
东南丘陵山区深水水库两种浮床植物脱氮效率对比研究*
2024-01-13 07:40倪子怡程新良王裕成朱广伟朱梦圆孙起亮
湖泊科学 2024年1期
倪子怡,许 海,詹 旭,程新良,王裕成,朱广伟,朱梦圆,邹 伟,孙起亮
(1:中国科学院南京地理与湖泊研究所,湖泊与环境国家重点实验室,南京 210008) (2:江南大学环境与土木工程学院,无锡 214122) (3:浙江省杭州市生态环境局淳安分局,杭州 311700) (4:浙江省杭州市桐庐县环境保护监测站,桐庐 311500)
水库对调节流域生态环境和社会经济发展具有重要作用。进入21世纪以来,全球水资源短缺形势日益严峻,人类饮用水的供给越来越依赖于水库,水库功能由发电、防洪转向供水已成为世界性的趋势。我国是水资源短缺的国家,城市缺水甚为严重,水库已成为我国许多城市的重要饮用水水源地,在水资源调配中发挥着极其重要的作用。统计显示,2019年我国水库总库容已增加到9306亿m3,大型水库座数和水库总库容均翻了一番[1]。对于水源地型水库而言,水质安全保障极为重要。然而,随着人类活动和全球气候变暖,水体富营养化逐渐加剧[2],引发的季节性藻类水华严重威胁了水生态安全,不仅恶化水质影响人体健康,还破坏了水生态系统的功能平衡,自然生态过程被中断并引发一系列的连锁反应[3]。2020年生态环境部监测资料显示,中富营养及以上的湖泊占比约为90.9%,贫营养湖库占比仅为9.1%[1]。
氮是水体富营养化的主要驱动因素之一[4]。在过去的30年里,中国陆地系统的人为氮输入增加了两倍多,大量的氮以溶解性硝酸盐形式输入河流和湖泊,加剧河湖富营养化问题,同时对饮用水供应构成严重威胁[5]。一些库湾水体不仅受陆域氮素输入影响,其水体流动性变差和库区经济社会活动加剧等因素使库湾水体富营养化问题更为突出[6]。因此水体氮素的控制成为水库生态系统的迫切任务。对于许多丘陵山区水库,尽管流域自然植被较好,水质总体保持良好,但受极端降雨和水文过程影响,面源污染加剧,而流域土地资源又非常紧张,缺乏足够面积的拦截湿地截留流域污染负荷,在这种流域进行水体污染治理工作难度更高。同时对于深水湖泊而言,由于水体容易形成温跃层,底层水温较低[7],反硝化潜力较弱,而上层水体由于处于好氧状态,同时水体碳源较少[8],缺少反硝化发生的环境,很难发生反硝化脱氮过程,尤其针对局部库湾较高营养盐浓度水体的净化工作更具挑战性。
生态浮床是一种经济、可持续的绿色水体修复技术[9],不额外占用土地,还兼具一定的经济或景观效益,其运用无土栽培技术原理,采用现代农艺和生态工程措施将水生植物或驯化的陆生植物种植于漂浮的载体上,通过植物、根系微生物的吸收同化和硝化-反硝化脱氮等过程达到净化水体氮污染的目的[10]。生态浮床除通过植物吸收和收获带走水体氮素外,浮床植物在水体中生长会形成发达的根系,为水体中各种微生物提供了良好的固着载体。植物在生长过程中也在不停向周围环境释放各种有机酸、糖和维生素等有机物[11],能够为微生物反硝化作用提供碳源从而加强氮的去除。在多项模拟实验和实际工程中,生态浮床均展现出良好的水质净化效果[12-15]。生态浮床通常应用于净化高氮磷浓度水体,如养殖水体、城市富营养化河道、污水处理厂尾水等[16-17],对于丘陵山区深水水库水体生态浮床技术的适用性研究较少。在生态浮床的长期运行中,其净化效果受多种因素影响,例如光照和营养盐浓度[18-20]。Wobbe等[21]综合广泛研究指出,严格控制植物对于光的捕获是至关重要的,因为光具有双面性:它能推动光合作用,但光照过强对部分植物是有害的。在较高光照射下经常观察到植物明显的光抑制现象和光合能力的下降[22]。氮和磷是植物生长必需的营养元素,植物因自身的生长特性,对于水体营养盐浓度有着不同的耐受性和生理反馈,进而影响水质净化效果[23]。因此,对于不同水质的水体需要选择适宜的植物品种以深度净化水质。
千岛湖,又名新安江水库,是浙江省最为重要的饮用水源保护地[24]。近年来,千岛湖水质面临着诸多威胁,如2007年坪山水域中水华束丝藻异常增殖,2009、2010年安阳水域出现曲壳藻、鱼腥藻异常增殖等现象。大量氮输入导致千岛湖湖区以含氮物质污染为主,水质下降,并成为引发富营养化和藻类水华暴发的重要因素,且在局部库湾更为严重[24-25]。因此,控制千岛湖水体氮浓度、降低藻华暴发风险十分必要。然而,千岛湖是深水水库,岸坡陡峭,生境单一,流域土地资源紧张,缺乏足够面积的净化湿地[26]。同时水体碳浓度很低,5-8月千岛湖全湖水体溶解性有机碳浓度均值仅为1.73 mg/L[8],湖体反硝化脱氮作用受到明显的碳限制,水体氮素自净能力较低。目前的研究多集中于通过室内模拟实验探究生态浮床对于高氮磷浓度污水的净化效果及其影响因素,野外环境中对于像千岛湖这种深水水库且碳源不足背景下生态浮床的可行性研究较少。此外,探究光照强度对水生植物的影响多聚焦于沉水植物,在此种水质背景下不同类型浮床植物受营养盐浓度和光强影响的对比研究较为缺乏。
鉴于此,本研究选取湿生植物空心菜(Ipomoeaaquatica)和水生植物粉绿狐尾藻(Myriophyllumaquaticum)两种不同类型的典型植物为对象,通过在千岛湖水体开展模拟实验,对比研究营养盐浓度和遮光水平对两种植物生长和水体氮素净化的效果及其机制,以期为我国类似水库氮素深度净化提供科技支撑。
1 材料与方法
1.1 实验区域
千岛湖(29°22′~29°50′N,118°36′~119°14′E),位于浙江省杭州市淳安县,是1957年开建、1959年9月21日截流蓄水的新安江水库[27]。水库水面面积580 km2(水位为108 m时),库容为178亿m3,平均水深31 m,最大水深100 m。千岛湖地处典型的亚热带季风气候区,温暖湿润,四季分明[28]。近60年来气温逐步升高,极端天气增多,2020年平均气温18.3℃,较历史平均值(17.3℃)稍高。年均降水量为1636.5 mm,年日照时数达1951 h[2]。根据千岛湖生态系统研究站自动监测数据显示,7-9月小金山断面处平均有效光照为241.76 W/m2。新安江是最主要的入库河流,约占入库径流的2/3[29]。千岛湖的营养盐达标问题长期困扰着水库环境的治理,其水体总氮(TN)、总磷(TP)浓度时空差异明显,淳安县境内以西北库湾最高,全年均值分别为1.18和0.031 mg/L,其次为中心库区,从时间来看,夏季营养盐浓度普遍较高,TN和TP浓度平均可达0.91和0.025 mg/L[27]。采样点位于国控监测断面小金山(图1),水深约40 m,距上游浙江省与安徽省交界的街口断面约28.6 km,极易受上游河流断面的影响,从而使水质无法稳定达标[30]。实验区域位于小金山水域一库湾,四周无光线遮挡。
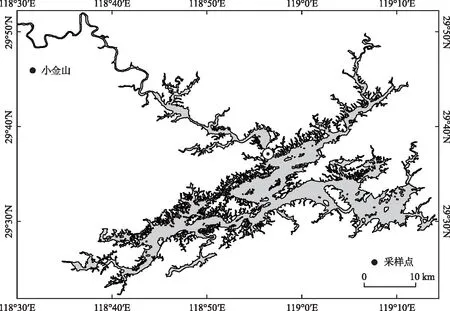
图1 实验区域方位Fig.1 Location of research area
1.2 供试植物
空心菜属于一年生蔓生植物,能耐38℃高温,对不同浓度水体均具有很强的适应性和氮磷去除效果,根系发达且生物量大,是理想的浮床植物材料。
粉绿狐尾藻,俗称“净水精灵”,是一种多年生沉水或浮水植物,在南亚热带地区可全年生长,生态风险低,对富营养化水体的生态治理具有得天独厚的优势[31]。
1.3 实验设计
实验于2022年8月9日-9月6日在浙江省千岛湖生态系统研究站进行。选用2.2 m×1.2 m×3.0 m的帆布方形围隔作为生态浮床主体,内置2 m×1 m的尼龙格网作为载体,每个格网统一放入初始生物量为2 kg的植物(种植密度为0.76 kg/m2),置于千岛湖小金山断面湖水中以模拟原位生态浮床效果,实验布置见图2。选取扦插苗空心菜和粉绿狐尾藻作为实验植物。实验开始前,植物先预培养一周,待植物长势稳定后选取长势较好且相近的植物,用于生态浮床模拟实验。

图2 实验布置(上图为原位湖水处理,下图为氮磷加富处理)Fig.2 Experiment layout (the above diagram is in-situ lake treatment, the below is nitrogen and phosphorus enrichment treatment)
模拟实验包括2种营养盐浓度和3种光强水平。营养盐水平包括:原位湖水(即将植物固定于格网上,置于小金山断面原水下自然生长)和氮磷加富(即在围隔中进行,抽取小金山原水并在此基础上添加氮磷营养盐)。因为原位水体营养盐浓度较低,前期预实验显示水体一旦装于培养桶内,很快就被植物吸收导致植物因营养衰竭而死亡,因此设置湖水原位培养处理,使植物能够处于稳定的营养盐浓度环境,用于测试原位水体营养盐浓度下植物是否能够生长。氮磷加富处理是在围隔中进行,主要是测试氮磷浓度高于原位水体时植物的生长状况,验证原位水体植物生长是否受到氮磷限制。实验用水采自千岛湖小金山断面水域,用硝酸钾和磷酸二氢钾试剂分别配制氮、磷营养液,两组实验初始水样的物理化学指标如表1所示。光强水平包括:遮光0%(全阳光)、遮光40%和遮光75%,通过将不同孔径尺寸的遮阳网覆盖于格网上,以达到目标遮阳水平。因实验在野外千岛湖湖体原位培养,为避免降雨等因素干扰,顶部设有透明防雨棚覆盖了整个围隔组(包括0%、40%和75%)。连续3天分别于9:00、12:00和15:00用德力西照度计(DLY-1801)测量各遮光处理的实际光照强度,见表2。以无植物、氮磷加富和遮光0%组合处理为实验对照。实验共设13个处理,每个处理有3个平行。
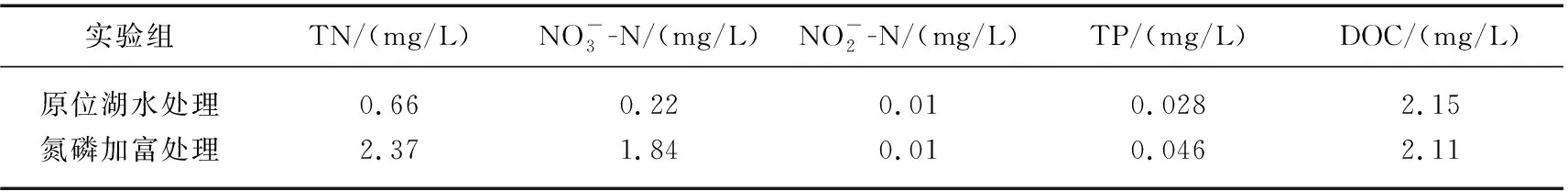
表1 实验初始水质指标Tab.1 Initial water quality index

表2 不同遮光处理的实测光强Tab.2 Measured light intensity of different shading treatments
1.4 植物生长指标测定
由于原位湖水组处于开放水体,主要度量植物长势,不对其进行水体营养盐浓度测定。在实验开始和结束时期随机抽取植物,用直尺测量植物单株株高、根长,用电子天平(常熟意欧电子天平,DT502)称量植物生物量(湿质量)。随后烘干称重,过100 mm×25.4 mm筛网研磨后测量植物体内氮磷含量(意大利元素分析仪,Eurovector EA3000)。
1.5 水体理化指标测定
1.6 植物根系反硝化速率测定
取氮磷加富组各处理第14天植物进行根系反硝化速率测定,每个处理设3个平行。培养袋选用容积为2 L的聚氟乙烯取样袋,外设2个阀门,三边封口,一边配密封条,厚度为50 μm,具有良好的密闭性能。将植物置于培养袋中,加入1.5 L与初始氮磷加富处理同等营养盐浓度的湖水,排尽培养袋中气体并用密封条进行密封,黑暗恒温20℃培养8 h。
分别在实验初始(0 h)和结束(8 h)通过培养袋出水阀将水缓慢抽入到12 mL空瓶(Labco Exetainer)中,瓶子装满时继续引流并缓慢抽出橡皮管,使液面形成凸状,然后拧紧瓶盖。每个培养袋取3个平行样,采集完后立即用注射器打入50%质量分数的ZnCl2溶液终止其微生物反应并密封保存。
样品采用膜接口进样质谱仪(Bay instrument, USA)测定,并使用Kana等[34]提出的N2∶Ar比法进行计算。由于氩气(Ar)稳定的化学性质,而氮气(N2)受生物过程影响,因此使用N2∶Ar方法的计算精度远高于单独使用N2,计算方法如式(1)所示:
Δ[N2]=[N2]/[Ar]×[Ar]*-[N2]*
(1)
式中,Δ[N2]为N2浓度相对于自然条件下的净增量,μmol/L;[N2]/[Ar]为经质谱仪矫正过的水样中N2与Ar浓度比值;[N2]*、[Ar]*分别为特定温度、盐度条件下的理论平衡浓度,μmol/L,通过标准水样温度和盐度值代入Weiss[35]的方程计算得到.
1.7 水体污染物去除率计算
污染物去除率(η)计算公式为:
(2)
式中,C0为污染物起始浓度,mg/L;Ci为第i次取样的污染物浓度,mg/L。
TN净去除效率(μ)计算公式为:
(3)
式中,μ为TN净去除效率,mg/(m2·d);ΔC0为对照处理开始与结束污染物浓度差,mg/L;ΔCi为植物处理开始与结束污染物浓度差,mg/L;V为水体体积,L;S为面积,m2;t为时间,d。由于空心菜处理氮浓度在14天时已经耗尽,空心菜处理按14天计算,粉绿狐尾藻处理按28天计算。
植物同化吸收氮速率(γ)的计算公式为:
(4)
式中,γ为植物同化吸收氮速率,mg/(m2·d);m1和m2分别为初始和结束时植物干质量,g;ρ1和ρ2分别为初始和结束时植物体内氮含量,mg/g。
1.8 统计分析
实验数据采用IBM SPSS 27.0软件进行方差分析,使用单因素ANOVA检验法进行差异显著性分析(P<0.05),采用Origin 2018软件作图。
2 结果与分析
2.1 氮磷浓度和光强对植物生长的影响
实验期间空心菜和粉绿狐尾藻单株生长情况如表3、4所示。原位湖水和氮磷加富条件下的两种植物均生长良好,实验后其株高、根长、生物量均比实验前显著增加。空心菜的相对生长速率平均约为0.049 g/(g·d),氮磷加富下空心菜的长势明显更好,其株高和生物量均是原位湖水下的1.6倍以上,根长增量约为25.86 cm。并且氮磷加富下以遮光40%环境中的空心菜单株株高、根长和生物量最高,株高和生物量分别是遮光0%环境下的1.3和1.5倍左右。遮光75%处理组由于遮阳网密实以及光照不足,实验中后期植株腐烂衰亡,因此于实验第21天时将植物移除。
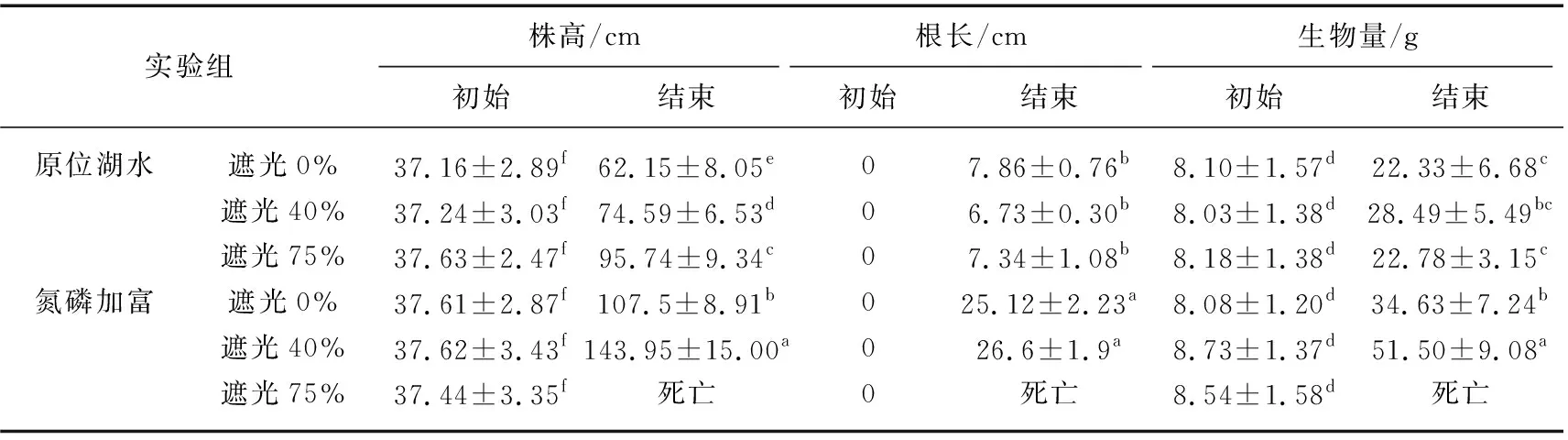
表3 空心菜单株生长情况*Tab.3 Single plant growth of I. aquatica

表4 粉绿狐尾藻单株生长情况Tab.4 Single plant growth of M. aquaticum
不同处理下粉绿狐尾藻的相对生长速率约为0.026 g/(g·d)。粉绿狐尾藻株高受营养盐影响显著,氮磷加富下平均株高相较原位湖水增加约6.84 cm,但单株根长与生物量随氮磷添加均无明显影响,均值分别为4.18 cm和9.93 g。此外,光强对粉绿狐尾藻长势的影响很小。
实验结果表明,空心菜长势受营养盐浓度和光强影响显著,氮磷加富和遮光40%的环境下生长最为迅速。相反,粉绿狐尾藻生长受氮磷浓度和光照影响均较小。
2.2 不同光强下生态浮床对水体氮素的净化效果
水体总氮浓度变化及去除率如图3、4所示。实验期间,空白对照组TN浓度基本维持稳定,均值为2.15 mg/L。观察发现,前期空心菜处理水体TN浓度快速下降,在第14 天基本降至最低,其中以遮光40%下TN去除率显著高于其他遮光水平,达82.33%。实验结束时由于水体可利用氮素基本消耗殆尽,空心菜不同遮光处理间TN去除率无明显差异,平均约为74.88%。对比空心菜,在前期粉绿狐尾藻处理组水体TN浓度下降不明显,但其去除率随时间而增加。实验结束时,粉绿狐尾藻处理组不同遮光水平下水体TN浓度均值约为1.46 mg/L,其中以遮光75%下水体TN去除率最高(43.18%),显著高于其他遮光处理。
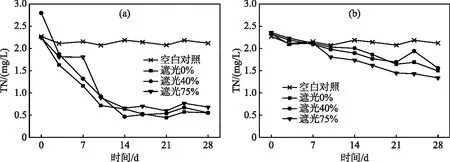
图3 水体TN浓度变化((a)对照+空心菜;(b)对照+粉绿狐尾藻))Fig.3 Variation of total nitrogen concentration ((a) control + I. aquatica; (b) control + M. aquaticum))
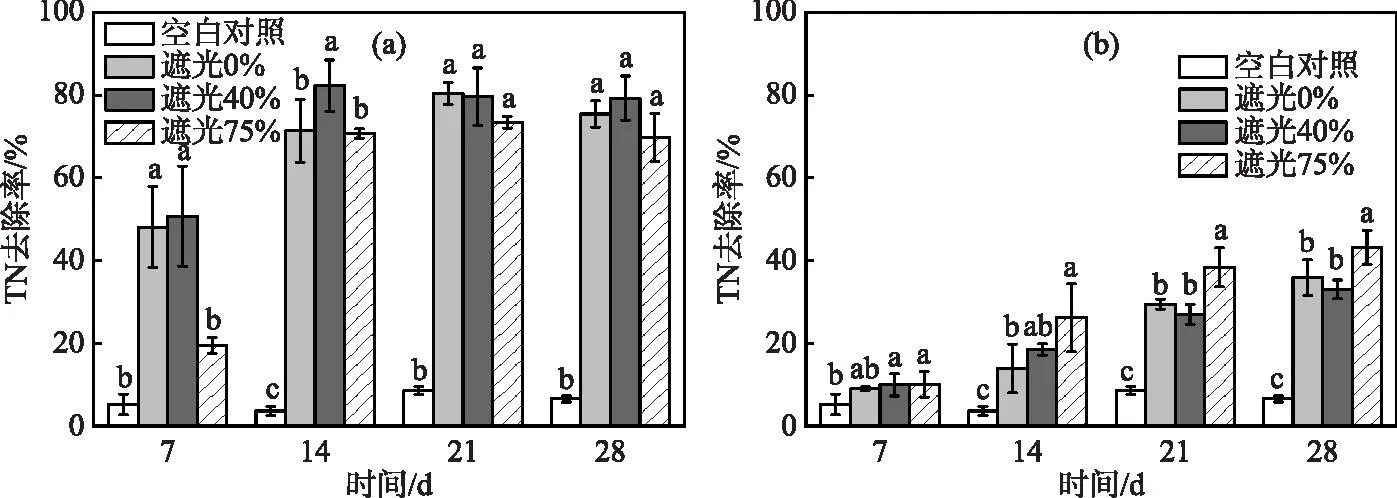
图4 水体TN去除率变化:(a)对照+空心菜;(b)对照+粉绿狐尾藻(字母不同表示不同处理之间差异显著(P<0.05),字母相同表示差异不显著,下同)Fig.4 Variation of total nitrogen removal rates ((a) control + I. aquatica; (b) control + M. aquaticum))
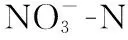
图5 水体浓度变化((a)对照+空心菜;(b)对照+粉绿狐尾藻))Fig.5 Variation of nitrate concentration ((a) control + I. aquatica; (b) control + M. aquaticum))
空心菜的TN净去除效率(171.55 mg/(m2·d))约是粉绿狐尾藻(36.85 mg/(m2·d))的4.66倍。其中,空心菜在遮光40%环境下的TN净去除效率最高,为213.30 mg/(m2·d),比不遮光处理组提高35.5%。粉绿狐尾藻在遮光75%环境下的TN净去除效率最高,为44.23 mg/(m2·d),约是遮光40%环境下的1.41倍(图6)。
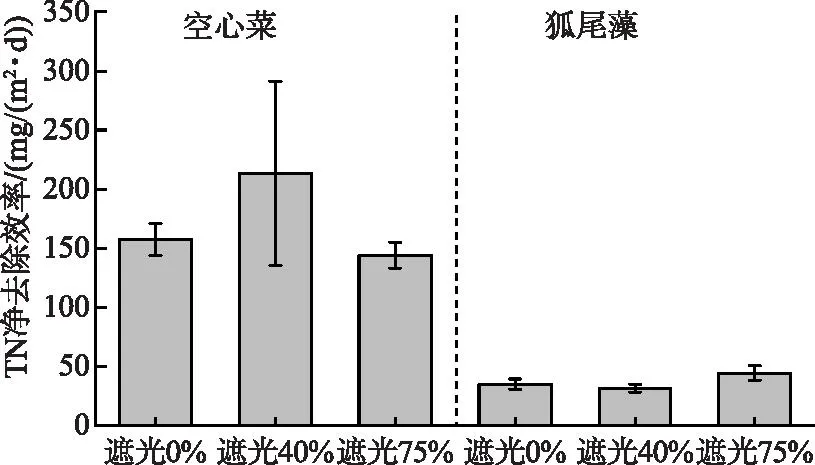
图6 不同处理TN净去除效率Fig.6 Net total nitrogen removal efficiency in different treatments
综上所述,空心菜和粉绿狐尾藻对于水体氮浓度均有良好的净化效果,空心菜对TN的净去除能力约为粉绿狐尾藻的4.66倍。其中,遮光40%和75%的环境分别提升了空心菜和粉绿狐尾藻对氮的去除效果。
2.3 不同光强下生态浮床植物氮素去除途径分析
实验期间空心菜的干物质氮含量平均从30.39 g/kg降低至24.93 g/kg。但相较于不遮光环境,遮光40%下的的空心菜体内氮含量显著增加,为27.59 g/kg。与空心菜不同,实验后粉绿狐尾藻的干物质氮含量略增加,平均从8.53 g/kg升高至9.53 g/kg。不同遮光条件下粉绿狐尾藻的体内氮含量,以遮光75%环境下最高,为10.00 g/kg(表5)。

表5 植物体内氮含量的变化Tab.5 Variation of nitrogen content in plants
遮光0%和40%下空心菜同化吸收氮速率分别为142.54和161.12 mg/(m2·d),不同环境下空心菜同化吸收作用占TN去除的贡献率均在70%以上。对于粉绿狐尾藻,其同化作用贡献率约为50%,以遮光75%下同化吸收氮速率最高(27.81 mg/(m2·d)),并且反硝化及其他作用氮去除速率同样达到最高值,为24.00 mg/(m2·d)(图7)。
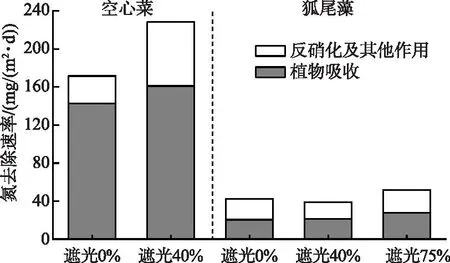
图7 不同处理氮去除途径速率Fig.7 Nitrogen removal pathway rates in different treatments
空心菜处理根系整体反硝化作用强于粉绿狐尾藻,反硝化速率平均值分别为5.79和4.12 μmol N2/(L·h),均高于空白对照组(2.83 μmol N2/(L·h))。其中,遮光40%下的空心菜根系反硝化速率最高,为7.60 μmol N2/(L·h),显著高于其他处理。对于粉绿狐尾藻,以遮光75%条件下的粉绿狐尾藻反硝化速率最快,为4.65 μmol N2/(L·h),以遮光40%环境下最低,为3.65 μmol N2/(L·h),差异显著(图8)。

图8 不同处理根系反硝化速率Fig.8 Denitrification rate of root system in different treatments
3 讨论
3.1 不同营养盐浓度和光照强度对两种浮床植物生长的影响
植物作为生态浮床的核心组成部分,生物量越高对水中污染物的吸收和净化能力越强[36-37],但植物生长往往受到关键环境因素的限制,这些因素可能会影响其水质净化效率。空心菜已经广泛应用于生态浮床水质净化中,杨绍聪等[38]研究发现水培空心菜产量显著高于土栽,且更有利于根须生长。本研究中空心菜的相对生长率明显高于粉绿狐尾藻,但空心菜体内氮含量却略微降低。究其原因,一方面在于空心菜原本种植于高浓度池塘水体中,另一方面在于生物量大幅增加导致的“稀释效应”。由此说明,空心菜吸收的氮素主要用于合成自身物质结构,生物量快速积累。粉绿狐尾藻吸收的氮一部分用于富集在植物组织中,一部分用于自身生长繁殖。
空心菜的生长受营养盐和遮光水平影响显著,在氮磷加富和遮光40%的环境下生长得明显更好,这表明低营养盐浓度、全光照和75%遮光环境均会在一定程度上抑制空心菜的生长。空心菜的株高随遮光程度显著增加,这与前人的研究相似,前人研究发现,在遮荫条件下植物茎组织伸长,认为这是植物体内生理机制的调控方式之一,通过调节体内生长素以适应外界刺激[39],也通常被解释为伸长组织以获取更多的光照[40]。光照虽是植物能量和物质的来源,但也可能成为其潜在的胁迫因素[41]。本研究发现,氮磷加富组在遮光40%下空心菜的单株生物量最高,推测遮光40%的遮阳网为空心菜创造出了最适宜其生长的小气候环境,既避免夏季强光直射对空心菜生理器官的损伤,又能迎合其喜光的特性以保障光合作用的顺利进行。正如杨小锋等[42]在夏季对空心菜覆盖不同材料发现,透光率为65%的黑色遮阳网覆盖的空心菜质量增加最大,并且遮光还能提升空心菜的卫生品质。但是在原位湖体组,未发现空心菜生物量有显著差异,这主要是因为在低营养水平下,营养盐的影响比光强更大,水下部分对于养分的竞争更为重要[43]。此外,由于同一物种的不同性状特征也会对光强产生不同的反应,光照强度对其根长并无明显影响[22]。
本研究中营养盐浓度的增加对粉绿狐尾藻的生物量和根长无明显影响,说明粉绿狐尾藻对于水体营养盐浓度具有广泛的适应性。和Tan等[44]的结果一致,发现在低营养盐水体中氮磷浓度的增加对狐尾藻的生长几乎没有影响。这也印证了Chambers等[45]的观点,即沉水植物对富营养化的敏感性不受水体中氮、磷浓度变化所直接引起的生理反应的影响。此外,水生植物粉绿狐尾藻属于阴生植物,其光饱和点及光补偿点比陆生阳生植物低很多[46]。本研究中粉绿狐尾藻的单株生物量在不同光照条件下均增加了一倍以上,说明实验所设置的光照强度在粉绿狐尾藻的耐受范围内,遮阳75%的环境也能满足其对光照的需求。关于光照对狐尾藻生长的影响还未有定论,有部分学者认为随光照强度的增加,狐尾藻叶片叶绿素含量下降,丙二醛含量上升,表现出明显的强光抑制现象[22,47],也有研究认为狐尾藻的根系和生物量随光照强度的增加而明显提高[44,48]。本实验中光照对于狐尾藻的生长影响很小,几乎没有差异,桑雨璇等[49]经研究同样发现此现象,认为原因在于植物对光照的需求。综上所述,本研究认为空心菜的生长受营养盐和光强影响显著,但夏季营养盐浓度和光照对于粉绿狐尾藻生长的影响较小。
3.2 不同光强下两种浮床植物对氮素的净化途径解析
不同植物对水体营养盐去除效果存在显著性差异,这种差异主要与植物类型及其生理特性有关[26],本实验中空心菜对于氮的去除率显著高于粉绿狐尾藻,多项研究通过对比几种植物发现空心菜在水质净化方面的价值和突出特点[12,16,50-51]。温度也会直接或间接地影响植物的生长和氮吸收[52]。实验期间平均水温约为32℃,对于空心菜来说处于最佳范围,粉绿狐尾藻适宜在16~26℃下生长[26],实验结束时空心菜的相对生长速率约为粉绿狐尾藻的2倍,而生态浮床中植物的净增量和其对氮、磷的直接吸收以及增效作用密切相关[53]。
生态浮床中氮的去除途径主要包括植物吸收和微生物硝化-反硝化还原成N2作用[51,54]。本研究发现空心菜对于水体氮去除以植物同化作用为主,占比70%以上,对于粉绿狐尾藻,同化吸收和微生物作用均是其重要的脱氮途径,各占比50%左右。Chen等[55]得到相似的结果,认为空心菜因其体内组织氮含量高的特性,脱氮以植物吸收为主。而关于生态浮床脱氮的主要途径还存在争议,有部分学者认为植物吸收氮量只占系统总去除氮量的很小部分(低于20%),大部分氮去除依靠微生物作用[51,56],造成此现象的原因在于受试水质的差异,相对来说,在高负荷背景下植物对营养盐的吸收积累能力有限。千岛湖属于较清洁水体,氮浓度相对较低,可利用氮素首先满足植物迫切的生长需要。其次,硝态氮作为反硝化的底物,一直被认为是影响反硝化速率的首要因素[57],加之水体碳源不足的特点进一步限制其反硝化速率。而遮光水平对于两种植物同化吸收的影响主要是通过改变其长势,遮光40%的环境显著提高了空心菜的生物量和体内氮含量,进一步提高其同化吸收氮速率。一般来说叶绿体数量与组织氮含量呈正比,由此还能提高空心菜的光合效率。而在不同光强下粉绿狐尾藻体内氮含量差异不显著,对于氮的吸收速率影响较小。正如Yuan等[58]发现即使光控制实验中会对粉绿狐尾藻的生长产生影响,但其体内氮含量几乎没有差异。
比较根系反硝化作用可知,空心菜根系反硝化速率明显高于粉绿狐尾藻,其中遮光40%下的空心菜处理最高,而粉绿狐尾藻处理以遮光75%环境下最佳,这与根系伸长和根系分泌物密切相关。在初始均无根系的情况下,实验结束时空心菜的根系长度(约25 cm)明显高于粉绿狐尾藻(约4.7 cm),发达的根系能够为微生物提供更大的附着面积[19],特别是空心菜的根系聚集了大量的反硝化细菌,大大加速了脱氮进程[59]。此外,Yang等[51]研究发现空心菜根系分泌物和径向氧损失显著高于其他植物,与其总氮去除效率呈显著正相关关系。径向氧损失有助于在植物根际构建好氧-缺氧-厌氧微环境,为微生物的硝化-反硝化作用创造理想环境条件[60-61]。根系分泌物的主要成分是小分子有机物,更容易被微生物利用,可以促进反硝化相关酶活性的提高[51],并为反硝化过程提供碳源[62]。朱辉翔等[63]对比4种材料发现狐尾藻的释碳能力最低,这也是造成其根系反硝化速率较低的原因之一。
3.3 千岛湖生态浮床推广的管理建议
适宜的营养盐浓度能够促进植物生长,进而使植物快速稳定地吸收水体中的污染物[64]。然而,并非所有植物均随营养盐浓度的增加而长势更好,当水体中的氮、磷浓度超过植物生长所需时,反而会威胁植物生长。比如Xu等[23]发现马来眼子菜在低营养盐浓度水体中生长情况和水质净化效果更好,认为沉水植物对营养盐的耐受程度较低。本研究中空心菜和粉绿狐尾藻这两种植物对水体营养盐浓度呈现出不同的响应情况,空心菜适合在较高氮、磷浓度水体下生长,而粉绿狐尾藻受营养盐浓度影响较小。千岛湖是华东地区最大的水库,水体TN浓度空间差异明显,其中安徽段为1.60 mg/L,淳安县境内均值为0.89 mg/L,TN浓度沿新安江主航道方向呈逐步下降趋势,最低值在西南库湾,约0.73 mg/L[27]。位于水库库尾河口区的街口断面是污染最严重的水域,承接60%的流域来水[28,65]。朱广伟等[27]指出夏季千岛湖蓝藻水华异常增殖现象明显,甚至有蓝藻水华风险,其中8月东南库湾出现蓝藻生物量峰值。西北库湾在4、5月也出现了明显的硅藻异常增殖现象,而营养盐控制是降低藻类水华风险的根本之策。在生态浮床的实际推广中,可以根据不同水体富营养化因子特征,优化植物配置,以达到更好的水体富营养化防治效果[66]。本研究基于对空心菜和粉绿狐尾藻这两种植物水质净化效果的研究,认为夏季在氮污染严重以及藻华高风险暴发水域,例如街口断面、营养盐浓度高的库湾、污水厂排放口等,为能够最大限度净化水质,建议生态浮床以种植空心菜为主,并配套铺设遮光40%的遮阳网以进一步提高其产量和水质净化效果。正如Chen等[16]研究发现在空心菜与其他植物组合系统的水质净化效果并没有优于单一空心菜种植系统,因为空心菜几乎承担组合系统中一半的养分积累。在中低浓度的水域,建议采取空心菜和粉绿狐尾藻混合种植模式,并分别覆盖遮阳40%和75%的遮阳网。既能弥补粉绿狐尾藻夏季较低的氮净化效率,多种植物种类又能提高水体微生物多样性,进而加强微生物的降解作用[67],以实现经济和环境的双重效益。此外,遮阳网与生态浮床的耦合能够进一步降低藻类水华暴发的风险,这是生态浮床载体、遮阳网的遮光、根系分泌物和化感物质等共同作用的结果[19]。值得注意的是,空心菜的生长周期一般在30天左右,Zhou等[68]指出应在植物腐烂前收割,否则储存在水上部分的营养物质就会再次进入水中,并且Wang等[69]发现夏季植物水上部分储存了最高浓度的营养,秋季资源会向水下部分转移。因此建议夏季定期对空心菜进行刈割,并在冷季到来之前整株收割以永久去除污染物。研究结果可为于千岛湖生态浮床的规模化推广和管理提供科技支撑。
4 结论
1)夏季空心菜生长受营养盐浓度和光照影响明显,氮磷加富以及40%遮光能为其提供最适宜生长的环境;而粉绿狐尾藻长势受氮磷浓度和光照影响较小。
2)空心菜浮床氮净化效果显著高于粉绿狐尾藻。空心菜脱氮以同化作用为主,遮光40%的环境通过明显提升空心菜吸收氮量和根系反硝化速率,大幅提升水质净化能力。对于粉绿狐尾藻,植物同化和反硝化作用贡献率各占50%左右。
3)在千岛湖生态浮床规模化推广中,建议夏季在营养盐浓度相对较高的水域种植空心菜,并覆盖遮光40%的遮阳网;在中低营养盐浓度水域,建议采取空心菜和粉绿狐尾藻组合种植模式,并分别覆盖遮光40%和75%的遮阳网,以提升环境和经济双重效益。
致谢:感谢千岛湖生态系统研究站在野外实验中提供的帮助与支持,感谢中国科学院南京地理与湖泊研究所吴铭杰、朱慧、孙宏伟、胡春华、刘莉、李昌杰等在样品采集及水质测定中提供的帮助。
猜你喜欢
水产养殖(2022年2期)2022-11-05
人文天下(2022年7期)2022-09-15
绿色科技(2020年24期)2021-01-09
今日农业(2019年14期)2019-09-18
——千岛湖站
法人(2019年1期)2019-01-10
福建畜牧兽医(2016年5期)2016-12-16
环境科技(2016年1期)2016-11-08
快乐作文·低年级(2016年2期)2016-02-24
小学生作文选刊·中高年级版(2015年9期)2015-09-23
恋爱婚姻家庭·养生版(2013年9期)2013-05-14
