
邛海大型水生植物时空分布及其退化原因*
2024-01-13 07:40董艳珍张培育李小艳郑丙辉
湖泊科学 2024年1期
董艳珍,张培育,李小艳,郑丙辉
(1:西昌学院,四川高原湿地生态与环保应用技术实验室,西昌 615013) (2:中国科学院水生生物研究所,武汉 430072) (3:中国环境科学研究院,湖泊水污染治理与生态修复技术国家工程实验室,北京 100012)
水生植物是湖泊水生态系统的重要组成部分[1]。水生植物能够固着和稳定沉积物、降低悬浮颗粒物浓度、促进水体中磷的沉降和减少沉积物中磷的释放、滞留与削减污染物、释放氧气、抑制浮游植物生长[2-5]。影响沉水植物生长和分布的主要因子有营养盐、光照强度、底质、悬浮物、水流、温度和pH等[6]。邛海是云贵高原11个面积大于25 km2的天然湖泊之一,是四川省第二大天然湖泊。邛海是西昌市的饮用水水源地,也是邛海泸山国家级风景名胜区的核心区,承担着流域水资源供给、下游安宁河生态流量保障、局地气候调节和生物多样性维护的功能。30多年来,围绕邛海环境问题研究主要集中在水污染和富营养化等方面,如时瑶等研究了2020年邛海氮磷时空分布特征及其生态影响[7],许宵宵等开展了2003-2014年邛海水体富营养化评价[8],孙宏亮等开展了2013年邛海水生生物多样性调查及富营养状态评价[9],周申立等开展了1998-2004年邛海水生生态系统健康评价研究[10],姚维志等开展了1993年邛海浮游植物与水质污染及富营养化研究[11]。1980s后期,开始有邛海大型水生植物调查研究报道。国家“七五”科技攻关项目“中国典型湖泊富营养化调查”成果对邛海环境状况做了较全面的调查[12]。王治农介绍了1990s邛海水生植物分布情况[13]。2002年,云南省环境科学研究院开展了邛海水生植物调查,取得了一组比较完整的水生植物分布数据[14]。2009年,张宇等报道了邛海及附近湿地水生维管植物种类组成[15]。李小艳等在2017年4月和2018年4月调查了邛海浅水湖带及其附近人工湿地中沉水植物,报道了沉水植物的种类及出现频度[16]。自1960s、1970s起,周边被填湖造田、围湖造塘、网箱养鱼,近2/3的滩涂、苇塘遭到破坏和不合理开发利用,邛海面积由34 km2降至26.7 km2,湿地生态遭到严重破坏。竺美等完成了邛海生态环境保护总体方案,初步阐明邛海生态环境问题及保护对策[17]。近年来,西昌市实施了“三退三还”、湿地恢复与建设等工程,邛海湿地面积大幅增加,但是邛海水生植被没有出现恢复迹象,甚至在2022年还出现春季束丝藻水华和秋季鱼腥藻水华,水生态系统健康问题令人担忧。本文基于2021-2022年邛海水生植物系统调查结果,初步阐明邛海大型水生植物现状,结合文献资料,分析其变化趋势,阐明邛海沉水植被退化的原因,提出水生植被恢复的对策建议。
1 研究区域及数据来源
1.1 研究区概况
邛海流域(27°47′~28°01′N,102°07′~102°23′E)属于长江流域,雅砻江水系,安宁河的上游,邛海位于西昌城区西南面。邛海流域面积307.67 km2。邛海系公元前116年因一次地震沉陷而成的淡水湖泊,湖面呈L型。2003年实测结果南北最长为10.3 km,东西最宽为5.6 km。邛海水下地形周边坡度变化较大,水底平缓。东北方向地形较为复杂。2003年实测时湖面水位高为1509.82 m,湖岸线长约37.4 km,面积约为27.408 km2,库容量为2.93×108m3,最大水深为18.32 m,平均水深为10.95 m[18]。邛海流域气候属亚热带高原季风气候,冬暖夏凉、干湿分明,年平均气温为20℃,年降水量为1000 mm,多年平均湖面降水量2650×104m3,湖泊补给系数9.97,湖水滞留时间约834 d[14]。其中,官坝河、高仓河和鹅掌河为主要入湖河流,其河口冲积扇成为水生植物生长的主要区域[19-20]。在邛海西岸北端的海河为邛海唯一出水口,湖水排入安宁河。
1.2 数据来源与调查方法
2021年7月和12月在沿岸和敞水区分别布设15和3个点位开展定点观测(图1)。水生植物的调查方法参考洱海相关调查方法[21]。每个样点至少设置3个样方,由于邛海大部分区域的水生植物分布在距离岸边20 m内和水深小于2 m的区域,调查过程中采样点布设并不是完全遵循垂直于岸线方向,而是在有水生植物分布区域随机布设。估测每个样方内水生植物的盖度,使用抓钩采草器(面积为0.05 m2)采集整株水生植物,现场区分种类,称量植物鲜重。水生植物的高度通过卷尺随机测量5株植物的株高,计算出该物种的平均株高。物种鉴定参考《中国植物志》[22]。2022年5月和9月沿湖岸线开展巡航观测,探寻水生植物分布边界以及最大分布水深,使用星瑞源资源宝软件勾画水生植物分布图斑,估算水生植物分布面积。

图1 邛海水生植物调查布点Fig.1 The aquatic plant survey sites of Lake Qionghai
2 结果与分析
2.1 水生植物调查结果
2.1.1 水生植物生境特征与生物量 2021年7和12月调查的各点位主要生境要素和水生植物分布见附表Ⅰ。沉水植物的空间分布主要取决于水域的生境因子[23],基底则是水生植物生长的最关键因子。邛海近岸的基底特征控制着水生植被的种群结构分布:
北岸水域:包括6、7、10和11号点。水深0.95~2.1 m,透明度0.95~1.5 m,以淤泥基底为主,是目前邛海水生植物分布最丰富的区域,分布有挺水植物、浮叶植物和沉水植物,水生植物分布宽度为50~120 m。
高枧湾水域:包括12、13、14、15和16号点。其中12、13、14和15号点位于高枧湾东北岸,水深0.65~1.5 m,透明度0.3~0.6 m,淤泥基底,挺水植物和浮叶植物分布较多,沉水植物极少;水生植物分布宽度为10~200 m。16号点位于高枧湾西岸邛海公园附近,水深1.5 m,透明度1.0 m,人工种植挺水植物莲、菰和浮叶植物荇菜,莲的分布宽度约为500 m。
西岸水域:包括1和17号点。水深1.8~2.0 m,透明度1.5~1.8 m,石砾(抛石人工堤岸)基底,1号点有少量水生植物分布,植物分布宽度不足10 m,17号点无水生植物分布。
南岸水域:包括2、3和4号点。水深1.2~1.9 m,清澈见底,2、3号点为石砾和砂质底,分布有少量水生植物,分布宽度为12~20 m;4号点(核桃村湾区)为淤泥基底,分布有大量的莲,分布宽度约为100 m。
东岸水域:5号点,水深2.1 m,清澈见底,基岩基底,坡度30°以上,仅在沟坎处有少量水生植物,分布宽度约为15 m。
开敞水域:包括8、9和18号点。水深大于9 m,而透明度仅为2~3 m,水下光环境条件不适于水生植物生长。
3-4月邛海水生植物处于萌发阶段,生物量较低,7月水生植物生长旺盛。2021年7和12月各测点生物量如图2所示,沉水植物的高度为20~135 cm,生物量为0.15~3.4 kg/m2,优势种群为穗状狐尾藻、大茨藻;浮叶植物高度与所在水域的水深一致,一般为100~180 cm,生物量为2.0~4.5 kg/m2,优势种群为荇菜和菱;挺水植物莲分布的水域水深为100~150 cm,莲的生物量为0.5~3.3 kg/m2,在海河口南侧、月亮湾底和核桃村3个水域成为优势种。12月随着水生植物枯萎和鸟类啄食,水生植物生物量明显下降,沉水植物的生物量为0.1~2.9 kg/m2,高度为25~100 cm,优势种群为苦草、穗状狐尾藻;浮叶植物生物量为0.04~6.8 kg/m2, 高度与水深一致,为100~180 cm,优势种群为荇菜。

图2 2021年7月和12月各采样点的水生植物生物量Fig.2 The biomass of aquatic plants at each sampling site in July and December 2021
2.1.2 水生植物种类 根据2021-2022年调查,共记录邛海水生植物23种(表1),其中沉水植物8种,占比34.78%;浮叶植物3种,占比13.04%;漂浮植物5种,占比21.74%;挺水植物7种,占比30.43%。除2021年12月水生植物种类较少外(13种),其余3个季节水生植物种类相差不大。
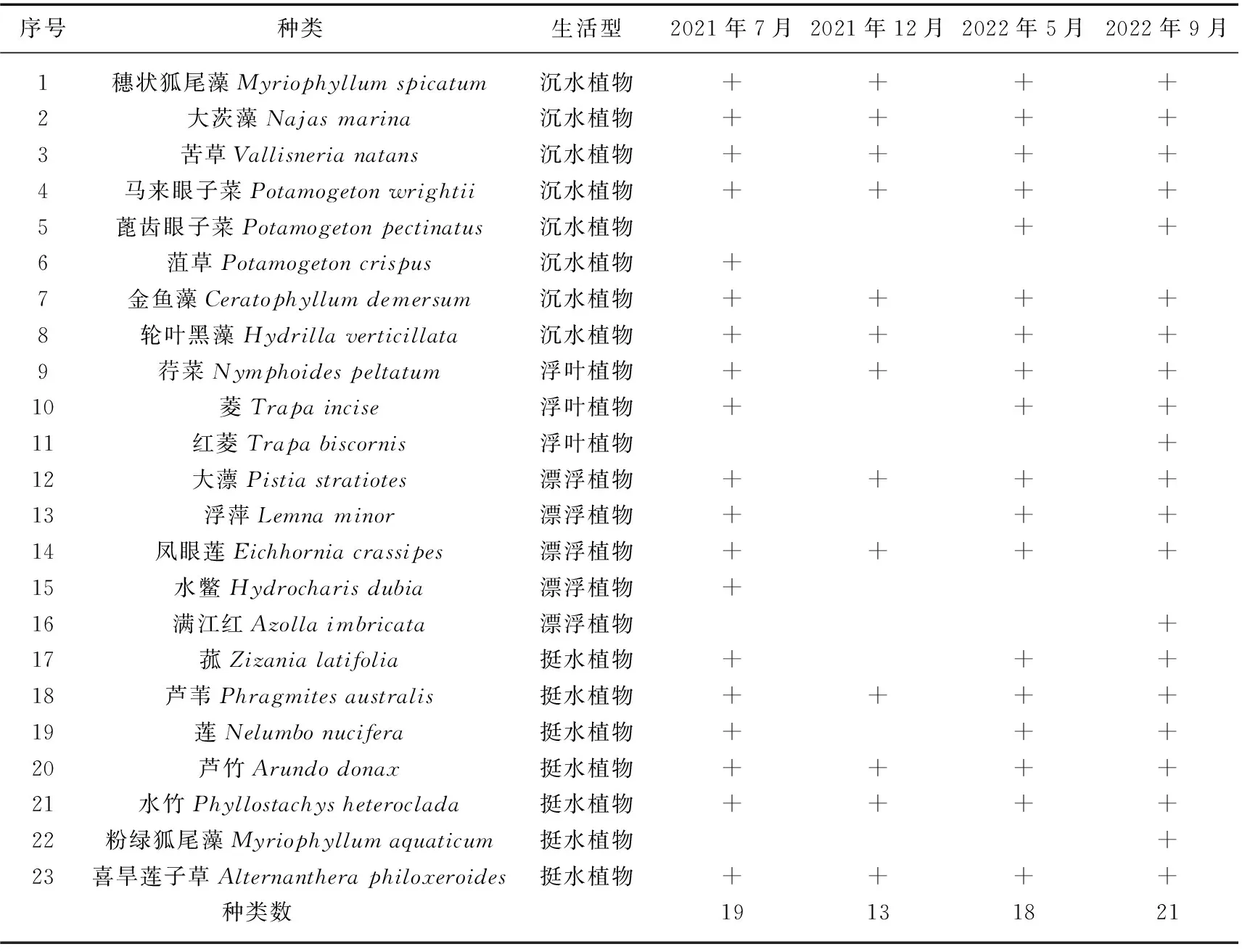
表1 邛海水生植物名录*Tab.1 Aquatic plants in Lake Qionghai
2.1.3 主要水生植物群落分布特征 邛海水生植物主要有9种群落,包括苦草、穗状狐尾藻、金鱼藻和大茨藻4种沉水植物群落,荇菜和菱两种浮叶植物群落,莲、菰和芦苇3种挺水植物群落(表2)。邛海主要沉水植物种类与2017-2018年调查相似[16],但在2021-2022年的4次调查中均未发现穿叶眼子菜(Potamogetonperfoliatus)。

表2 邛海主要水生植物群落及特征Tab.2 Main aquatic plant communities and characteristics of Lake Qionghai
2.2 邛海水生植物空间分布
2022年5月和9月沿湖岸线巡航观测以及2021年定点监测结果,邛海水生植物空间分布如图3所示。邛海水生植物主要分布在北部小渔村附近沿岸带(距岸200 m有水生植物分布)和西岸海门桥水域(距离岸边300 m 左右),其余主要分布于邛海各湖湾及湖岸水深小于2 m的地方。由于邛海整体水深较大,适合水生植物生长的区域不多。水生植物分布总面积约为1.3 km2。相比夏季,冬季植物分布范围缩小。空间分布特征与2017-2018年调查结果相近[16]。
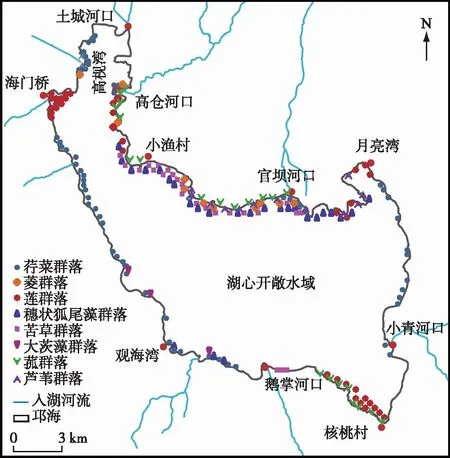
图3 邛海水生植物分布Fig.3 Distribution of aquatic plants in Lake Qionghai
(1)北片区从月亮湾到土城河口全长约11 km的湖岸,其中月亮湾到高仓河段是邛海沉水植物分布的主要区域,沉水植物最大分布宽度约300 m,面积约0.8 km2。苦草群落在此区成片分布,占此区水草面积的1/2左右。穗状狐尾藻群落在此区呈斑块状分布,冬春季节常为苦草群落的伴生种,夏秋季节则成为优势种。荇菜群落和菱群落在此区均呈斑块状分布,主要分布在湖湾和河口处。湖岸主要分布有芦苇群落和菰群落。而高仓河口至土城河口段除岸边有少量挺水植物和浮叶植物分布外,浅水区无沉水植物分布。
(2)西片区从土城河口到观海湾约9 km的湖岸,水草分布面积约0.25 km2,其中土城河口到海门桥段是莲群落和荇菜群落集中分布的区域,海门桥到观海湾段荇菜群落和穗状狐尾藻群落呈斑块状分布,斑块面积小。
(3)南片区从观海湾到核桃村全长约6 km的湖岸,水草分布面积约0.15 km2。莲集中分布在核桃村附近,形成2个大的群落,面积超过此区水生植物面积的1/2;大茨藻、荇菜、穗状狐尾藻、苦草和菱群落在此区零星分布。
(4)东片区从核桃村到月亮湾全长约7 km的湖岸。此片区水生植物较少,零星分布有穗状狐尾藻群落、苦草群落和荇菜群落,总面积不足0.1 km2。
2.3 邛海水生植物演变特征
与历史资料对比发现,1990s前到2002年邛海水生植物种类减少明显,2002-2022年种类总体变化不大,但群落结构及空间分布上均发生较大变化(表3),如水生植物分布面积大幅度缩减,分布水深变浅,沉水植物群落结构简单化,分布范围变窄,浮叶植物与挺水植物分布范围扩张等。

表3 1988-2022年邛海水生植物演变情况Tab.3 Evolution of aquatic plants in Lake Qionghai from 1988 to 2022
1980s后期到1990s初,邛海水生植物面积占湖面积的20%左右[12-13],到2002年水生植物分布水深不超过3 m,面积约2.8 km2,约为湖泊面积的10%[14];2022年邛海水生植物分布水深不超过2 m,面积约1.3 km2,约为湖面积的5%。
1980s后期到1990s初,大型维管束植物有40余种,苦草、菹草、马来眼子菜、篦齿眼子菜、微齿眼子菜、金鱼藻、轮叶黑藻和大茨藻8种沉水植物是邛海水生植物的常见优势种,分布广泛。其数量少于高原湖泊的洱海,洱海挺水植物15 种、浮叶和漂浮植物各7 种、沉水植物26 种[24]。2003年邛海沉水植物群落有6种,眼子菜科植物菹草和微齿眼子菜未形成群落。到2021-2022年,只剩4种沉水植物群落,眼子菜科植物中的菹草、微齿眼子菜、篦齿眼子菜、马来眼子菜以及轮叶黑藻均未能形成群落,仅在局部区域作为狐尾藻群落的伴生种出现。而目前在邛海湖区广泛分布,成为沉水植物优种之一的穗状狐尾藻在1990s却并非水生植物优势种。
浮叶植物和挺水植物的分布范围扩张。1990s荇菜不是邛海水生植物的优势种;2003年邛海南岸和西岸均未出现荇菜群落,北岸也只在湖湾少量分布,但到2021-2022年,几乎全湖湖湾均有分布,面积约占水生植物总面积的20%。除原来分布的核桃村,2022年莲从月亮湾到观海石湖滨带、朱家河湖滨带、核桃村湖滨带均有增加或扩展分布。
可见,邛海水生植被面积缩减十分严重,沉水植物群落单一化,眼子菜科的沉水植物群落逐渐减少,穗状狐尾藻群落成为优势,近年来浮叶植物荇菜和挺水植物莲扩张显著。2000年前后,是邛海水生植物分布面积、种群结构产生突变的关键时期。
3 讨论
大量研究证实,湖泊中水生植物分布及生物量一方面与湖泊中物理、化学生境条件直接相关,如:水体透明度、湖泊水深、风浪强度、营养盐水平、底质理化特性等均会对水生植物的繁殖、生长产生影响。另一方面,也与湖泊中生物要素有关联,如:蓝藻水华、附着生物、鱼类和底栖生物等均会对沉水植物的生长、繁殖产生影响[1]。总体上看,邛海水生植物多样性偏低,会导致水生动物的多样性降低,这样形成的食物网结构过于简单,不利于水体生态系统的物质循环和能量流动,且水生植物群落抗干扰的能力比较差,在富营养化及其他人类活动胁迫下,水生植物容易衰退,从而降低生态系统的服务功能[25-26]。邛海水生植被退化主要有两大原因,一是1998年洪水导致邛海水生植被遭受到突变性破坏,二是邛海关键生境改变导致水生植被渐变性退化。
3.1 水生植被面积缩减的突变性原因
3.1.1 1998年邛海洪水事件 据邛海渔民介绍,历史上高枧湾水域是水生植物分布较多的水域。1988-1989年与2002年两期调查结果表明,邛海水生植被面积和种类存在显著变化,也可以说是一种突变。该突变发生在1998年,原因是1998年6月28日-7月2日,西昌、德昌、喜德三县市降雨量超过160 mm,安宁河水位陡涨[27],邛海水位由1998年6月29日的1507.52 m上升到7月3日的1508.08 m,涨幅高达1.16 m。邛海下游的东河高泥沙洪水从海河倒灌,叠加干沟河的高泥沙洪水进入邛海高枧湾,使得高枧湾出现大面积泥沙淤积,导致邛海高枧湾数千亩水生植物死亡,截至2022年海门桥至小渔村沿岸水生植物仅有少量穗状狐尾藻,植株叶片被浮泥覆盖,生长情况较差。
3.1.2 洪水之后水生植被退化加剧 大量沉积物再悬浮,降低了水体的透明度及水下光照强度;另外,再悬浮的颗粒物可以附着于沉水植物的表面,影响沉水植物的光合作用、生长与植被发育。浅水湖泊中水生植物的分布可能直接与风浪的大小有关,大的风浪可导致湖泊中的沉水植物消失。风浪除了可以直接损伤水生植物外,所引起的沉积物再悬浮是限制湖泊中沉水植物分布的另一个原因。太湖风浪与透明度观测和模拟结果表明,浅水湖泊表层悬浮物浓度和透明度具有很好的相关性,不同水深处的悬浮物浓度与风速的变化趋势大致相近,风速越大,悬浮物浓度越大[28-29]。1998年的洪水过程带来的大量粉质黏土在高枧湾沉积,使得高枧湾北部、中部底泥以粒径相对较小的粉质黏土为主。高枧湾底泥中粒径为0.005~0.075 mm的粉粒,约占70%;还有粒径小于0.005 mm的黏粒,约占20%,这两种组分占比达到90%左右,两种底泥颗粒易悬浮,均不易沉降。高枧湾是山谷风的汇集处,风浪较大。在风速为15 m/s时,邛海最大浪高可达1.5 m,高枧湾大部分区域水深为1.5 m,风浪基本可以影响湖底,造成湖底泥沙再悬浮;沉水植被的退化又降低了底泥的固化能力,同样的扰动条件下底泥再悬浮加剧,进一步恶化了水下光环境,形成恶性循环。因此,1998年洪水事件既是邛海水生植被退化突变点,也是进入水生态退化恶性循环的起点。
3.2 水生植被面积缩减的渐变性原因
3.2.1 运行水位变化 湖泊水位变化对水生植物生长有显著的影响[30]。浅水湖泊中伴随着水位的上升,水体透明度与水深的比值逐渐降低,造成湖泊底部的光照强度显著下降,限制了沉水植物的生长发育[31]。2002年海河闸建成运行,改变了邛海水位节律。海河闸建成前(1991-2001年)和建成后(2009-2021年)邛海水位统计表明,建闸后邛海年平均水位为1510.29 m(范围为1510.19~1510.36 m),比建闸前(范围为1509.79~1510.23 m)升高了0.30 m;建闸后邛海年最大变幅为0.61 m(范围为0.39~1.50 m),比建闸前(范围为0.73~2.24 m)减少了0.62 m。特别是水生植物萌发的4-5月,连续15日、连续30日和连续45日最低平均水位分别升高0.47、0.44和0.42 m(表4)。
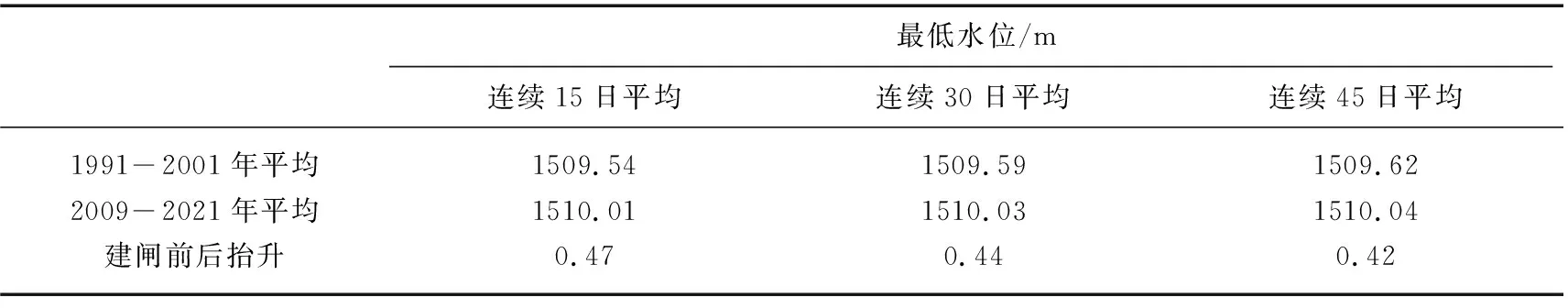
表4 海河建闸前后邛海最低水位Tab.4 The lowest water level of Lake Qionghai before and after gate construction of Haihe River
由于邛海岸线已经固定,受近岸水下地形坡度的影响,水位抬升造成适宜水生植物生长的空间减少,如小渔村岸段,1980s、1990s该岸段水生植被宽度约为400 m,2021-2022年调查,除莲外,其他水生植被最大宽度仅为200 m左右。因此,海河闸建成运行对邛海水生植物生长有着直接的影响。
随水深增加,苦草生长速率下降,生物量减少[32-33]。湖泊运行水位抬升,适宜沉水植物生长的水域范围进一步缩减,水生植物和草型生态系统退化加速[34]。
1954年巢湖建闸以前,巢湖水生植物生长茂盛,植被发育良好,水生植被面积多达200 km2,约占该湖泊面积的1/4。1962年巢湖闸建成运行,由于建闸运行水位抬升、变幅减少以及养殖和污染等因素的影响,巢湖的水生植物在数量和种类上都迅速减少。1981-1983年调查巢湖水草覆盖面积仅占湖泊面积的2.54%[35]。
3.2.2 透明度降低 王洪铸等在2001 年12 月-2003 年3 月对长江流域4个湖泊区域的大型水生植被进行研究,指出在平均水体深度、塞克盘深度(透明度盘深度)、温度、总氮、总磷和叶绿素a等因素中,透明度与水生植被生长的平均水体深度比是影响植被生物量的关键因素[36]。1986-1991年连续监测结果表明,邛海水体透明度为2.5~5.3 m[37],根据1994-2000年监测数据,1995年后邛海水质出现明显下降趋势,1999年透明度下降到历年最低,不足1.0 m[38]。东部平原典型湖泊水体透明度与悬浮颗粒物浓度之间具有明显的相关性,真光层深度与透明度的关系为:Zeu=2.46SD+0.11[39]。以高枧湾水域最为明显,底泥再悬浮造成水下光环境恶化,水体透明度仅为0.3~0.5 m,最好时段也不超过1.0 m,高枧湾真光层深度仅为0.85~1.34 m,而水深在1.5~1.8 m左右,透明度与水深比值仅为0.3~0.4。因此,高枧湾水域光环境难以满足沉水植物生长的要求,这是高枧湾沉水植物匮乏的原因。同样,太湖透明度降低叠加水位明显上升,造成透明度与水位(水深)的比值显著降低,特别是草型湖区1998-2019年透明度与水位(水深)的比值降低了43.5%,导致沉水植被生长发育被极大地限制[40]。
3.2.3 其他因素 2020年,邛海总体水质良好,处于中营养状态,叶绿素a平均浓度约为10 μg/L,最高浓度出现在高枧湾,主湖区没有出现明显的蓝藻水华现象[41],因此对大型水生植物繁育、生长的影响较小。一般认为生物入侵对水生系统的影响要大于对陆生生态系统的影响,入侵种的泛滥不仅挤压了本地物种的生存空间,而且漂浮植物还会对沉水植物生长空间形成极大的胁迫,容易导致沉水植物的衰退[25]。调查发现至少4种入侵水生植物(凤眼莲、喜旱莲子草、大薸和粉绿狐尾藻),主要分布在湖滨湿地区域,因此目前水生植物入侵现象对邛海的水生植被影响有限。草食性鱼类大量放养是我国很多湖泊水生植被减少的主要原因之一,并由此导致草型湖泊向藻型湖泊演替,加速水体富营养化进程[42]。底栖型鲤科鱼类等的放养会促使底泥频繁地再悬浮,导致水体透明度下降以及内源性营养盐释放,引起水生植物退化。而孔德平等研究表明,2013和2015年实际采集到邛海湖体鱼类共13种,太湖新银鱼、鲢、鳙、鲫、鲤、条、红鳍原鲌、草鱼、黄颡鱼等经济鱼类以及棒花鱼、麦穗鱼等小型鱼类是邛海主要的鱼类组成,邛海底栖型和食草性鱼类并没有显著增加[43],因此排除了这一可能。
3.3 邛海水生态修复的建议
综上分析,水生植物退化的因素多样,关键是水生植物生境条件改变[1,6]。建议调整邛海运行水位的法律规定,尽可能恢复水位运行的自然节律,特别是降低植物萌发季节水位。其次,增加水体透明度,具体要满足:真光层深度与水深比不低于1,没有水华蓝藻的频繁侵入,从而促进水生植物生长和草型生态系统的发育[34,44]。建议加强西昌东部平坝区城镇生活污水收集处理,减少入湖负荷;加强高枧湾上游流域森林植被恢复,减少泥沙输入。再次,强化已建湖滨湿地生态工程的管理,发挥其生态效益;采取人工干预措施,恢复湖区挺水植物、浮叶植物和沉水植物,优化食物网结构,逐步修复邛海水生态系统[45]。总之,水生态修复要坚持以自然恢复为主、人为干预为辅的原则,化整为零,分区分片修复。
4 结论
1)邛海湖区有水生植物23种,分布在水深2 m内,总面积约1.3 km2。邛海水生植物主要分布在湖泊北面月亮湾到高仓河口一带,其面积约占水草分布总面积的60%,湖泊西面和南面水生植物呈斑块状分布,东面水生植物较少,呈零星分布。
2)从1990s到现在的30年间,邛海水生植物分布由湖面积的20%左右缩减到不足湖面积的5%;沉水植物群落结构呈现出单一化,穗状狐尾藻群落逐渐取代了眼子菜科沉水植物群落;荇菜等浮叶植物和莲等挺水植物分布范围扩张。
3)1998年洪水从海河倒灌是邛海水生植被退化的突变性原因,导致了高枧湾沉水植被几乎全部消失;邛海运行水位抬升和透明度下降是邛海水生植物分布面积缩减和群落单一化的渐变性原因;2002年邛海出口水闸建成运行,4-5月连续15、30和45天最低平均水位分别抬升了0.47、0.44和0.42 m,影响整个邛海水生植物萌发与生长。高枧湾透明度仅为0.5~1.0 m,是影响区域沉水植物繁育生长的主要因素;西岸、南岸和东岸基底构成是水生植物生物量的主要限制性因素。
4)建议开展邛海水环境-水资源-水生态统筹治理。通过减少污染负荷输入,改善水体透明度;通过恢复邛海水位运行的自然节律,最大程度地满足春季水生植物萌发生长的需求;通过人工干预方式修复水生植被,调控食物网结构,逐步改善水生植被生境,实现邛海水生态系统健康。
5 附录
附表Ⅰ见电子版(DOI: 10.18307/2024.0135)。
猜你喜欢
东坡赤壁诗词(2020年3期)2020-07-04
花卉(2020年12期)2020-01-09
农产品市场周刊(2019年14期)2019-09-09
——以武汉紫阳湖为例
绿色科技(2019年2期)2019-05-21
消费导刊(2018年10期)2018-08-20
少年文艺·开心阅读作文(2018年7期)2018-08-12
大科技(2016年1期)2016-07-17
系统工程学报(2015年3期)2015-02-28
现代企业(2015年5期)2015-02-28
现代企业(2015年5期)2015-02-28
