
青藏高原东南缘三江并流区植被物候时空变化及其影响因素分析
2024-01-12 00:06谢作轮肖冬梅黄浩鸿陈志云刘友存
嘉应学院学报 2023年6期
谢作轮,肖冬梅,黄浩鸿,蒋 娜,陈志云,刘友存
(嘉应学院 地理科学与旅游学院,广东 梅州 514015)
0 前言
植物物候是指自然界中植物受各种环境要素影响而出现以年为周期的各种现象,如植物的萌芽、展叶、开花、结果、落叶等现象[1-3].其作为一种综合性响应指标[4],是陆地生态环境条件季节和年际变化最直观、最敏感的生物指示器[5-6],对自然环境变化的响应较为敏感,可以有效地反映全球环境的变化状况[7].第六次IPCC 评估报告指出,过去20 年间全球气温上升约0.99 ℃[8].受全球气候变暖影响,植被物候发生了不同程度的变化[9],近年来,在植被物候的时空变化、物候与气候及其他环境因子的相关性研究取得较大进展.在中纬度区域,开展了物候变化研究,文献[10]发现温度是制约北半球中高纬度地区的植被春季物候的主要因素;针对生态环境脆弱区域的气候变化对植被物候的影响比较明显,但各个区域的植被物候变化对气候因子的响应不同,文献[12]发现伏牛山地区森林植物受三月份平均气温下降的影响生长季始期出现延迟趋势.文献[11]利用AVHRR NDVI3g(1983—2011)数据集研究了尼日利亚植被物候对降水和土地覆盖变化的响应;文献[13]发现米仓山区春季在水热条件充足下植物生长发育开始阶段提早,而秋季受雨水的增加使得植物生长发育末期提早.文献[14]发现植被物候在寒带地区主要受温度的影响,而温带地区则受降水和气温的共同影响.目前,国内对植被物候变化研究集中于北方地区、青藏高原以及祁连山脉[15-17],对西南地区的研究较为缺乏[18].
三江并流区位于青藏高原向云贵高原过渡的横断山脉的纵谷地带核心区,生物种类丰富,是全球生物多样性保护热点地区[19].该地区内气候特征复杂多样,山地垂直地带性差异明显,地表生态系统脆弱[20].本文基于2001—2019 年MODIS EVI 数据、DEM 数据、土地利用数据及气象数据等,研究三江并流区植被物候的长时间序列变化趋势和空间分异规律,探讨植被物候在时空演化对气候变化的响应,以期为保护与改善生态环境、延续三江并流区复杂多样的生态系统提供理论参考,并揭示该地区植被活动对全球变暖趋势下的响应机制.
1 研究地区和研究方法
1.1 研究地区概况
三江并流区地处青藏高原东南缘,位于云南、西藏与四川的交界地带(25°30′~29°N,98°~100°30′E),属于横断山脉纵谷地带,区域内地质构造活动活跃,地势起伏大,河谷纵切,怒江、澜沧江、金沙江并行而流,涵盖范围达1.70 km2(图1).研究区天气受夏季风和青藏地区的高原气候的相互影响,全年降水主要集中在5 至10 月份,11 至翌年4 月份为旱季,加之当地平均海拔垂直高差将近6 000 m,峰高谷深,气候具有明显的季节性变化和地形垂直分化[21].由于地理和气候状况的复杂多变,该区域汇集了北半球由热带至寒带八个分带的各种气候类型和生物群落,物种种类丰富,堪称“世界生物基因库”,也是地球最直观的体温表和中国珍稀濒危动植物的避难所[22].

图1 研究区的地理位置及其地形特征
1.2 数据来源和预处理
1.2.1 EVI 数据
1.2.2 土地利用数据
采用Globeland30 全球地表覆盖数据(30 m,2000 年、2010 年和2020 年3 期)和清华大学宫鹏教授课题组的全球土地覆盖数据FROM-GLC(30 m,2015 年1 期).先将两个数据集重采样至250 m,再将Globeland30 提取三江并流区内耕地、森林、草地和灌木地的像元,获得三江并流区植被类型数据,作为研究区域植被物候参数提取的掩膜文件,最后利用FROM-GLC 数据对研究区域不同海拔带的物候状况进行研究.
1.2.3 气象数据
采用中国气象数据网(http://data.cma.cn)的气象数据,选取三江并流区及其附近地区的15 个气象站点的逐日平均气温、逐日降水量进而计算月均温、月降水量数据,再运用ANUSPLIN 工具将空间上离散的气象数据插值为90 m 的连续栅格数据,最后将插值结果重采样为250 m,并与物候数据进行相关分析.
构型创新设计环节将制图教学与三维构型和二维图样生成相结合,既增加了学生对本课程的学习兴趣和求知欲望,又培养了构型设计能力,对提高学生综合素质起到了积极的作用。实践证明具有良好的可操作性。
1.2.4 DEM 数据
采用NASA(https://www.earthdata.nasa.gov)发布的DEM 数据,其空间分辨率为30 m×30 m,根据三江并流区的地形属性特征与空间插值要求,首先将其重采样为90 m 的空间分辨率,再与气象数据进行空间插值.
1.3 研究方法
1.3.1 重构时间序列
MODIS-EVI 数据集采用最大值合成法(MVC)处理,虽然能够在一定的程度上有效减少大气云雾对EVI 像元的干扰,但由于受传感器、遥感卫星飞行环境、数据传输误差和土壤地表形态变化等诸多随机因素变化的综合影响,仍存在较多的随机噪音,需要通过平滑处理来重构时间序列.TIMESAT 软件提供三种平滑方式,包括S-G(Savitzky-Golay)滤波算法、D-L(Double-Logistic)滤波算法和A-G(Asymmetrical-Gaussian)滤波算法等[23-25],可以有效去除随机噪声.结合各算法的优劣性和精度差异[26],经过多次试验对比,发现S-G 滤波法具有良好的保真性,能够较好地重构三江并流区EVI 数据的时间序列.
1.3.2 动态阈值法
动态阈值法可以结合不同时期和区域的实际情况,动态地给EVI 设定不同的阈值,最终确定植被物候参数.考虑到三江并流区地理特征和EVI 时序变化状况,结合前人的研究成果[27-28],本研究将EVI 植被指数生长曲线左右振幅的阈值分别设置为30%和20%,并逐像元提取逐年的植被物候参数.在时间序列上,每个时期超过左振幅阈值的日期节点定义为生长季始期(SOS)、低于右振幅阈值的日期节点为生长季末期(EOS),两者的差值为生长季长度(LOS).最后采用年序日对三江并流区的植被物候期进行换算,即把物候期期数转化为距每年的1 月1 日的实际天数.
1.3.3 Anusplin 插值法
Anusplin 是基于普通薄盘与局部薄盘样条函数插值理论的工具[29].局部薄盘光滑样条是对薄盘光滑样条原型的扩展,可引入多个因子作为协变量与气象要素进行空间插值[31].已有研究表明,Anusplin 方法更适合于对我国西南山区的气象数据进行插值[32].
1.3.4 趋势分析与检验
采用非参数化估计方法Theil-Sen 斜率(T-Sen)[33]来计算三江并流区植被物候参数的变化趋势[13],并引入Mann-Kendall(M-K)方法来对植被物候参数时间序列趋势的显著性进行检验.本研究中根据T-Sen的计算检验结果和M-K 统计检验,分别把植被物候变化趋势及显著性分成四个层次(见表1).

表1 植被物候变化趋势及显著性
2 结果与分析
2.1 三江并流区植被物候空间分布特征
三江并流区地形落差大、气候复杂,为了研究该地区植被物候的空间分布规律,计算了2001—2019年三江并流区植被的生长季始期(SOS)、生长季末期(EOS)、生长季长度(LOS)的多年均值(见图2).SOS整体上呈由东部向西部延迟、河谷地带早高山地带晚的分布格局,时间集中在80—140 d(3月中旬至5月中旬),占植被总面积约86.6%.东北部地区SOS较早,多为第60—80 d;西部地区SOS较晚,受地形影响,呈现条带状间隔分布,高海拔地区多为第120—140 d;其余地区SOS多为第80—120 d.EOS整体上呈东部高山地区结束早、西部河谷地区结束晚的空间分布格局,时间集中在290—330 d(10月中旬至11月下旬),占植被总面积约92.5%.东部地区EOS较早,多在第310 d之前;西部地区整体上EOS较晚,最晚区域位于南部河谷地带,多在第330 d之后.LOS整体上时间集中在180—240 d,约占植被总面积的85.4%.西部地区生长季长度呈现纵列分布,河谷长高山短;但在东北部部分河谷区域生长季长度较短,是因为该区域平均海拔高,受高原气候以及植被类型的影响,植被集中在夏秋季生长,生长季长度短.
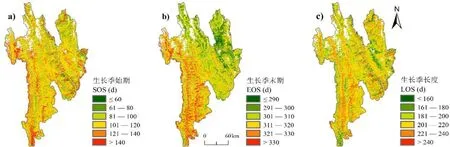
图2 2001—2019 年三江并流区植被物候多年均值空间分布
基于已有研究[35-37],利用DEM、FROM-GLC数据以及植被物候的多年均值,选取50 m高程为间隔,研究三江并流区内植被物候对植被类型和海拔因素的响应机制(见图3).海拔在1 800 m以下的区域,其主要植被类型为草地和暖温性针叶林,在该海拔段内,气候呈现为干热特征,整体上SOS、LOS主要受降水影响,而EOS受温度影响;随着海拔的升高,温度降低,降水增加,冬春旱情影响减小,SOS提前,而EOS因气温降低而提前,但整体上LOS延长.海拔在1 800~3 300 m内的区域以暖温性针叶林、温凉性针叶林和常绿混交林为主,整体趋势为SOS和EOS提前、LOS缩短.海拔在3 300~4 100 m的区域以寒温性针叶林和针叶—阔叶混交林为主,但在部分海拔段受植被类型影响,物候期具有差异性,如SOS和LOS在3 300~3 800 m、EOS在3 500~3 800 m因混交林的植被生长特性,出现SOS、EOS略微延迟、LOS缩短的非地带性差异.海拔在4 100 m以上区域以高山灌丛、草甸为主,因高山类植被生长主要受温度影响,并且随海拔升高,温度降低,呈现出SOS和EOS延迟、LOS缩短的总体趋势.除以上所提及的植被类型外,三江并流区存在许多其他植物群落,如常绿阔叶林、硬叶常绿阔叶林、干热性稀树灌木草丛、暖温性灌丛和灌草丛等,但群落面积占总面积百分比并不高,在此不进行详细分析.

图3 不同海拔上植被物候的变化情况
2.2 植被物候年际时空变化趋势
2.2.1 植被物候年际时间变化趋势
根据植被物候各参数逐年平均值计算得到三江并流区植被物候近19 年在时间上的年际变化趋势(见图4).其中SOS 呈提前趋势,平均提早1.71 d·(10 a)-1;而2010 年的SOS 出现大幅度延迟,该现象与研究区2009 年冬季开始至2010 年春季的极端干旱事件密切相关,即从2009 年冬季降水大幅度减少,2010年春旱现象严重加重,导致2010 年植被萌芽返青时间大幅度延迟.EOS 呈延迟趋势,平均延迟1.46 d·(10a)-1;LOS 呈延长趋势,平均延长3.17 d·(10 a)-1,而由于2010 年SOS 的延迟导致同年的LOS 出现大幅度缩短,即缩短至180 d 左右.
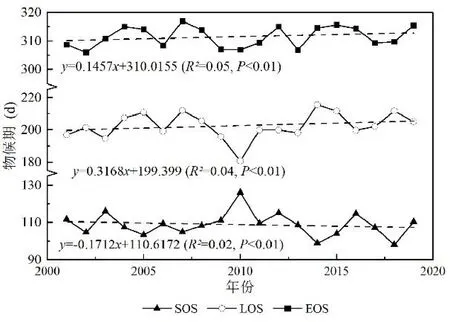
图4 2001-2019 年三江并流区植被物候年际变化趋势
2.2.2 植被物候年际空间变化趋势
为了研究三江并流区植被物候的年际空间变化趋势,对2001—2019 年内物候参数的栅格数据进行逐像元变化趋势分析,得到SOS、EOS 和LOS 的年际空间变化趋势(见图5).SOS 大部分地区呈现不显著提前趋势,而显著提前区域在河谷地带呈带状分布,显著延迟区域分布在东部边缘地区和部分高山地带.EOS 呈现不显著延迟趋势,不显著提前区域分布于低海拔河谷地带,其中还有零星点状分布的显著提前区域,大部分高山地带为延迟趋势.LOS 主要呈现为不显著延长趋势,在边缘地区、南部部分河谷地带、东部河谷地带,LOS 为缩短趋势,东部边缘地区为显著缩短趋势.

图5 2001—2019 年三江并流区植被物候年际变化空间分布
2.3 植被物候动态对气温和降水的响应
2.3.1 生长季始期对平均气温和降水的响应
从图6 可以看出,在3 月份,研究区北部高海拔地区的SOS 受温度和降水共同影响,且均为正向影响,其中降水比温度主控性更高;南部的低海拔与河谷区域受降水的负向影响更为显著,即降水增加,SOS 延迟.SOS 与4 月份的偏相关关系以降水的正向关系为主,但温度在高海拔区域依旧存在显著的正向影响.SOS 与5 月份的偏相关关系以温度的正向关系为主,在东南部低海拔河谷区域,SOS 与5 月份的温度和降水均存在显著的正向影响.虽然温度和降水在不同月份以及不同区域对SOS 的影响有所差异,但在高海拔地区,SOS 与温度一直存在显著的正相关性.
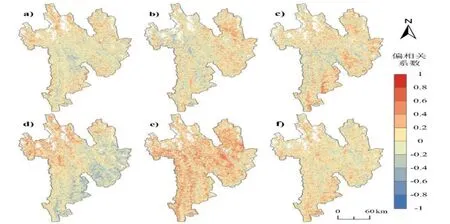
图6 三江并流区植被物候生长季始期与3—5 月份平均气温/降水的偏相关系数空间分布
2.3.2 生长季末期对平均气温和降水的响应
从图7 可以看出,EOS 与10 月份气温和降水的偏相关分析结果存在地域性差异,在北部、东部区域,气温和降水整体上均为正相关,其中气温的显著性高于降水;在南部、西部区域,降水对EOS 的显著性更强,且以负向影响偏多,而温度在高海拔区域以正向影响为主导.EOS 与11 月份的降水存在显著的正相关关系,同时温度主要在高海拔区域呈现为正向影响.EOS 在研究区东北部区域以及高海拔地区与12 月份气温的偏相关系数值主要为正值,虽然12 月份降水整体上为正相关关系,但是由于气象数据中12 月份的多年平均降水整体偏少,在此不予为考虑因素,但冬季降水对来年的SOS 变化具有一定的影响.
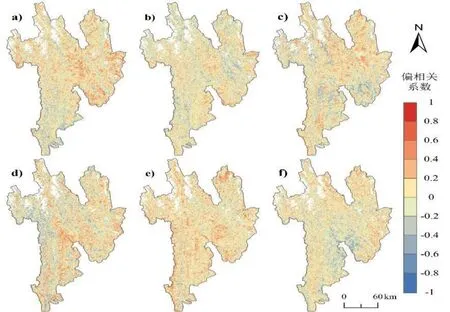
图7 三江并流区植被物候生长季末期与10—12 月份平均气温/降水的偏相关系数空间分布
2.3.3 生长季长度对平均气温和降水的响应
生长季长度主要由SOS 和EOS 决定,分别受到3—5 月份和10—12 月份的影响.研究区生长季集中在180—240 d,数据显示6—9 月份温度及降水的变化与生长季长度存在相关性,降水主要呈现一定的正相关性,而气温除7 月份为负相关外其余月份呈现正相关,与七月份基础气温较高,升温导致水分蒸散发增多导致的后序的干旱事件相关联.
从图8 可以看出,LOS 与6 月份的气温存在显著的正相关关系,与6 月份降水也以正相关关系为主,零星河谷区域存在负相关关系.而在7 月份,LOS 受降水的正向影响更为显著,7 月份温度整体上为负向影响.在8—9 月份,LOS 对气温、降水的响应存在明显的时空差异,8 月份降水的显著性高于气温,且以正向影响为主,但降水在部分高海拔地区的偏相关系数值为负值;9 月份气温的显著性高于降水,整体上为正相关关系,温度在部分高海拔地区为负向影响,而降水在北部、东部区域以正向影响为主,在南部区域则以负向影响为主.
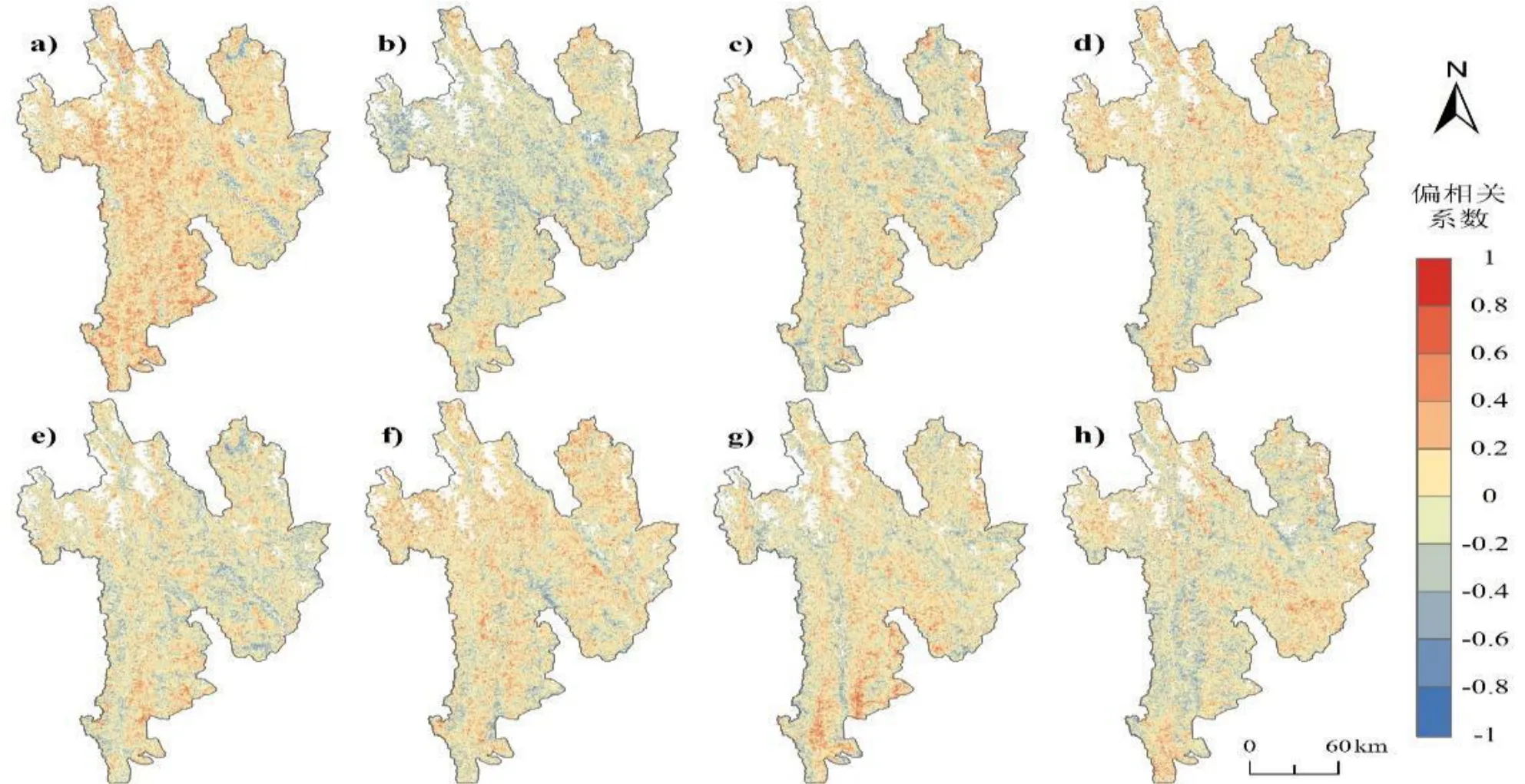
图8 三江并流区植被物候生长季长度与6—9 月份平均气温/降水的偏相关系数空间分布
3 讨论
在气候变暖的全球大环境下,三江并流区的植被物候也发生了相应的变化,植被物候呈生长季始期提前、生长季末期延迟、生长季长度延长的趋势与全球升温的趋势存在相关性[8].气候变化是多种地理要素因子的综合反映,而物候是对各种环境要素影响下以年为周期的自然现象,因地区差异,不同地区植被物候对不同气候因子的响应机制也不相同,仍存在许多植被物候研究的难点亟待深入探讨.
3.1 物候数据精度的提高
本文的MODIS EVI 数据虽然经过最大合成法在一定程度上消除云雾对EVI 的影响,但由于传感器的因素,仍存在一些难以处理的随机噪声.基于S-G 滤波的平滑去噪,在去噪过程中也不可避免将其中的一些细节去除,从而影响物候期提取的精度,因此S-G 滤波窗口大小和多项式阶数的取值需多次实验取最优,以实现保真性和去噪效果的最大化.同时16 d 的时间分辨率在对物候期的估算研究来说时间间隔较长,可以使用HANTS 方法对16 d 的EVI 数据重采样为8 d,甚至是1 d 的时间序列数据,来提高物候期提取的精度[38].
3.2 物候参数数值变化的主导因素
本文中的SOS、EOS 的主控因素在不同月份不尽相同,SOS 在3—4 月份受降水为主加气温的共同影响,而主控因素在5 月份转变为气温,主要原因是该地区冬春干旱的气候事件,3—4 月份的降水基本上是该地区的一轮降水,所以在此时段降水是主控因素.不过在高海拔地区,气温一直都是正向的主导因素,虽然在3—4 月份的主控性低于降水,但SOS 的大部分地区整体上在3—5 月份均受气温的正向影响,即气温升高,生长季始期提前,而生长季末期整体上受降水的主控因素为主,该结果与张晓东等在伏牛山森林植被物候结果一致[12].对于不同区域植被物候影响主控因子开展定量化的驱动力分析将有助于进一步理解植物物候的气候变化响应机制,在此基础上可以制定出更加合理的气候变化应对方案.三江并流区是地形起伏变化较大,干热河谷和高海拔并存的生态敏感区域,对于气候变化的过程可以进一步增加野外控制实验,获得不同气候变化情况下的植被生长状况,从而为保护区应对气候变化提供科学支撑.
3.3 极端气候事件对物候参数的影响
本文研究结果发现,2010 年SOS 出现大幅度延迟现象.经过查验19 年的逐月降水和逐月气温气象数据发现,2009 年冬季至2010 年春季出现极端干旱事件[39],2009 年10 月的降水量为27.45 mm,远远低于同月的多年平均降水67.47 mm.同时2010 年春季升温速度也高于多年平均值,春季升温快,导致区域潜在蒸散发的快速增加,更加剧了旱情,因此2010 年研究区受季风影响较大的西部、南部地区的SOS 集中在第120—140 d.
3.4 植被类型差异对植被物候结果的影响
因三江并流区气候复杂多样,植被类型非常丰富,不同植被对气候变化的响应机制有所差异,如文献[40]研究发现,草甸、沼泽植被生长季始期为延迟趋势,而草原、荒漠和高山植被的生长季始期为提前趋势.在本研究中也发现高山花卉等高山植物类的生长期集中在夏秋季,和整体上的植被物候期结果有时间上的差异.因此可以根据不同植被类型进行物候期参数提取,探讨研究不同植被类型物候的时空差异以及对气候变化的不同响应机制.
由于研究区海拔差异和植被差异巨大,物候对气候因素的变化极为敏感,比如研究区2009 年冬季的极端干旱导致2010 年物候生长季始期的大幅度延迟,而极端高温、极端低温事件对于植被物候的影响有待于进一步探究.此外,气候因素对植被的研究多存在一定的时滞性,不同植被类型的相应时间也存在一定差异,这个需要高分辨率物候数据的支持下,结合野外定点观测数据的支持下进行更加详细的讨论.
4 结论
本文以三江并流区为研究区,基于16 d 的MODIS EVI 数据、气象数据、土地利用数据和DEM 数据,利用S-G 滤波法重构时间序列,通过动态阈值法提取植被物候SOS、EOS 和LOS 等参数并转换为年序日,对2001—2019 年空间分布格局和长时序列变化趋势进行分析,并探讨植被物候变化与气温、降水的相关性.主要结论如下:
(1)三江并流区植被物候的SOS 集中在第80—140 d,呈现由东部向西部延迟、河谷地带早高山地带晚的规律.EOS 集中在第290—330 d,呈现东部早西部晚的规律.LOS 集中在180—240 d.SOS、EOS 和LOS分别呈显著提前、显著延迟、显著延长趋势,分别平均提前1.71 d·(10a)-1、延迟1.46 d·(10a)-1、延长3.17 d·(10a)-1.
(2)SOS 在3—4 月份以降水为主导响应因素,但3 月份降水的正负向影响存在空间差异,北部高海拔地区受降水的正向影响为主,而南部的低海拔与河谷区域受降水的负向影响更为显著;SOS 与4 月份降水主要呈正相关,与3—4 月份的气温整体上以正相关为主,同时生长季始期与5 月份的气温和降水主要呈正相关,其中温度的正向影响占主导.
(3)EOS 与10 月份气温和降水的偏相关分析结果存在地域性差异,在北部、东部区域以气温的正向影响为主,在南部、西部区域以降水的负向影响为主;EOS 与11—12 月份的气温和降水均呈正相关,11 月份的降水的正相关性更高,12 月份则以气温的正向影响为主,12 月份降水因数值偏低,不纳入考虑因素.
(4)LOS 与6 月份气温和7 月份降水存在显著的正相关关系,同时6 月份降水以正相关为主,7 月份温度整体上为负向影响;LOS 与8—9 月份的气温和降水均呈正相关,但8 月份降水正向影响高于气温,9 月份气温正向影响高于降水.
猜你喜欢
军事文摘(2023年20期)2023-10-31
成都信息工程大学学报(2022年3期)2022-07-21
热带作物学报(2022年6期)2022-07-08
小哥白尼(野生动物)(2022年3期)2022-06-16
今日农业(2021年2期)2021-03-19
民族音乐(2018年1期)2018-04-18
西南农业学报(2017年5期)2017-06-23
河北林业科技(2016年5期)2016-11-08
西南农业学报(2016年5期)2016-05-17
小雪花·成长指南(2015年10期)2015-10-23
