
禾草Epichloё属内生真菌分类学研究进展
2024-01-06 09:11杜明祥李春杰陈泰祥
草地学报 2023年12期
杜明祥,王 添,李春杰,陈泰祥
(草种创新与草地农业生态系统全国重点实验室/兰州大学草地农业科技学院,甘肃 兰州 730020)
禾草内生真菌是在宿主禾草的组织间隙完成其全部或大部分生命周期,但不引起宿主禾草产生外部明显症状的一类内生真菌[1-2],其分类学地位属于子囊菌门(Ascomycota)、肉座菌目(Hypocreales)、麦角菌科(Clavicipitaceae)的香柱菌属(Epichloё)[3-6]。1898年,德国学者Vogl[1]从毒麦(Loliumtemulentum)种子中分离出了Epichloё属禾草内生真菌,这是全世界首次发现的禾草内生真菌。这类真菌主要分布在宿主禾草的地上部分,包括叶鞘、茎秆,也有少部分分布于宿主叶片表面[3,7],并与宿主禾草经历了长时间的协同进化[8],形成了十分稳定的互利共生关系[9],其侵入禾草既不会破坏宿主禾草的细胞结构,也不会同病原菌一样入侵宿主细胞对宿主的生长发育产生负面影响,其主要从宿主细胞间隙吸收生长所需的营养,与宿主交换碳、氮、磷等多种营养元素,产生多种次生代谢产物增强宿主的免疫防御能力,保护其免受各种生物和非生物胁迫的伤害[10-13]。
Epichloё属禾草内生真菌是目前研究最广泛的一类内生真菌[14],其一方面可为宿主植物带来诸多增益作用,另一方面,部分共生体产生的麦角类和吲哚二萜类生物碱可引致草食家畜中毒,影响草地畜牧业的健康发展,这一两面性使得其在全世界范围内广泛引起了农学家、畜牧学家、植物病理学家、植物生理学家以及分子生物学家等的兴趣[2]。目前已在全世界80属300多种禾草中发现了Epichloё属内生真菌[15],所鉴定的内生真菌种类多样,且其分类地位也随着研究的深入不断发生变化。本文旨在对禾草内生真菌的命名历史进行系统归纳,详尽描述目前已经报道的禾草内生真菌种类多样性和形态学多样性,并初步总结我国禾草内生真菌种类多样性,以期为未来新禾草内生真菌资源的分类提供参考依据,并为未来我国优良禾草内生真菌资源的挖掘与利用提供基础数据。
1 禾草内生真菌的命名历史
最初,禾草内生真菌的鉴定主要依靠以分生孢子形态差异和培养特性为基础的形态学特征,随着分子生物学的发展与进步,越来越多的分子生物学技术被广泛应用于禾草内生真菌的分类学研究[16]。目前,形态学特征、宿主特异性、系统发育学以及生物碱多样性等方法已广泛应用于禾草内生真菌分类学鉴定。系统发育学主要利用物种的遗传信息--DNA序列来构建系统发育关系[17],确定新种与已知种的亲缘关系,从而确定其分类地位。系统发育学表明,Epichloё属内生真菌的起源有三种,一种是直接来源于某一种内生真菌,一种是有性型之间的种间杂交,还有一种是有性型和无性型的种间杂交,通过种间杂交重组,产生了新的遗传性状,从而出现了新的种、变种和亚种[18]。这些内生真菌在植物中的传播具有两种不同方式,一种是在禾草种子萌发时,种皮和胚乳中的有性型内生真菌会侵入禾草茎秆和叶鞘部位的细胞间隙[19],接着在部分分蘖的叶鞘上产生带有分生孢子的子座[20-21],植种蝇属(Botanophila)昆虫携带这些分生孢子传播到周围植物上,这一传播方式即为内生真菌的水平传播[22-25]。另一种是有性型内生真菌通过种间杂交产生无性型内生真菌,无性型内生真菌随着宿主的分蘖和抽穗传播至种子中,这种通过种子从亲代向子代传播的方式,称为垂直传播,也是无性型内生真菌唯一的传播方式[26-27]。
Epichloё属最初为一个亚属,而Epichloёtyphina在1849年归入其中并成为该属的模式种,在1865年,Epichloё亚属被提升为属水平。之后经多年的研究,Epichloё属的更多的成员得以被发现。与此同时,对于Epichloё属无性型的分类和命名一直饱受争议。在历经多次命名后,Morgan等[28]在Acremonium内部构建了一个属:Albolanosa,并将Epichloё属无性型归于其中。但相关学者发现这一分类方式并不合理,又利用18SrDNA的系统发育学信息将属于Epichloё属无性型的8个有效种独立出来,构建了仅包含禾本科植物内生真菌的新属:Neotyphodium[29]。随后,根据分子系统发育学、宿主兼容性、宿主范围、形态特征以及依据无性型菌株与Epichloё属各个种的遗传进化关系等,世界各地又陆续报道了多个有性型Epichloё属及其无性型Neotyphodium属内生真菌[30-34]。
在经典分类学中,依据形态特征,Neotyphodium属内生真菌被认为是Epichloё属的无性世代[35]。学者们利用系统发育学数据,尤其是根据β-微管蛋白(tubB)及翻译延长因子1-α(tefA)两对管家基因的分析,发现了越来越多的新种,但这些研究结果也同时表明Neotyphodium属的真菌与其有性型的Epichloё属真菌之间存在着较为复杂的遗传进化关系。一些Neotyphodium属的种是由Epichloё属的某一个种演化而来的,而更多的Neotyphodium属则源自两个或三个不同的Epichloё spp.的种间杂交[36-38]。例如,N.lolli,N.aotearoae和N.inebrians等是直接由Epichloё属的某一个种演化而来的[36,39],N.australiense是由E.festucae和E.typhina杂交后演化而来,N.occulans则是由E.bromicola和一个未确定的种杂交后演化而来。将无性型和有性型的内生真菌分别命名为“Neotyphodium”和“Epichloё”的分类法掩饰了真菌的进化多样性、生命周期及其与宿主的互作等众多问题,使得很多研究被复杂化或误解,如子座的形成在内生真菌有性繁殖过程中是必不可少的,部分Neotyphodium属内生真菌并没有完整的有性阶段,但仍在宿主表面形成了子座[32,40];很多Neotyphodium属内生真菌是由缺乏有性繁殖能力的内生真菌种间杂交而来,但存在部分有性型内生真菌由于其可产生含有子囊孢子的子囊壳而被归入Epichloё属[41]。由于Neotyphodium属与Epichloё属之间复杂的杂交进化关系,仅凭借形态学特征将Epichloё属的无性型归于另一单独属的分类方式存在巨大的缺陷,难以准确深层次对内生真菌进行分类与鉴定。因此,结合系统发育学证据对Epichloё属及其无性型Neotyphodium属内生真菌进行重新调整具有很大的必要性[14,36,42]。
2012年于澳大利亚墨尔本召开的第18届国际植物学大会上,规范了多性型真菌命名的规则,提出了“one fungus,one name”的原则。这一原则规定,每种真菌都应该有一个能表示其生长特性及孢子形态的单一名称。基于Neotyphodium属和Epichloё属之间错综复杂的杂交进化关系、双命名系统的缺陷以及对真菌所制定的新的命名规则,Leuchtmann等[43]于2014年对Epichloё属及其无性型Neotyphodium属真菌重新进行了规整。基于种的系统发育、交配兼容性概念与优先发表原则,将Epichloё属的无性型Neotyphodium属内生真菌全部归入Epichloё属。在Epichloё属中进行重新组合时,除E.mollis作为一个新名称外,其余均保留了原始分类单元名称。基于系统发育所确定的位置,将某些种归类至亚种或变种的水平,将同种异名的种进行了重新命名和规整,此外,去除了Acremoniumchilense(=Neotyphodiumchilense)以及Neotyphodiumstarrii两个种。重新命名规整之后,Epichloё属真菌共10个有性型和24个无性型,确认了3个亚种以及6个变种。
自对禾草内生真菌分类系统重新规整以来,新报道的种都按照新的分类系统进行分类。Tatsiana等[44]通过测定野生早熟禾(Poaalsodes)种群中内生真菌的侵染频率和变异性,根据内生真菌侵染野生早熟禾产生的生物碱来评估内生真菌的多样性,发现了北美地区的新种Epichloёalsodes和一个变种Epichloёschardliivar.pennsylvanica。Leuchtmann等[45]从马特威西早熟禾(Poamatthewsii)中分离鉴定了一个内生真菌新种,命名为Epichloёnovae-zelandiae,其具有三个拷贝,分别是E.amarillans,E.bromicola和E.typhinasubsp.poae。Tian等[46]对分离自我国西北地区中华羊茅的内生真菌的菌落和分生孢子形态特征进行了研究,并利用tefA和tubB基因内含子序列确定了该内生真菌的分类地位,结果表明该种为新种并将其命名为了Epichloёsinensis。Thünen等[47]从单花臭草(Meliauniflora)分离出了内生真菌,经鉴定发现该内生真菌是新种并将其命名为Epichloёscottii。同时,与其它有性型内生真菌tubB等[48]序列对比表明E.scottii也是一有性型种,另外发现该种是无性型E.dissecta的有性来源之一。Leuchtmann和Schardl利用tubB和tefA基因序列分析了欧洲三种短柄草属(Brachypodiumsylvaticum,B.pinnatum和B.phoenicoides)和三种拂子茅属(Calamagrostisarundinacea,C.purpurea和C.villosa)植物的Epichloё内生真菌多样性,发现从拂子茅属中分离出的两种内生真菌与其它内生真菌在系统发育树中居于不同分支,形态学和系统发育学证据表明这两个种内生真菌均为有性型内生真菌且为新种,因此分别命名为Epichloёcalamagrostidis和Epichloёftanensis。
2 禾草内生真菌的分类
截止2023年,已正式报道的Epichloё属内生真菌共有51种,其中包含21种(亚种、变种)有性型和30种(亚种、变种)无性型,各个种的宿主范围、来源、杂交特性、传播特性以及形态学特征见表1、表2和表3[43-47,49]。有性型内生真菌多为单倍体,即只存在单拷贝内含子基因序列,无性型内生真菌中,大部分为多倍体,即存在多拷贝内含子基因序列[50]。在所有有性型内生真菌中仅E.liyangensis为双拷贝杂交种,在所有无性型内生真菌中,E.australiensis,E.cabralii,E.canadensis,E.danica,E.disjuncta,E.funkii,E.guerinii,E.hordelymi,E.melicicola,E.occultans,E.pampeana,E.siegelii,E.sinica,E.sinofestucae,E.tembladerae,E.uncinata,E.hybrida,E.alsodes,E.schardliivar.pennsylvanica和E.sinensis这20种为双拷贝杂交种,E.chisosa,E.coenophiala和E.novae-zelandiae为三拷贝杂交种,而E.aotearoae,E.gansuensis,E.inebrians,E.mollis,E.sibirica和E.stromatonga这6种内生真菌均为单拷贝非杂交种,这类内生真菌可能是由有性型内生真菌丧失有性生殖能力进化而来。在杂交种中普遍发现了E.bromicola和E.typhina这两种拷贝,且这两种内生真菌的宿主种类也最为丰富,已在至少6种禾草中发现了E.bromicola,至少7种禾草中发现了E.typhina,这两种内生真菌是绝大部分杂交种的起源。

表1 有性型Epichloё属内生真菌分类Table 1 Teleomorph-typified species of Epichloё endophyte
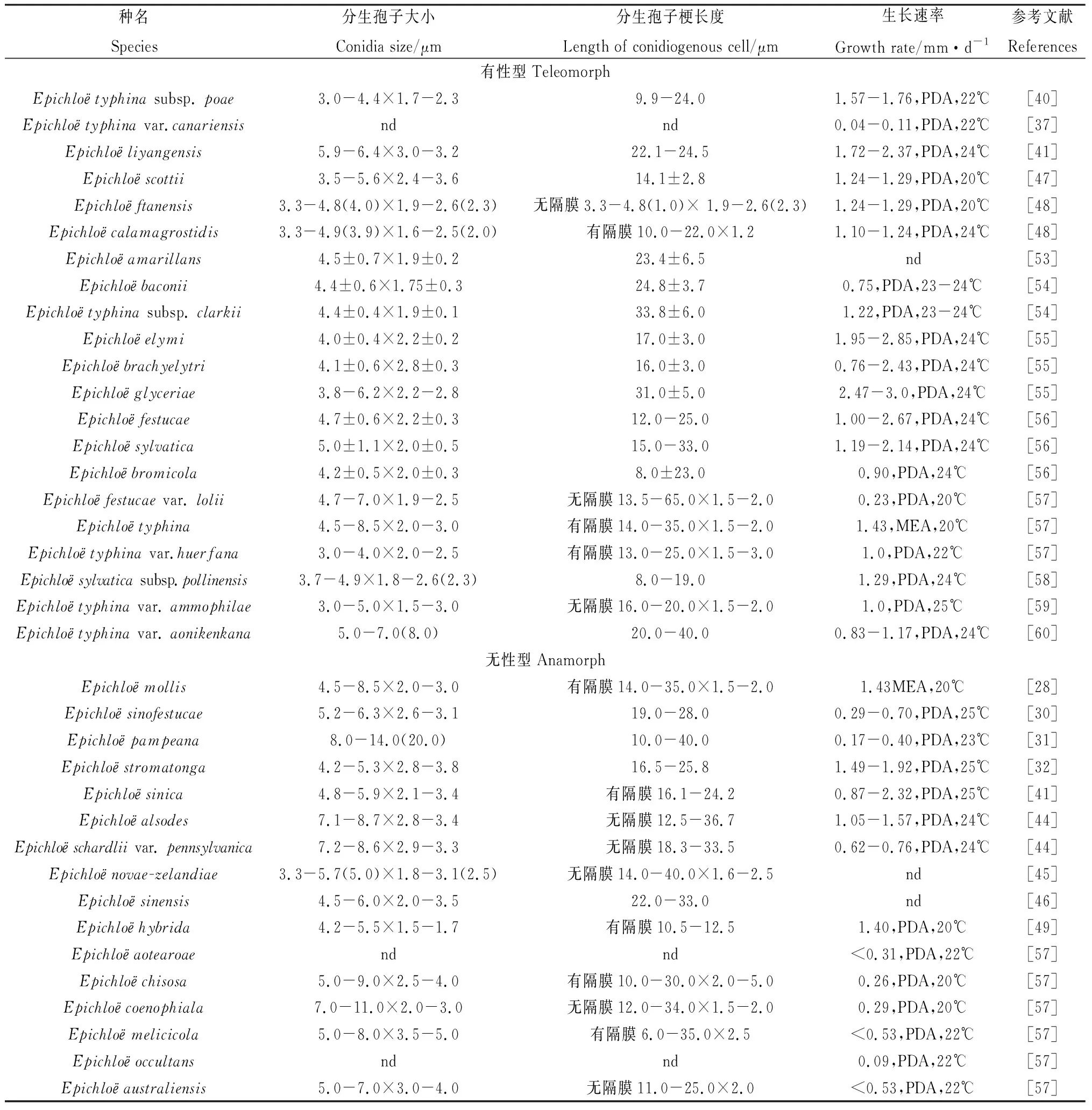
表3 Epichloё属内生真菌形态学特征Table 3 Morphological characters of Epichloё endophyte species
由表3可知,不同种禾草内生真菌的形态学特征均存在一定的差异。另有研究表明,同种禾草所携带的内生真菌以及同种内生真菌的形态学特征均会因地理区域的差异而不同[16]。如研究人员从我国内蒙古中东部地区6个不同地理种群的羊茅(Festucaovina)中共分离出19株形态特征各异的内生真菌[51];对我国西北地区不同地理种群、不同组织的21株E.gansuensis菌株进行形态多样性分析,结果发现这些菌株的菌落颜色、质地、生长速率、分生孢子大小、形状等都存在较大的差异[52]。
3 中国已发现的禾草内生真菌
我国Epichloё属内生真菌的分类学研究进展也取得了显著成果,迄今为止,已从我国至少21个属77种禾草中发现了Epichloё属内生真菌的存在,并通过进一步的分离与鉴定,报道了12种禾草内生真菌(表4)[66-67]。从醉马草中共分离出E.gansuensis和E.inebrians两种内生真菌[36,68-69];E.liyangensis主要定殖于草地早熟禾(Poapratensis)中;羽茅(Achnatherumsibiricum)主要被E.sibirica和E.gansuensis所侵染;从鹅观草(Elymuskamoj)中分离出了E.sinica;从小颖羊茅(Festucaparviglum)中分离出了E.sinofestucae;E.stromatolonga和E.typhina分别侵染拂子茅(Calamagrostisepigeio)和鸭茅(Dactylisglomerata)两种禾草[21,70-72]。已从披碱草(Elymusdahuricus)、圆柱披碱草(Elymuscylindricus)、肥披碱草(Elymusexcelsus)、垂穗披碱草(Elymusnutans)、西藏披碱草(Elymustibeticus)和麦宾草(Elymustangutorum)等几种披碱草属禾草中分离出了E.bromicola内生真菌[67],除披碱草属禾草外,从野大麦(Hordeumbrevisubulatum)[20]、羊草(Leymuschinensis)[73-74]、短柄草(Brachypodiumsylvaticum)[75]和大雀麦(Bromusmagnus)[76]中也分离出了E.bromicola内生真菌,除此之外,从我国青海和甘肃的垂穗披碱草(Elymusnutans)中还分离出了E.glyceriae。从采集自我国新疆等地的布顿大麦(Hordeumbogdanii)中分离出的内生真菌被鉴定为Epichloё sp. HboTG-2(H.bogdaniiTaxonomicGroup2:E.bromicola×E.typhina)[77]。

表4 我国Epichloё属禾草内生真菌分布Table 4 Distribution of Epichloё endophyte fungi in China
4 利用内生真菌进行种质创新研究进展
利用Epichloё属内生真菌进行种质创新,即选取优良菌株(可提高宿主抗逆性、仅产生抗虫生物碱而不产生对家畜有毒的生物碱),通过人工接种的方式将其接种至新宿主体内获得抗逆、对家畜无毒、可稳定遗传的新共生体、新种质[11,82],这种育种方式为育种学开拓了一个新的研究思路和技术[83]。20世纪30年代,美国育种科学家成功培育出第一个含有内生真菌的牧草新品种‘Kentucky-31’[84]。1991年,新西兰科学家利用人工接种技术将禾草内生真菌菌株Endosafe®接种至黑麦草中,构建了第一个人工禾草-内生真菌共生体[11]。自此,科学家们从大量内生真菌共生体中筛选出了不含对家畜具有毒性的生物碱而可产生抗虫生物碱的内生真菌菌株,将所筛选的有益菌株接种于牧草中构建新共生体,如将只产生loline和peramine两种抗虫生物碱的E.coenophiala内生真菌菌株的AR542接种于高羊茅品种‘Jesup’和‘Georgia 5’中,构建出了高越冬率和存活率的新共生体,且能防止家畜出现中毒症状[85];将AR542菌系接种到高羊茅品种‘Jesup’中创制出新品种MaxQTM[82];将UA4内生真菌菌系接种于高羊茅品种‘HiMag’中创制出了镁含量和钙含量较高的新品种ArkPlusTM,此品种保持了较低的家畜中毒率[86]。新西兰与澳大利亚联合也已发现了诸多有益内生真菌菌株如N.loliiAR1菌系以投放到市场中[87]。我国目前也已开展了利用禾草内生真菌进行抗性育种的研究,将分离于野大麦的内生真菌(Epichloёbromicola)通过人工接种的方式接种至栽培大麦(Hordeumvulgare)体内,创制出了野大麦内生真菌E.bromicola-大麦新种质,内生真菌将其提高原宿主植物生长的优良特征赋予新的宿主中[88]。除应用于牧草育种,禾草内生真菌目前也广泛应用于草坪草育种:美国、新西兰等多个国家已培育出诸多携带禾草内生真菌的抗逆草坪草新品种,其中坪用型高羊茅和黑麦草‘Advent’‘Assure’‘Dandy’‘Dasher II’‘Gettysburg’‘Pinnacle’等新品种已投放到市场中[89];新西兰科学家利用较高毒性的内生真菌菌株AR95和AR601联合培育出具有抗鸟采食的新品种‘AVANEX’,因其防鸟效果显著,该品种已在新西兰克赖斯特彻奇(Christchurch)国际机场和汉密顿(Hamilton)国际机场投入使用[90-91]。将禾草内生真菌应用于植物抗性育种,不仅能提高植物在不良环境中的生存能力,同时还能缓解水资源匮乏,减少化肥农药的使用,并且还能对盐碱地进行有效地开发利用。因此,充分利用现有资源并不断挖掘新禾草内生真菌来进行抗性育种具有极大的必要性和可行性。
5 总结与展望
自然生态系统中,几乎所有植物都能够与真菌形成共生关系[92],与导致宿主植物生长减缓,甚至死亡的致病微生物不同,禾草内生真菌在保证宿主植物应对外界环境变化过程中的生长发育方面发挥着重要作用。内生真菌的侵染可提高宿主对至少40余种害虫的抗性[93],也能提高禾草对细交链孢(Alternariaalternata)、离蠕孢(Bipolarissorokiniana)等多种病原菌的抗性[94]。除生物胁迫外,内生真菌对禾草抵御非生物胁迫的增益作用也已得到充分证明,如E.bromicola与野大麦共生可减轻盐胁迫对野大麦的伤害[20],E.sinensis可提高中华羊茅抗旱和抗盐碱能力,E.gansuensis与醉马草共生可有效提高醉马草抗寒、抗旱和抗盐碱能力[95-96],E.festucae和E.gansuensis等内生真菌还能提高宿主禾草对Zn,Al,Cd等重金属的耐受性[97-99]。除此之外,禾草内生真菌也会对宿主禾草根际土壤的物理、化学特性以及根际微生物生物量和微生物群落及功能产生影响[96,102-104]。内生真菌还能作为植物生长调节剂和生物农药的来源[100]。研究表明,内生真菌可通过产生生长素(Auxin,IAA)等植物激素来促进植物生长,已有学者通过离体培养高羊茅E.coenophiala内生真菌获得IAA。Latch研究发现,受内生真菌侵染的黑麦草植株生物量的增加与赤霉素(Gibberellins,GA)有关[101]。此外,部分内生真菌的代谢物具有抗病原菌和抗虫等生物学活性,因此可从这些内生真菌中提取出具有抗病虫活性的化合物,或可利用挖掘的抗病虫活性的化合物作为先导化合物进一步来研发新的生物农药[40,102]。目前,对内生真菌提高宿主抗逆性的研究大多只停留在生理层面,而对其分子机理研究仍不够深入,随着分子生物学技术的不断发展,今后应在禾草内生真菌共生体抗逆的分子机理研究方面不断深入,以此为禾草抗逆定向改良提供理论基础。
Epichloё属禾草内生真菌因其为禾草带来的诸多益处而具有较高的利用价值,同时其种类繁多,对其进行合理分类具有极大的必要性。通过形态学特征、DNA序列比对的系统发育关系以及生物碱合成基因鉴定和交配型分析等多种方法,可明确内生真菌的分类地位[68,103]。国内外学者对Epichloё属内生真菌的分类学地位进行了重新调整,理清了各个种之间所存在的直接或间接的联系,为内生真菌不同种之间所存在的种间杂交提供了充分的证据。同时,系统发育学方法的运用,极大地促进了我们对内生真菌进化起源的认知,为内生真菌在育种、植物生长调节剂和生物农药等方面的应用奠定了坚实的基础。
目前国内外有关禾草内生真菌的研究已取得了突破性进展,越来越多的优良资源被不断发掘。我国自20世纪90年代末开始就已陆续开展了禾草内生真菌的研究,虽起步较晚,但目前在禾草内生真菌多样性研究中成果显著,不断挖掘出了新种,已和其他国家趋于同步[15-16],但总体而言,我国禾草内生真菌资源调查区域仍不够全面,应该进一步加强我国禾草内生真菌种质资源调查,如应从冷季型草坪草到转向暖季型植物,对草原以外的区域也应进行资源调查,以期发现更多可应用于畜牧业生产的禾草内生真菌。小麦(TriticumaestivumL.)与水稻(Oryzasativa)等粮食作物也属于禾本科,已有学者尝试通过人工接种将禾草内生真菌接种至粮食作物中,但由于宿主特异性和不融合性而导致成功率较低,因此未来有必要探明禾草与内生真菌不融合的原因,利用基因组学和代谢组学等技术理清内生真菌与禾草之间信号转导与物质交流过程,从而利用基因编辑等技术来创制兼容性较高的禾本科植物-内生真菌共生体新材料、新种质。
猜你喜欢
现代苏州(2022年5期)2022-03-21
草地学报(2021年7期)2021-08-03
湖北农机化(2020年4期)2020-07-24
世界农药(2019年4期)2019-12-30
今日农业(2019年11期)2019-08-15
乡村地理(2018年2期)2018-09-19
南方农业·下旬(2017年11期)2018-01-09
中国饲料(2017年15期)2017-08-22
新疆农业科学(2016年6期)2016-07-23
杂草学报(2015年2期)2016-01-04
