, 且大多数小于1 mm, 经鉴定多为半合成纤维, 且在有色纤维和碎片中都附着有塑料工业中常见的酞菁颜料。两种海星体内微塑料丰度差异或许与生活习性有关,Henricia属通常摄食环境中的悬浮颗粒[24], 而Odontaster属则多为食肉动物, 通常以海豹肉、鱼类和其他棘皮动物为食[25]。目前, 摄食方式对水生生物摄入微塑料丰度的影响尚不确定, 有的学者认为摄食方式和摄入微塑料数量之间没有关系[26], 也有学者认为滤食性动物与杂食性和肉食性动物相比能摄入更多的微塑料[22,27], 但也有学者发现肉食性动物体内的微塑料丰度要高于滤食性动物[28-29]。
诺伊迈尔坚固海胆(Sterechinusneumayeri)是南极浅水区域最丰富的棘皮动物门海胆纲动物,分布在810 m 以浅水域[30]。它是一种杂食性动物,主要以藻类为食, 并在高初级生产力区域, 可将藻类转变成碎屑返还到生态系统中, 被食腐动物吸收[31]。该种海胆具有代谢速率低、生长速度缓慢[32]和遗传多样性低[33]的特点, 很容易受到环境变化的影响, 且容易富集环境中的微塑料[34]。南极沿岸考察站附近存在大量的人类活动, 这也加剧了其受微塑料污染的风险。2019 年, Bergami等[34]对乔治王岛南极海胆的研究发现, 纳米级的微塑料被摄入后会造成南极海胆体内吞噬细胞的吞噬功能受损, 且会干扰氧化应激和凋亡通路相关基因的表达, 从而破坏其免疫系统。
南极大磷虾(Euphausiasuperba)作为南极生态系统中的关键物种, 在南极食物网中发挥着重要作用, 其生物量极其丰富, 是南大洋众多高营养级捕食者的重要食物来源[35]。南极大磷虾主要以浮游植物为食, 但也会捕食樽海鞘、桡足类和磷虾等其他浮游动物[36]。南极大磷虾为滤食性动物, 这也导致其很容易摄入微塑料[37]。Dawson等[35]研究发现, 在实验室条件下, 南极大磷虾可以将摄入的31.5 μm 的微塑料粉碎成粒径小于1 μm的碎片, 且在没有藻类的条件下存在直接摄入微塑料的现象。
1.2 鱼类
南极海洋鱼类约有300 余种, 分属49 个科,仅占全球鱼类的1.3%[38-39]。1972 年, Carpenter等[40]首次在辐鳍鱼类的肠道和水体中发现了平均粒径1 mm 的聚苯乙烯小球。此后, 关于此类研究大多数集中于北半球, 而关于南半球甚至南大洋鱼类的微塑料研究则相对匮乏[41]。莫氏犬牙南极鱼(Dissostichusmawsoni)是南大洋深水区的鱼类, 通常生活在2200 m 水深处。2016 年, Cannon 等[41]在莫氏犬牙南极鱼胃肠道中发现了绿棕色、坚硬的塑料碎片, 经鉴定为丙烯酸树脂, 这表明南大洋的深水区也存在塑料污染。上述研究中发现的塑料都是通过肉眼识别出来的, 或许其体内还包含更小的微塑料和纤维。2022 年, Zhang等[42]首次对南极罗斯海和阿蒙森海的鲈形目鱼类的胃肠道进行微塑料研究, 发现罗斯海的鲈形目微塑料检出率和丰度(50%和1.286 个·个体-1)均高于阿蒙森海(36%和1.227 个·个体-1), 且罗斯海鲈形目胃肠道微塑料成分多为聚丙烯酰胺(Polyacrylamide, PAM), 粒径为100~200 μm, 阿蒙森海鲈形目胃肠道微塑料成分多为人造纤维(rayon), 粒径为500~1000 μm。两个海域鲈形目鱼类体内微塑料丰度出现差异的原因可能是不同的考察站、废水处理设施和渔业活动的排放, 以及洋流和局部环流等因素。鲈形目鱼类体内所含微塑料可能通过食物网传播对其他南极生物和人类健康产生不利影响[42]。
1.3 鸟类
自20 世纪70 年代初人们认识到海洋塑料污染问题以来, 海鸟吞食塑料颗粒的报告一直在稳步增加[43]。由于海鸟习惯接近海面和船舶后觅食,因此特别容易摄入塑料, 据统计, 约有78%的海鸟摄入过塑料制品[44]。在南极海洋生物中, 海鸟被认为是监测生态环境变化的最方便的生物指标之一, 也被建议作为许多海洋环境中塑料污染的指标[45], 这与其独特的生理结构有关, 以鹱形目(Procellariiformes)的海鸟为例, 它们的幽门括约肌狭窄且有一定的角度, 可以将难消化的物质保留在胃肠道中[46]。关于南极区域鸟类摄入微塑料的研究最早可以追溯到20 世纪80 年代, 有研究发现来自南极的候鸟鸽锯鹱(Pachyptiladesolata)前胃和砂囊中存在塑料碎片[43]。1984—1987 年,van Franeker 和Bell[47]通过调查南极洲阿德里岛的海燕, 发现黄蹼洋海燕(Oceaniresoceanicus)的胃含物中塑料浓度较高, 平均每只有4.4 个塑料颗粒, 且雏鸟体内的塑料浓度要大于成鸟。1996—2018 年, Perold 等[48]持续调查了亚南极区域的马里恩岛的鸟类巢穴, 获得了共计1486 件筑巢污染物, 大多数是硬质塑料和与渔业相关的塑料制品, 少量的食品包装及塑料袋等。1994—2019 年, Phillips 和Waluda[49]通过调查南乔治亚的鸟类栖息地和巢穴, 在漂泊信天翁(Diomedea exulans)、灰头信天翁(Thalassarchechrysostoma)和黑眉信天翁(Thalassarchemelanophris)的巢穴中分别收集到541、548 和124 件筑巢污染物, 其中漂泊信天翁摄入并带回的大多数是塑料袋(44.9%)、食品包装(23.0%)和手套(5.3%)等, 而灰头信天翁和黑眉信天翁摄入的大多数是各种类型的塑料(61.9%和62.1%)、瓶盖(25.3%和17.4%),其次是塑料袋(5.2%和9.1%)。此外还调查了它们雏鸟反刍的胃含物, 发现漂泊信天翁的塑料摄入率比黑眉信天翁高5.7 倍, 灰头信天翁的比黑眉信天翁的高1.4 倍。
南极企鹅, 如金图企鹅(Pygoscelispapua), 一直被认为是监测南极海洋生态系统污染程度的指示生物, 它们几乎全年都在自己的栖息地活动[45],和习惯迁徙的候鸟相比, 更能反映南极当地的微塑料污染状况。2019 年, Bessa 等[45]在南乔治亚岛和南奥克尼群岛上总共采集了80 个金图企鹅的粪便样本, 通过研究发现其中约20%含有微塑料,大部分呈纤维状, 平均长度为1266±1378 μm, 最小长度仅为76 μm, 主要成分为聚乙烯、聚丙烯和半合成纤维素等。近年来相关研究和分析发现[12-16],金图企鹅体内的微塑料可能来自3 种途径: (1)一些颜色鲜艳的塑料碎片被误认为食物直接摄食;(2)通过饵料生物间接摄食; (3)通过受污染的水或沉积物偶然摄入, 已有研究证明南极区域的水体和沉积物中存在微塑料[13-16]。
王企鹅(Aptenodytespatagonicus)在整个南大洋的亚南极岛屿繁殖, 是最重要的鸟类消费者之一[50], 它们能下潜到400 m 的深度, 并主要以中上层鱼类为食[51]。Le Guen 等[51]通过研究2017年在南乔治亚采集的3 种不同生活史的王企鹅粪便(育雏期、孵化期、未繁殖期), 发现其中84.7%为纤维素, 3%为动物绒毛, 12.3%为人工合成纤维,所呈现的都是纤维状的微塑料污染物, 并且孵化期王企鹅粪便中微纤维量比育雏期王企鹅高, 推测可能是育雏期王企鹅反刍食物给幼鸟从而导致自身微纤维含量低, 或是由于孵化期王企鹅的觅食行程比育雏期更长, 更容易接触到微纤维。
Fragão 等[52]通过研究南极半岛和斯科舍海域的帽带企鹅(Pygoscelisantarcticus)、阿德利企鹅(Pygoscelisadeliae)和金图企鹅的粪便, 在317 个粪便样品中发现了92 个人造塑料颗粒, 在所有提取的颗粒中有35%被鉴定为微塑料, 主要是聚乙烯和聚酯合成物。Panasiuk 等[53]通过研究乔治王岛上的阿德利企鹅的胃含物, 发现其中99.9%为南极大磷虾, 也发现了一些粒径大于1 cm 的塑料碎片, 其形状扁平疑似渔网碎片, 颜色多样, 这意味着有不同的来源, 但对塑料成分并未深究。总之, 不同物种企鹅的觅食策略不同, 比如非洲企鹅(Spheniscusdemersus)捕食上层鱼类, 王企鹅捕食中层鱼类, 金图企鹅在靠近海床的地方觅食。通过研究这些企鹅, 可以确定哪种觅食策略受微塑料污染的影响最大, 这反过来又有助于确定未来微塑料污染风险相对较高和较低的物种。
值得注意的是, 在所有企鹅的粪便研究中,纤维状的微塑料污染物占了很大的比重[45,51-52]。有研究表明, 到目前为止在海洋表层水中最丰富的微塑料是微纤维[54], 微纤维通常被认为是由合成材料制成, 如聚酯或聚酰胺(Polyamide, PA)等。此外, 在海洋中也发现了一些自然纤维如羊毛和棉花等。De Falco 等[55]发现, 5 kg 的聚酯纤维纺织物的洗涤废水能释放超过600 万个微纤维。Gröndahl 等[56]调查发现在南极洲的71 个考察站中, 约有52%没有废水处理系统。假设每个人在南极考察站每周洗1 次衣服或被褥, 每周清洗3~11 件人工合成织物, 那将会有介于5~255 亿个的塑料纤维释放到南大洋, 假设90%的纤维在废水处理过程中被去除, 那么在10 年内释放到南大洋的塑料纤维将达到250 亿个[57]。洗衣废水中释放的微纤维可能是重要的微塑料污染源。现阶段关于在南极的水体或沉积物中微纤维的报告很少,尽管在废水排放源附近可能会较容易检测到微纤维, 但是由于稀释效应, 在开阔海域中检测会更加困难。据统计, 2017 年, 全球共生产了100 万吨纤维产品, 其中人工合成纤维占70%[51], 且纺织纤维生产数量逐年增加, 但目前没有建立对污染废水排放入海的全球监管体系, 故迫切需要在南极海洋生态系统中, 监测和评估自然和人工合成的微纤维的赋存特征和生态风险。
1.4 海洋哺乳动物
海洋哺乳动物尤其是鳍脚类动物, 它们能直接摄食含有微塑料的整个猎物, 故其摄入微塑料往往与猎物本身有关。1991 年和1997 年, Eriksson和Burton[58]在麦夸里岛收集了共计145 份南极毛皮海狮的粪便, 共检测出164 个塑料碎片, 均小于10 mm, 且与在海滩上发现的塑料漂浮物成分一致, 这些漂浮物的本体被冲上岸并在海岸线上分解后再次流入海里, 其中一些颗粒被鱼类摄食,这些鱼类后来被海豹摄食, 从而导致塑料碎片的生物积累。1999 年, McMahon 等[59]在麦夸里岛上收集了51 份新西兰海狮(Phocarctoshookeri)的粪便, 其中塑料碎片出现率为 11.5%, 粒径约为1 mm, 且有塑料出现的粪便中都检测出了大眼电灯鱼的耳石。1990—2001 年间, van den Hoff 等[60]在麦夸里岛的海豹繁殖地共收集了332 份幅北毛皮海狮(Arctocephalustropicalis)粪便, 检测出47个塑料碎片, 平均粒径为4.1 mm, 且粪便中存在大量的大眼电灯鱼(Electronasubaspera)耳石。研究发现, 处于繁殖期的成年海豹在为幼崽提供食物时, 会进行短距离觅食, 而以灯笼鱼科为首的中上层鱼类则是它们主要摄食对象, 约95%的海豹会在夜间潜至10 m 深水处觅食, 而灯笼鱼会在夜间垂直迁移至上层水域摄食浮游生物[60]。但是目前没有相关研究能够证明灯笼鱼体内含有微塑料, 具体情况有待后续的进一步研究。
低营养级生物对微塑料的摄取可能是微塑料进入海洋食物链的一个潜在途径。许多中上层鱼类存在昼夜垂直迁移的习性[61], 它们白天停留在较深的水层以躲避捕食者, 夜间迁徙至上层捕食浮游动物, 这种迁移特性可能导致微塑料从表层转移到海洋深处。高营养级消费者通过捕食低营养级消费者而生存, 低营养级消费者摄入微塑料造成的诸如堵塞或损害消化道, 虚假的食物满足感, 抑或是有毒化合物的转移等[61], 这些因素会直接影响它们的种群数量动态, 进而影响到高级消费者, 使得食物链生态失衡, 进而影响南极原本就简单的生态系统。
微塑料在鲸类动物中的研究目前十分匮乏,主要受限于伦理道德、法律和物流运输方面的限制, 现有的少数研究都是依靠机会性的接触已死亡的搁浅鲸鱼[62-63]。有关南极和南大洋的鲸类微塑料研究有待进一步开展。
2 南极各类营养级生物微塑料污染程度与其他区域比较
2.1 南极无脊椎动物微塑料污染程度与其他地区比较
为了更好地展现微塑料在南极生物体内污染程度, 本文将来自各大洋和近岸已发布的各类营养级生物摄入微塑料资料进行比较分析。
从全球范围来看, 南极无脊椎动物体内微塑料污染程度呈中等水平(表2), 无脊椎动物会直接摄入环境中存在的微塑料, 不同区域无脊椎动物体内微塑料丰度存在差异或许与摄食方式和栖息地环境不同有关。肉食性动物、草食性动物和滤食性动物体内微塑料丰度均存在一定差异, 沉积物中的微塑料往往也会影响一些底栖无脊椎动物。例如, Hennicke 等[64]在调查区域的沉积物中发现微塑料平均丰度范围为70~430 个·kg-1, 且沉积物中出现的微塑料颜色类型与生物样中一致,人口稠密地区的沉积物污染程度比偏远地区更严重。此外, 南极无脊椎动物体内出现的微塑料主要类型为聚酰胺和聚乙烯, 与其他地区报道一致,这些聚合物常用于渔具、绳索、食品包装和服装材料, 易通过陆地和海洋的人为活动输入海洋。
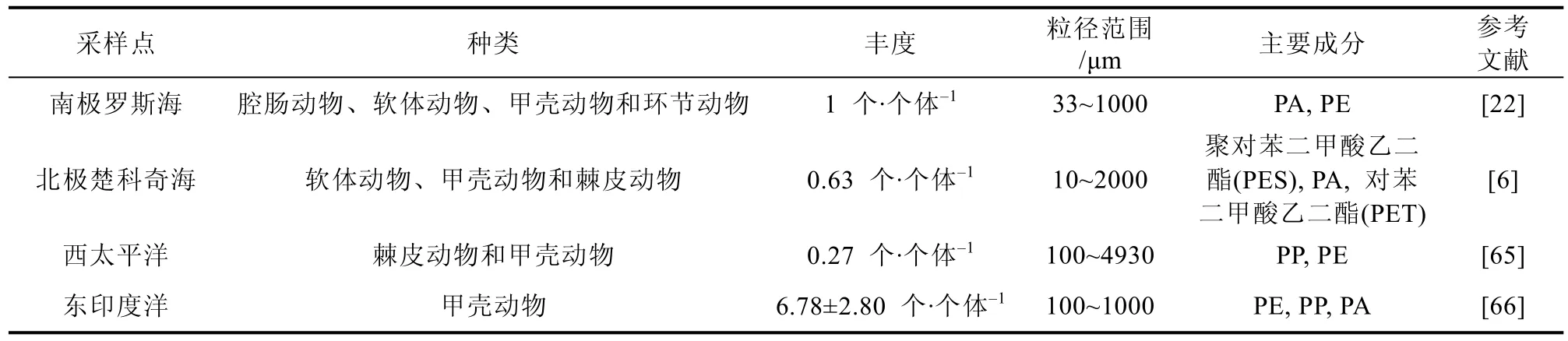
表2 不同地区无脊椎动物体内微塑料丰度Table 2. Microplastic abundances in invertebrates from different regions
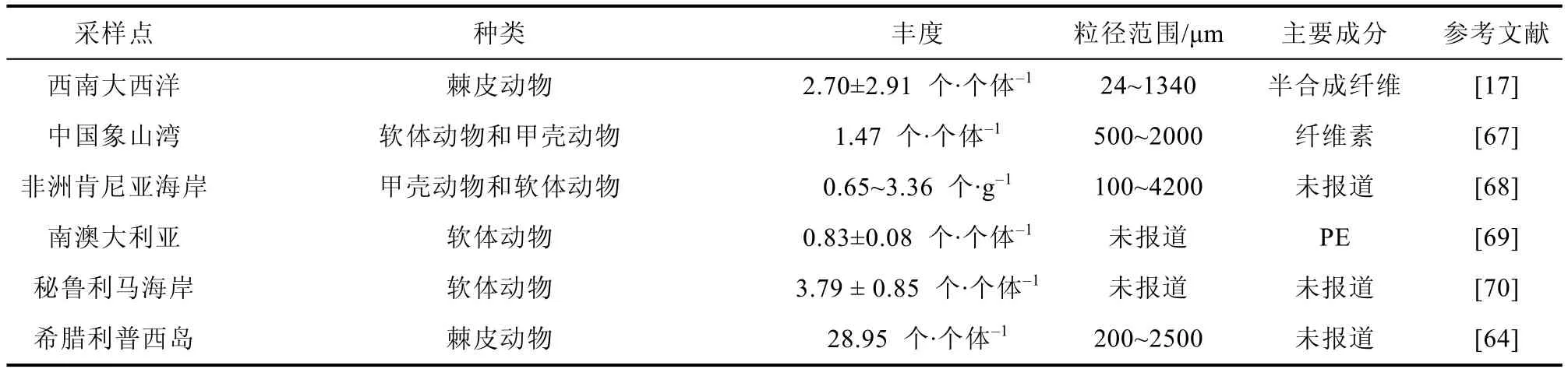
续表
2.2 南极鱼类微塑料污染程度与其他地区比较
南极鱼类体内微塑料污染程度在全球范围内处于中等水平(表3), 不同区域鱼类体内微塑料丰度差异或与栖息深度和环境有关。南极鱼类主要是底栖鱼类, 底栖鱼类是底栖生物的主要消费者,也以浮游动物为食[71], 可能会间接摄入来自猎物中的微塑料。南极罗斯海以及阿蒙森海存在频繁的科学调查活动, 废水处理厂、航运和洋流等都会促进微塑料的释放和传播, 这也一定程度上加剧南极鱼类对微塑料摄入量[42]。此外, 南极鱼类体内微塑料主要成分来自纺织工业, 包装和渔具, 这与大洋和近岸的报道一致[10,72-77], 这表明海洋鱼类体内存在的微塑料与人类活动息息相关。
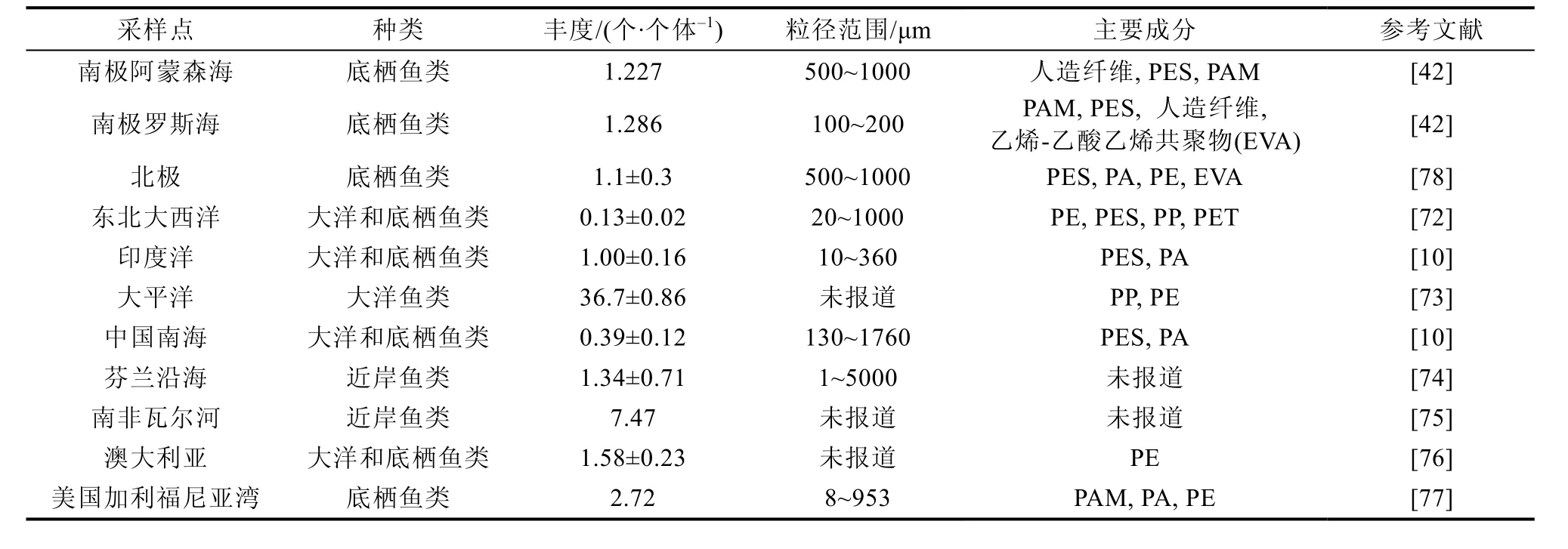
表3 不同地区鱼类体内微塑料丰度Table 3. Microplastic abundances in fish from different regions
2.3 南极鸟类微塑料污染程度与其他地区比较
南极鸟类体内微塑料丰度在全球范围内处于中等水平(表4)。因其独特的生理结构, 鸟类摄入的微塑料会暂时储存在砂囊和前胃中, 一段时间后才能通过肠道和泄殖腔排出体外[79], 这或许能解释粪便样中的微塑料丰度普遍低于胃肠道中微塑料丰度。南极企鹅粪便中的微塑料丰度较其他海鸟较低, 可能是企鹅的栖息地较为固定且移动范围较短, 而多数海鸟具有迁徙的习性, 能接触更多微塑料。南极企鹅粪便中涉及的微塑料主要成分来自纺织工业与渔业, 也出现在当地沉积物、海水中[13,57], 和全球其他区域报道一致。
2.4 南极海洋哺乳动物微塑料污染程度与其他地区比较
现有的研究表明, 全球范围内的海洋哺乳动物都存在不同程度的微塑料污染(表5), 其中鲸豚类体内微塑料丰度较为显著, Moore 等[62]在白鲸(Delphinapterusleucas)肠道中没有发现任何中大型塑料碎片, 推测白鲸并没有直接或者刻意去摄入塑料, 体内存在的微塑料更可能是来自于猎物。南极海洋哺乳动物体内微塑料污染程度在全球范围内处于低水平, 南极海豹粪便中发现的聚合物种类与调查区域海上漂浮物成分相同[58], 表明海豹可能吞食来自海上的微塑料, 或通过猎物间接摄入, 其主要成分聚乙烯和聚丙烯主要来自纺织工业与渔业, 也在全球众多鱼类体内发现,与其他地区报道一致, 表明微塑料很可能通过食物链进行传递。

表5 不同地区海洋哺乳类体内微塑料丰度Table5. microplastic abundances in marine mammals from different regions
3 总结与展望
关于南极生物摄入微塑料的研究表明, 从低营养级的无脊椎动物到高营养级的海洋哺乳动物都存在微塑料富集现象, 其中南极无脊椎动物、鱼类和鸟类体内微塑料污染程度在全球范围内呈中等水平, 南极海洋哺乳类动物体内微塑料污染程度普遍低于其他区域。低营养级生物摄入微塑料可能是微塑料进入海洋食物链的潜在途径之一,像南极大磷虾以及灯笼鱼等关键物种是否摄入微塑料还有待学者进一步探明。此外, 现阶段的研究多为描述性研究, 更多深入而全面的评估有待今后持续开展跟踪监测、测试分析和研究, 而且也应该更加关注微塑料的长期潜在影响。
猜你喜欢
海洋信息技术与应用(2022年1期)2022-06-05
小哥白尼(趣味科学)(2020年12期)2021-01-18
小哥白尼(神奇星球)(2020年12期)2021-01-18
儿童时代·幸福宝宝(2020年9期)2020-09-08
小天使·二年级语数英综合(2019年9期)2019-11-08
学生天地(2019年32期)2019-08-25
红领巾·探索(2018年12期)2018-01-26
阅读与作文(小学高年级版)(2016年12期)2016-12-27
金色少年(奇趣科普)(2016年8期)2016-09-21
英语学习·阳光英语(2013年3期)2013-07-30
