
腐殖酸营养液对镉胁迫下树仔菜幼苗生理生化特性及重金属积累的影响
2024-01-08 09:34酒元达叶海辉邹冬梅苏初连
中国瓜菜 2023年12期
范 琼,酒元达,叶海辉,冯 剑,邹冬梅,苏初连,赵 敏
(中国热带农业科学院分析测试中心·海南省热带果蔬产品质量安全重点实验室·农业农村部亚热带果品蔬菜质量安全控制重点实验室 海口 571101)
镉(Cd)是环境中常见的重金属元素之一,2014年4 月国家环境保护部和国土资源部联合发布的《全国土壤污染状况调查公报》显示,我国耕地土壤重金属总超标率为16.1%,其中重金属Cd 点位超标率为7.0%[1]。海南蔬菜产地土壤多为酸性,Cd 离子活化性较高[2],易被蔬菜吸收,Cd 在蔬菜内累积后会干扰正常的细胞功能和代谢,诱导植株根和叶的生理生化变化,而且还可经生物富集作用,通过食物链威胁人体健康[3]。因此,Cd 是海南蔬菜安全生产中的重要风险控制指标。
目前,施用含腐殖酸类物质的有机物料被认为是降低重金属迁移率的有效办法[4]。腐殖酸是一种高分子有机混合物,含有丰富的羧基、酚羟基、羰基和甲氧基等活性含氧官能团[5],通过与Cd2+发生吸附、螯合和络合作用,影响Cd2+的迁移和生物有效性[6]。在前期研究中,赵敏等[7]通过施用碱性腐殖酸水溶肥有效提高土壤pH,降低土壤和树仔菜的Cd含量。但也有一些研究报道认为腐殖酸对土壤重金属有活化作用,柏宏成[8]的研究表明,低浓度腐殖酸可以有效淋溶土壤Cd,活化土壤总Cd;邹传等[9]研究表明,可溶性腐殖酸可以活化和解吸Cd,腐殖酸滤渣则能吸附Cd,因此,腐殖酸的种类和环境因素等都会影响对重金属活化或钝化的作用。
树仔菜(Sauropus androgynus)属大戟科守宫木属植物,又名守宫木、天绿香、泰国枸杞、五指山野菜等,是最具特色的海南野菜之一[10]。笔者前期研究发现,树仔菜是一种易超量吸收Cd 并将其向上迁移的蔬菜[11]。但在Cd 污染土壤中树仔菜的重金属积累的生理特性还有待进一步研究,腐殖酸类营养液在高Cd 胁迫下对树仔菜的重金属积累和生理生化的影响还未见报道。
笔者通过Cd 胁迫盆栽试验,研究腐殖酸类营养液对不同Cd 浓度下树仔菜生长指标、叶绿素含量、生理生化活性的影响以及重金属含量的变化规律,为明晰腐殖酸类营养液对树仔菜在高Cd 胁迫下积累和生理生化影响的作用机制,寻找适合树仔菜种植在Cd 污染土壤中的农艺强化方法提供基础数据。
1 材料与方法
1.1 材料
供试土壤:盆栽用土壤采自海南省五指山市畅好乡树仔菜基地的表层土壤(0~20 cm),土壤类型为砂红壤土。土壤采回后经风干、研磨、过筛(2 mm)后保存。
供试材料:海南省五指山市广泛栽培的马来西亚种树仔菜由五指山市汇通农业综合开发有限公司提供。选择苗龄40 d 左右,大小均匀、无病虫害、无损伤的幼苗进行盆栽试验。
供试肥料:腐殖酸营养液由中国热带农业科学院分析测试中心提供,其中黄腐殖酸含量(w,后同)50%;复合肥料为雅苒国际有限公司生产,N、P、K质量比为15∶15∶15;蚯蚓粪购自海南星农夫生态科技有限公司,有机质含量为50.1%。
1.2 方法
1.2.1 试验设计 试验于2020 年8—9 月在海南省海口市中国热带农业科学院温室大棚内进行。选用体积比为1∶1 的砂红壤土和椰糠混合土样作为盆栽土壤,每盆装入1.5 kg 土壤,盆栽土基本理化性质如下:pH 5.2,有机质含量73.8 g·kg-1,阳离子交换量9.2 cmol+·kg-1,碱解氮含量103.3 mg·kg-1,速效钾含量1.33 g·kg-1,有效磷含量59.4 mg·kg-1,交换性钙含量20.7 cmol·kg-1,交换性镁含量1.90 cmol·kg-1,Zn 含量42.9 mg·kg-1,未有Cd 检出。
重金属Cd 以CdCl2水溶液形式,采用喷雾方式以4 个浓度(w,后同)梯度均匀喷洒至土壤中,分别为0(Cd0)、15(Cd15)、25(Cd25)、45 mg · kg-1(Cd45),计算配置相应浓度所需试剂质量和土壤60%田间持水量所需水量,用扎有细孔的保鲜膜封住盆口,在室温下老化2 周,其间采用称质量法补充丢失的水分。为保证土壤具有丰富的有机质,老化平衡后施入底肥和10%的蚯蚓粪,1 周后移栽树仔菜苗,每盆种植1 株,每个处理3 盆,试验3 次重复。试验分为试验区和对照区,试验区称取0.125 g碱性腐殖酸营养液溶于50 mL 清水后对每株进行灌根施肥,每10 d 施用1 次,对照区则不施用腐殖酸营养液,其他田间管理措施保持一致。种植4 个月后收获,采集的土壤样品经自然风干、磨细,分别过60 目和100 目筛,保存待测;植物样品洗净后,测定生物量,并分成根、叶两部分分别测定酶活性。
1.2.2 Cd 胁迫下树仔菜生长指标测定 采用直尺测定树仔菜株高;吸干树仔菜叶片和根部的水分,用电子天平直接测量地上部和根部的鲜质量;将树仔菜地上部和根部清洗干净后放入烘箱内105 ℃杀青30 min 后,在70 ℃下干燥72 h 至恒质量,用电子天平测量地上部和根部的干质量;采用长宽乘积法测定叶面积,叶长用直尺测量树仔菜展开叶尖至叶基的距离,叶宽为直尺测量树仔菜展开叶最宽部分的距离。
1.2.3 Cd 胁迫下树仔菜生理指标的测定 在树仔菜收获时将根和叶分开,装入塑封袋内,于-80 ℃超低温冰箱保存,备用。按照苏州科铭生物技术有限公司提供的试剂盒说明书,测定以下指标。采用分光光度法测定叶绿素含量;采用TBA 法测定丙二醛(MDA)含量,采用羟胺法测定超氧化物歧化酶(SOD)活性、采用比色法测定过氧化物酶(POD)活性、采用钼酸铵比色法测定过氧化氢酶(CAT)活性。
1.2.4 Cd 胁迫下树仔菜Cd 和Zn 含量的测定 称取0.25 g 树仔菜干样加入8 mL 硝酸放入消解炉中消解,定容至25 mL,采用电感耦合等离子体质谱仪测定树仔菜Cd 和Zn 含量[11]。
1.3 数据分析
采用Excel 2016、Origin 2018 制图,采用SPSS 22.0 软件进行试验数据的差异显著性分析和相关性分析。
2 结果与分析
2.1 腐殖酸营养液对Cd胁迫下树仔菜生长的影响
由表1 可知,在CK 处理中,随着Cd 胁迫浓度的增加,树仔菜叶面积、叶片数和株高逐渐下降。其中,Cd45 处理的树仔菜叶面积和株高分别比Cd0处理显著降低22.2%和16.2%,而叶片数在不同处理间差异不显著。树仔菜地上部干质量和总干质量呈先降低后升高的变化趋势,在Cd0~Cd25 处理下,树仔菜地上部干质量和总干质量均显著下降,Cd45 处理树仔菜总干质量比Cd15 和Cd25 处理分别增加5.2%和13.3%。
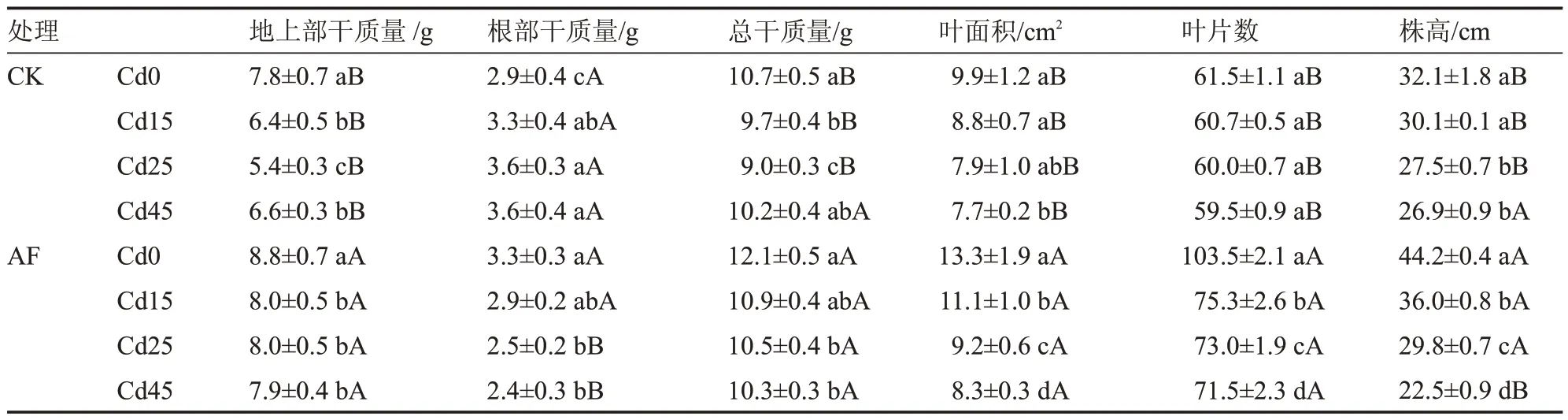
表1 腐殖酸营养液在Cd 胁迫下对树仔菜生长的影响Table 1 Effects of AF on growth of Sauropus androgynus under Cd stress
在AF 处理中,随着土壤Cd 浓度增加,树仔菜地上部干质量、根部干质量、总干质量、叶面积、叶片数、株高均呈下降趋势。其中,Cd45 处理的树仔菜总干质量、叶面积、叶片数和株高比Cd0 处理分别显著下降14.9%、37.6%、30.9%和49.1%。
与CK 处理相比,在Cd0~Cd45 处理下,AF 处理的树仔菜地上部干质量、叶面积和叶片数均显著高于CK 处理。AF 处理的树仔菜总干质量和株高在Cd0~Cd25 处理下显著高于CK 处理12.4%~16.7%和8.4%~37.7%。在Cd 浓度为45 mg·kg-1时,AF 处理与CK 处理的树仔菜总干质量差异不显著,AF 处理的株高显著低于CK 处理16.4%。这表明了高Cd 土壤下树仔菜在一定的时间和浓度范围内可以正常生长,施用腐殖酸营养液可促进树仔菜生长,但随着Cd 浓度的增加,树仔菜生长程度逐渐下降,尤其是在Cd 浓度达到45 mg·kg-1时,树仔菜株高受到显著抑制。
2.2 腐殖酸营养液对Cd胁迫下树仔菜根部和叶片Cd和Zn含量的影响
Cd 胁迫下树仔菜根部和叶片Cd 和Zn 含量变化见图1。由图1-A 可知,随着Cd 胁迫浓度的增加,CK 处理和AF 处理的树仔菜根部Cd 含量呈增加趋势。CK 处理下树仔菜根部Cd 含量分别为0.61、322.8、587.9、1 000.7 mg·kg-1;AF 处理下树仔菜根部Cd 含量分别为0.69、382.6、645.3、1 390.9 mg·kg-1。在Cd0~Cd25 处理下,两处理的树仔菜根部Cd 含量差异不显著,在Cd 浓度为45 mg·kg-1时,AF 处理的根部Cd 含量显著高于CK 处理39.0%。从Cd胁迫下树仔菜叶片Cd 含量变化可以看出,随着Cd浓度的增加,CK 处理的树仔菜叶片Cd 含量呈先升高后降低的变化趋势,AF 处理的树仔菜叶片Cd 含量则呈上升趋势。在Cd 浓度为45 mg·kg-1时,CK处理和AF 处理的树仔菜叶片Cd 含量分别为7.64和17.93 mg·kg-1,说明在正常农艺措施的情况下,土壤Cd 浓度到达一定值后树仔菜叶片Cd 含量会下降,但腐殖酸营养液施用后会促进Cd 在树仔菜根部和叶片中富集。

图1 Cd 胁迫下腐殖酸营养液对树仔菜根部和叶片Cd和Zn 含量的影响Fig.1 Effects of AF on the contents of Cd and Zn in roots and leaves of Sauropus androgynus under Cd stress
由图1-B 可知,随着土壤Cd 浓度增加,CK 处理的树仔菜根部Zn 含量均显著下降,AF 处理的树仔菜根部Zn 含量呈降-升-降的变化趋势,CK 处理的树仔菜根部Zn 含量均显著高于AF 处理。随着土壤Cd 浓度增加,CK 处理的树仔菜叶片的Zn 含量不断下降,而AF 处理中的树仔菜叶片的Zn含量逐渐升高。在Cd0~Cd25 处理下,AF 处理的树仔菜叶片Zn 含量均显著低于CK 处理,在Cd浓度为45 mg·kg-1时,AF 处理的树仔菜叶片Zn 含量显著高于CK 处理36.9%。说明在高浓度Cd 胁迫下,腐殖酸营养液可以促进Zn 在树仔菜叶片中富集。
2.3 腐殖酸营养液对Cd胁迫下树仔菜叶绿素含量的影响
由图2 可知,随着土壤Cd 胁迫浓度的升高,CK 处理中树仔菜叶绿素a 呈现先上升后下降的变化趋势,叶绿素b 含量呈先上升后下降再上升的变化趋势,AF 处理中树仔菜叶绿素a 和叶绿素b 含量均呈先下降后上升的变化趋势。在没有Cd 胁迫时,AF 处理的树仔菜叶绿素a 和叶绿素b 含量分别显著高于CK 处理77.8%和71.8%;在Cd 浓度为15 mg·kg-1时,AF 处理的树仔菜叶绿素a 和叶绿素b含量急剧下降,其中叶绿素a 含量显著低于CK 处理;在Cd 浓度为45 mg·kg-1时,两个处理的树仔菜叶绿素a 和叶绿素b 含量差异不显著。通过对叶绿素a/b 的计算(图2-C),发现CK 处理和AF 处理的叶绿素a/b 在Cd25 处达到最高,说明在Cd浓度为25 mg·kg-1时,树仔菜对Cd 的敏感度最强。
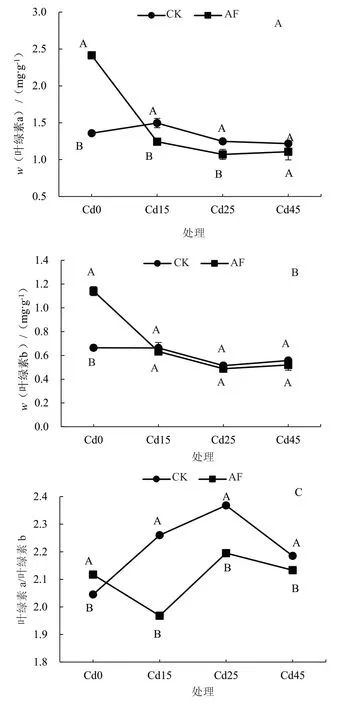
图2 Cd 胁迫下腐殖酸营养液对树仔菜叶绿素a、叶绿素b含量和叶绿素a/b 的影响Fig.2 Effects of AF on chlorophyll a,chlorophyll b content and chlorophyll a/b of Sauropus androgynus under Cd stress
2.4 腐殖酸营养液对Cd胁迫下树仔菜根部和叶片丙二醛含量的影响
由图3-A 可知,随着Cd 浓度升高,CK 处理的根部MDA 含量呈先升高后降低的变化趋势,AF 处理的根部MDA 含量呈逐渐升高趋势。在Cd0~Cd25 处 理 下,CK 处 理 和AF 处 理 的 根 部MDA 含量差异不显著;在Cd 浓度为45 mg·kg-1时,AF 处理的根部MDA 含量比CK 处理显著提高19.6%。从图3-B 可知,随着Cd 浓度的升高,CK 处理和AF 处理的叶片MDA 含量均呈先升高后降低的变化趋势。在Cd 浓度为25 mg·kg-1时,叶片MDA 含量最高。在Cd15~Cd45 处理下,AF 处理的叶片MDA 含量分别比CK 处理显著提高10.1%、10.8%和7.0%,且AF 处理和CK 处理叶片的MDA 含量均高于根部。说明随着土壤Cd 浓度增加,AF 处理中的根部膜脂过氧化程度逐渐增加,树仔菜有特殊的运输渠道将土壤中的Cd 运输到叶部,对叶片细胞质膜造成损伤,从而降低对根部的伤害程度,但是当Cd 浓度达到一定程度后,叶片细胞质膜损伤程度逐渐下降。

图3 Cd 胁迫下腐殖酸营养液对树仔菜根部和叶片的MDA 的影响Fig.3 Effects of AF on MDA of roots and leaves of Sauropus Androgynus under Cd stress
2.5 腐殖酸营养液对Cd胁迫下树仔菜根部和叶片抗氧化酶活性的影响
由图4 可知,在Cd 胁迫下树仔菜叶片和根部CAT、SOD、POD 活性的响应不同。从图4-A~B 可知,随着Cd 胁迫浓度的增加,AF 处理的树仔菜根部CAT 活性随着Cd 胁迫浓度的增加而提高,在Cd0 和Cd15 处理下,AF 与CK 处理差异不显著;在Cd25 和Cd45 处理下,AF 处理的树仔菜根部CAT 活性分别显著高于CK 处理44.5%和93.4%。AF 处理与CK 处理的叶片CAT 活性随着Cd 浓度增加均呈先升高后降低的变化趋势,在Cd浓度为15 mg·kg-1时,CAT 活性最高,AF 处理的叶片CAT 活性在Cd0、Cd15、Cd25、Cd45 处理下分别显著高于CK 处理101.5%、218.8%、309.1%和317.4%。
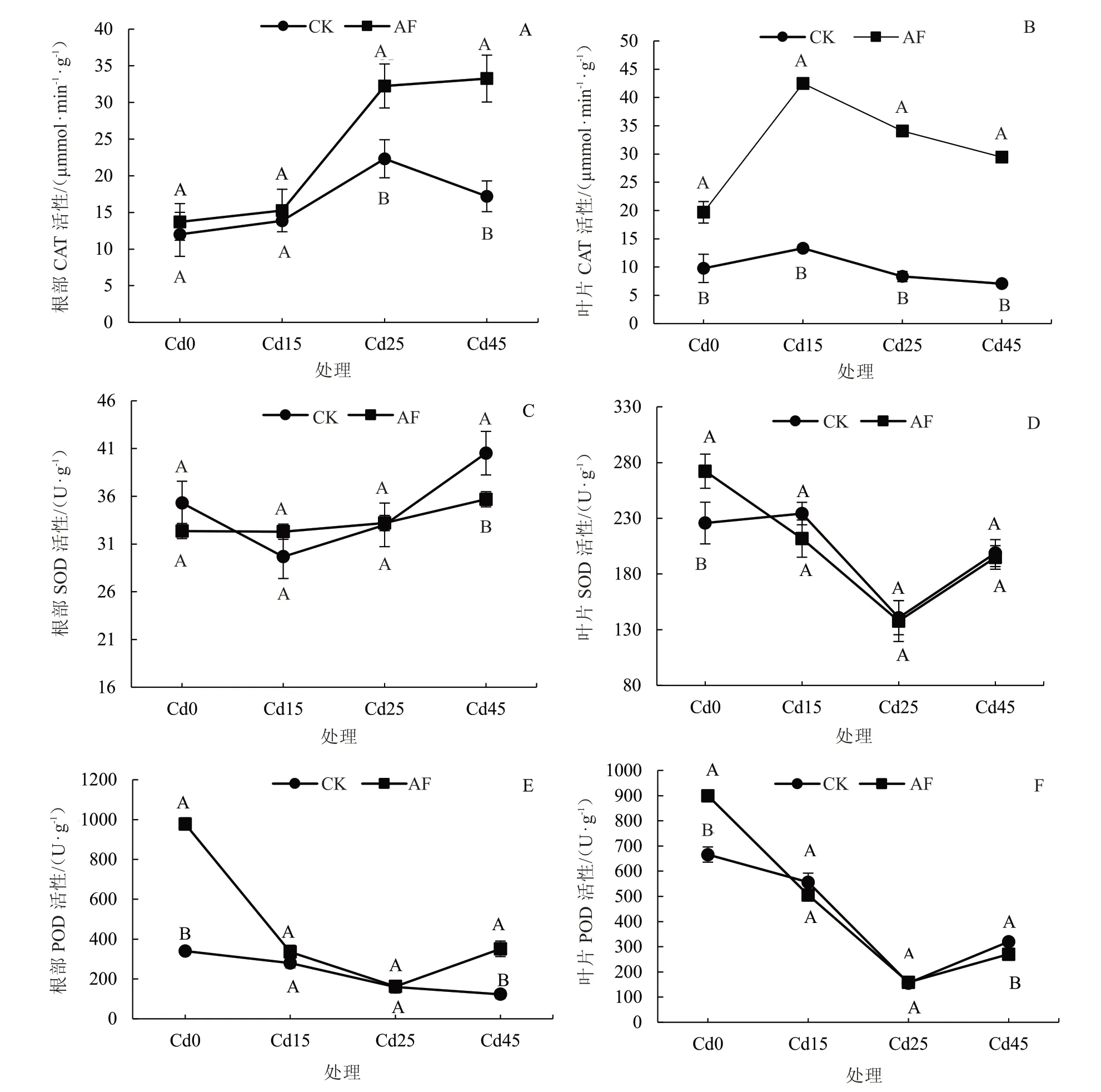
图4 Cd 胁迫下腐殖酸营养液对树仔菜根部和叶片的CAT、SOD、POD 活性的影响Fig.4 Effects of AF on CAT,SOD,POD activity of roots and leaves of Sauropus androgynus under Cd stress
从图4-C~D 可知,Cd 胁迫处理的树仔菜叶片中的SOD 活性大于根。随着土壤Cd 浓度增加,AF处理的根部SOD 活性逐渐升高,在Cd0~Cd25 处理下,CK 处理和AF 处理的根部SOD 活性没有显著差异,在Cd 浓度为45 mg·kg-1时,CK 处理的根部SOD 活性显著高于AF 处理。随着土壤Cd 浓度增加,AF 处理的叶片SOD 活性则呈现先降低后升高的变化趋势,在没有Cd 胁迫时,AF 处理的叶片SOD 活性显著高于CK 处理20.6%,在Cd15~Cd45处理下,AF 与CK 处理差异不显著。
从图4-E~F 可知,随着土壤Cd 浓度的升高,CK 处理中根部POD 活性逐渐下降,AF 处理的POD 活性则呈现先降低后升高的变化趋势。在Cd0 和Cd45 处理下,AF 处理的根部POD 活性分别显著高于CK 处理187.6%和186.4%。随着土壤Cd 浓度的升高,树仔菜叶片POD 活性的变化呈先降低后升高的变化趋势。在没有Cd 胁迫时,AF 处理的树仔菜叶片POD 活性显著高于CK 处理35.1%。
2.6 相关性分析
从表2 可知,树仔菜株高与根部Cd 含量呈显著负相关,与叶面积、叶绿素b 含量呈极显著正相关,与总干质量、叶绿素a 含量、叶片POD 和根部POD 活性呈显著正相关;树仔菜叶片Cd 含量与根部Cd 含量、根部CAT 活性呈显著正相关,与叶片MDA、根部MDA 含量呈极显著正相关,与根部Zn含量、叶片POD 活性呈显著负相关;树仔菜根部Cd含量与根部MDA 含量、根部CAT 活性呈显著正相关,与根部Zn 含量、株高、叶片POD 活性呈显著负相关;树仔菜根部Zn 含量与叶片Cd 含量、根部Cd含量和叶片MDA 含量呈显著负相关;叶绿素a 含量与叶绿素b 含量、根和叶片POD 活性呈极显著正相关,与株高、叶面积呈显著正相关,与叶片MDA 含量呈显著负相关;叶绿素b 含量与株高、叶面积、根部和叶片POD 活性呈极显著正相关,与叶片SOD 活性、总干质量呈显著正相关,与叶片MDA 含量呈显著负相关;叶绿素a/b 与总干质量呈显著负相关。
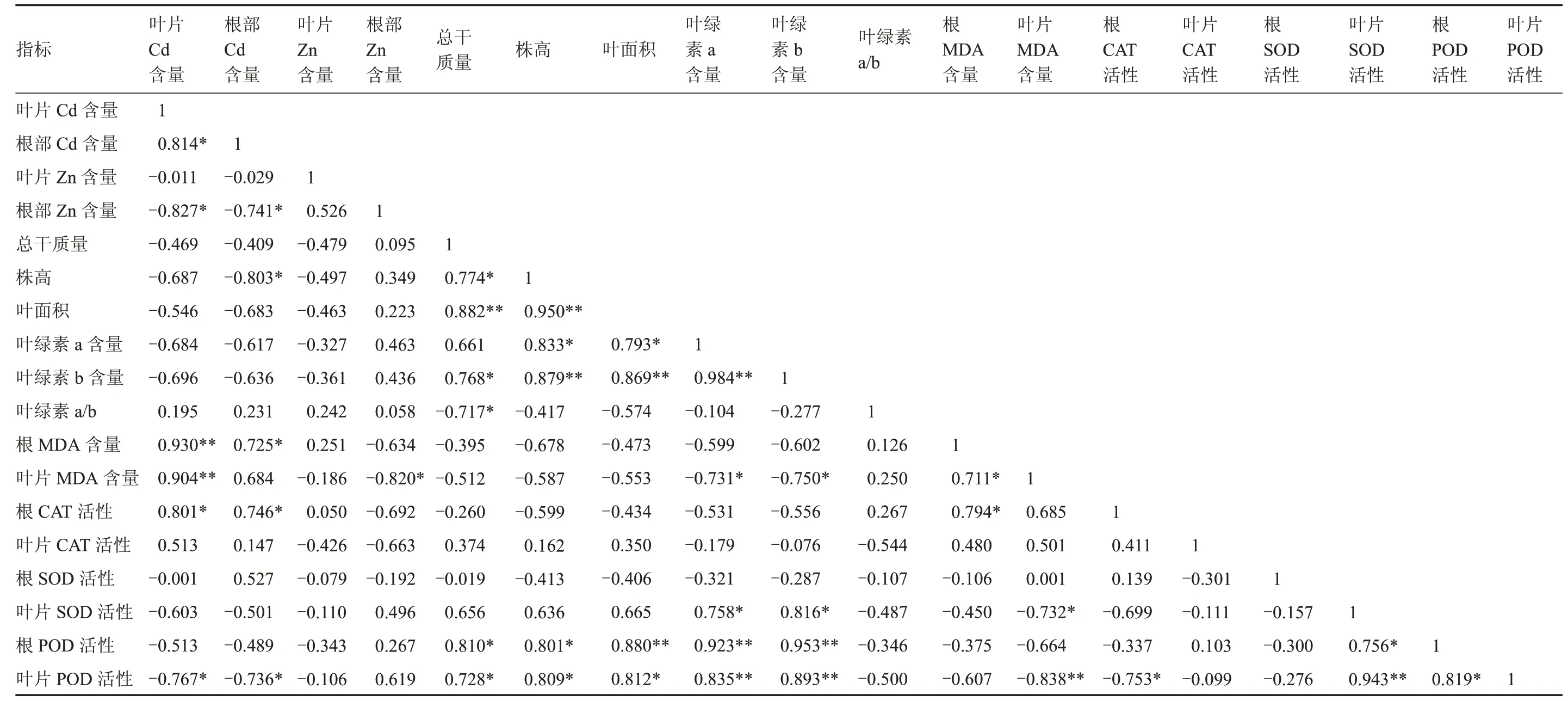
表2 树仔菜重金属含量和生理生化特性的相关性分析Table 2 Correlation analysis of heavy metal content and physio-biochemistry characteristic in Sauropus androgynus
3 讨论与结论
3.1 腐殖酸营养液对Cd胁迫下树仔菜幼苗生长的影响
Liu 等[12]研究表明,腐殖酸肥料可以提高土壤养分有效性和玉米养分吸收效率,严萍[13]研究证明,腐殖酸对小麦、水稻幼苗的株高、鲜干质量、根系活力都有一定的促进作用。在笔者的研究中,Cd 胁迫下施用腐殖酸营养液可促进树仔菜生长发育,在土壤Cd 浓度为0~25 mg·kg-1时,施用腐殖酸营养液的树仔菜幼苗总干质量、株高、叶面积和叶片数均高于对照组。但是在土壤Cd 浓度为45 mg·kg-1时,虽然AF 处理显著促进了Cd 在树仔菜根部的积累,树仔菜叶面积和叶片数均高于CK 处理,然而株高却显著低于CK 处理,树仔菜株高与根部Cd 含量呈显著负相关,说明了Cd 在树仔菜根部的积累量可能达到上限,导致树仔菜生长受到了抑制,主要是体现在株高上,但是并未出现树仔菜死亡的现象。
3.2 腐殖酸营养液对Cd 胁迫下树仔菜幼苗Cd 和Zn含量的影响
当Cd 胁迫浓度达到45 mg·kg-1时,CK 处理的树仔菜叶片的Cd 和Zn 含量显著降低,AF 处理后树仔菜的根部和叶片的Cd 含量以及叶片的Zn 含量均高于CK 处理,这可能是树仔菜在正常种植下土壤Cd 胁迫达到一定浓度后,根部代谢受损,向叶片迁移的速率下降,而施用腐殖酸营养液促进了树仔菜对土壤中Cd 和Zn 含量的吸收。王凯迪[14]研究表明,施用腐殖酸对土壤Cd 具有一定的活化作用。吴洪燕等[5]发现,低分子腐殖酸对土壤重金属的吸附络合能力更强,能提高Cd 的生物有效性。在本研究中,笔者施用的腐殖酸营养液主要成分为黄腐酸(含量≥50%),黄腐酸是一种易溶于酸碱的腐殖质,易和重金属离子形成易溶的络合物[15]。因此,在笔者的试验中,施用腐殖酸营养液显著提高了树仔菜Cd 和叶片Zn 的积累量,可能存在至少两方面的原因,一是腐殖酸营养液提高了土壤重金属的生物有效性,土壤Cd 和Zn 更容易被树仔菜吸收和迁移;二是腐殖酸营养液促进了树仔菜生物量提高,进而提高树仔菜对重金属的积累量。由于树仔菜以食用嫩梢(嫩茎10 cm)为主,一般种植6 个月后方可采摘嫩茎供食用,笔者的试验材料为生长2个月左右树仔菜幼苗,嫩梢尚不明显,因此,在Cd胁迫下腐殖酸营养液是否促进嫩梢对Cd 积累还有待进一步研究。
3.3 腐殖酸营养液对Cd胁迫下树仔菜幼苗叶绿素含量的影响
在笔者的试验中,没有Cd 胁迫时,施用腐殖酸营养液的树仔菜叶绿素a 和叶绿素b 含量均高于CK 处理组,但是Cd 浓度为15 mg·kg-1时,AF 处理的叶绿素a 和叶绿素b 含量急剧下降。叶绿素a/b被认为是衡量植物叶片感受重金属相对敏感的一个指标[16],在Cd15~Cd45 处理下,AF 组的叶绿素a/b 显著低于CK 组。叶绿素a 和叶绿素b 含量与叶片MDA 含量呈显著负相关,与叶片SOD 活性呈显著正相关,与叶片POD 活性呈极显著正相关,说明Cd 胁迫下树仔菜幼苗叶绿素的变化会使MDA 含量和酶活性等生理生化发生相应变化。朱润华等[17]的研究表明,随着Cd 胁迫浓度增加,越来越多的重金属沉积在细胞膜表面,破坏叶绿体的结构与功能,导致叶绿素降解。张大为等[18]的研究表明,高浓度的Cd 胁迫会显著抑制油菜的叶绿素含量,导致叶片黄化,与本研究结果一致。
3.4 腐殖酸营养液对Cd胁迫下树仔菜生理生化特性的影响
从生理生化的角度来看,腐殖酸营养液可以缓解树仔菜的氧化胁迫。MDA 是膜脂过氧化的最终分解产物,浓度越高,表明膜脂过氧化程度越高,细胞膜损伤越严重[16]。在笔者的研究中,在CK 组中,根部和叶片MDA 含量呈先升高后降低的变化趋势,说明树仔菜在正常种植下,胁迫程度超过树仔菜自身调节作用的极限后,相关渗透调节物质含量下降。施用腐殖酸营养液的树仔菜根部MDA 含量随着根部Cd 含量的增加而逐渐升高,这可能与AF处理促进树仔菜根部和叶片Cd 积累有关,树仔菜根部和叶片细胞膜受伤害的程度更加严重。过氧化氢酶、超氧化物歧化酶和过氧化物酶是抗氧化酶系统中控制植物体内活性氧积累的最主要的酶[17]。随着Cd 浓度增加,AF 处理的根部CAT 和SOD 活性不断升高,在Cd 浓度为45 mg·kg-1时,根部CAT和POD 活性均显著高于CK 处理,说明树仔菜受到Cd 胁迫时,整个酶保护系统起到了一定的缓解自由基危害的作用,从而将胁迫产生的活性氧维持在平衡水平。Garcia 等[20]研究表明,腐殖酸与植物的根系统存在互相作用,影响叶片和根部抗氧化防御系统中CAT 和SOD 活性,与本研究结果一致。此外,在笔者的研究中,叶片较根部变化趋势明显可能是由于叶片是植物主要的光合作用部位,酶活性变化较为敏感。这也说明了Cd 胁迫下树仔菜体内抗氧化系统消除Cd 诱导的ROS 能力没有降低,且在叶片中尚有所增强,因此树仔菜对Cd 的解毒能力较强,这应是其能富集Cd 的原因之一。
综上所述,在Cd 胁迫下腐殖酸营养液促进了树仔菜重金属的积累,并通过激发抗氧化酶保护系统来抵抗树仔菜中ROS 积累,在Cd 浓度为0~25 mg·kg-1范围内,腐殖酸营养液在促进树仔菜重金属积累的同时促进幼苗生长,为树仔菜在高Cd土壤中种植进行土壤生物修复提供了新的思路。
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-04
载人航天(2021年5期)2021-11-20
金属加工(热加工)(2020年12期)2020-02-06
猪业科学(2018年4期)2018-05-19
电子制作(2017年20期)2017-04-26
山东林业科技(2016年5期)2016-07-05
腹腔镜外科杂志(2016年11期)2016-06-01
浙江农业科学(2016年11期)2016-05-04
磁共振成像(2015年9期)2015-12-26
实用手外科杂志(2015年2期)2015-08-28
