
Bmttv在家蚕翅发育过程中的功能分析
2024-01-03 12:15:12李静雯陈胤霖贾文怡夏庆友化晓婷
昆虫学报 2023年11期
梁 燕, 李静雯, 陈胤霖, 贾文怡, 夏庆友, 化晓婷
(西南大学前沿交叉学科研究院生物学研究中心, 重庆 400715)
翅不仅仅是昆虫飞行的重要武器,更是昆虫适应环境、繁殖生存的一大利器。昆虫可以通过翅警示和躲避天敌。在进化生物学、生态学和生理学等多个领域,昆虫翅的研究都有着重要意义(Zera and Denno, 1997; McCullochetal., 2010)。家蚕Bombyxmori作为一种重要的经济昆虫,同时也是一种模式昆虫,完善的基因组信息和成熟的基因编辑平台为家蚕基因功能研究提供了良好的基础和平台(Xiaetal., 2004)。研究人员针对家蚕雏翅突变体进行了深入研究,发现其翅的短小是BombyxmoriApproximated(Bmapp)的启动子突变所导致(Wangetal., 2013);2022年,有研究团队鉴定并报道了Bombyxmorimacropterous(Bmmp)在家蚕翅发育过程中的功能(Zouetal., 2022)。
近年来,CRISPR/Cas9基因编辑技术(Maetal., 2014)的迅猛发展,使得通过敲除相关基因确定其表型再进一步探究其功能得以实现。基于此技术,在2020年,研究人员鉴定并获得了BmBlos2油蚕突变体,为解析油蚕的产生机制提供了模型(Yuetal., 2020)。1998年,ttv首次在果蝇Drosophila中被鉴定并发现其缺失会导致果蝇翅的缺陷(Bellaicheetal., 1998)。ttv作为糖基转移酶编码基因,属于EXT基因家族,是人类EXT1的同源基因,参与合成体内硫酸乙酰肝素(heparan sulfate, HS),进而合成硫酸乙酰肝素蛋白聚糖(heparan sulfate proteoglycans, HSPGs) (Takeietal., 2004)。HSPG是一种蛋白聚糖,由一个核心蛋白和与核心蛋白相连的HS粘多糖链组成(Linetal., 2004),可作为信号传播的共同受体促进信号传播,从而发挥调节细胞增殖、迁移和粘附的功能或者可以阻断配体与其受体结合,防止其接触同源受体以减少信号传播(Linetal., 2004; Xie and Li, 2019)。通过NCBI序列比对,在家蚕中也找到与之对应的3个基因ttv(GenBank登录号: BMSK0002707),botv(GenBank登录号: BMSK0008707)和sotv(GenBank登录号: BMSK0013095)。那么,在家蚕中,ttv是否具有与果蝇类似的功能参与家蚕翅发育目前仍是未知的。
本研究鉴定了家蚕Bmttv的CDS完整序列,进一步分析了蛋白结构域、同源序列比对、系统进化分析以及时空组织表达谱和亚细胞定位,研究结果不仅可为Bmttv的深入研究奠定基础,也为在家蚕中解析其翅发育功能提供参考。
1 材料与方法
1.1 实验材料
供试家蚕品种为D9L,所采用的家蚕胚胎细胞系BmE,用到的1180-Basic质粒均来自并保存于西南大学前沿交叉学科研究院生物学研究中心。使用新鲜的适龄叶饲养家蚕,保证蚕室的干净卫生,通风流畅,温度27 ℃。BmE 细胞所需培养基为含10%胎牛血清的Grace培养基,培养在27 ℃的恒温培养箱中,每48 h更换一次培养基,每5 d进行一次传代培养。
1.2 主要试剂
DNA提取试剂盒购于美基生物,RNA提取试剂盒、胶回收试剂盒均购于OMEGA,反转试剂盒、定量试剂均购于TaKaRa,质粒提取试剂盒购于Qiagen,Trans-T1和JM110感受态、限制性核酸内切酶购于NEB,引物由华大基因有限公司合成,抗体购于碧云天。
1.3 Bmttv CDS序列的克隆与鉴定
通过NCBI数据库(https:∥www.ncbi.nlm.nih.gov/)下载Bmttv序列,使用Primer 5设计BmttvCDS序列的引物Bmttv-CDS-F/Bmttv-CDS-R(表1),提取家蚕幼虫头部和中肠的基因组DNA,进行PCR扩增,PCR反应体系: DNA模板2 μL, PrimeSTAR Max Premix(2×) 25 μL, Bmttv-CDS-F/R引物各2 μL, RNase Free H2O 19 μL。反应程序: 98 ℃预变性30 s; 98 ℃变性10 s, 58 ℃退火20 s, 72 ℃延伸 30 s, 30个循环; 72 ℃延伸 5 min, 12 ℃保存。回收目的片段并进行T克隆,选取阳性克隆送华大基因进行测序。同时,使用RACE试剂盒(TaKaRa公司)进一步检测验证Bmttv基因序列,详细步骤参考试剂说明书。
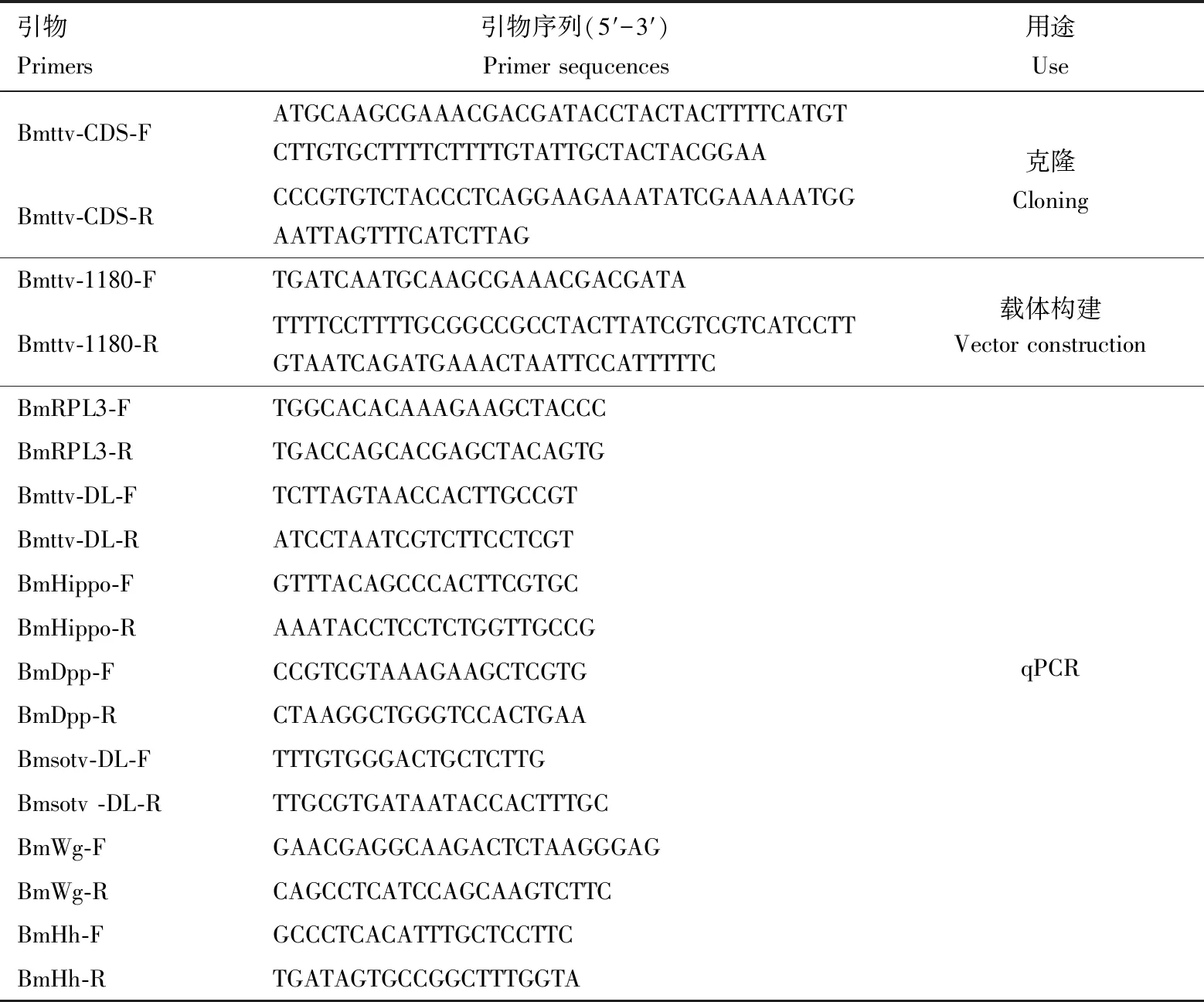
表1 引物信息
1.4 生物信息学分析
BmTTV蛋白序列来自silk DB3.0数据库,蛋白结构域预测网址是http:∥smart.embl-heidelberg.de/;黑腹果蝇D.melanogaster和智人Homosapiens的TTV同源基因CDS序列和蛋白序列均下载自NCBI数据库,同源序列比对网址是https:∥espript.ibcp.fr/ESPript/cgi-bin/ESPript.cgi;为了进一步了解BmTTV的系统进化关系,分别从NCBI数据库下载鳞翅目、双翅目、膜翅目以及哺乳动物TTV氨基酸序列进行系统进化关系分析。
1.5 基因表达量的qRT-PCR检测
为探究Bmttv在家蚕中的生物学功能,利用qRT-PCR分别检测Bmttv在家蚕不同发育阶段(5龄第1-7天幼虫、游走期幼虫、预蛹、1-9日龄蛹和1日龄成虫)、家蚕5龄第3天幼虫各组织(表皮、翅原基、精巢、气管、马氏管、丝腺、头、血淋巴、脂肪体和中肠)中以及在不同发育阶段翅原基中的表达量。上述样品按时期取材,液氮速冻后,冻存于-80 ℃冰箱,利用RNA提取试剂盒进行RNA的提取,使用反转录试剂盒以及实时荧光定量试剂盒(TaKaRa)的说明书进行qRT-PCR实验。每个实验样品均设置3个生物学重复,每个重复来自3头家蚕幼虫/3个细胞孔。以BmRPL3为内参基因。
1.6 细胞转染
待家蚕BmE细胞状态良好、密度达80%后进行铺板,选取12孔细胞培养板,提前铺入细胞爬片,取等量的细胞悬液加入细胞板中,轻微摇晃使细胞分布均匀,对照组使用的是1180-Basic质粒,实验组是Bmttv的过表达载体质粒(Bmttv-1180-Flag),每组设置3个重复孔。根据罗氏转染试剂说明书进行操作,待铺板12 h后进行转染,将质粒与转染试剂按1∶3(v/v)比例混合,再加入无抗无血清的Grace培养基;静置孵育15 min后,加入细胞中;转染6~8 h后更换为有抗生素(青霉素和链霉素)有血清的完全Grace培养基,转染48 h后进行免疫荧光实验。
1.7 免疫荧光亚细胞定位
将转染后的BmE细胞板里的培养基弃去,加入适量的4%多聚甲醛室温固定15 min,加入1×PBS置于摇床上漂洗3次,每次5 min;0.3% Triton X-100处理30 min,1×PBS漂洗3次,每次5 min;3% BSA室温封闭1 h,1×PBS漂洗3次,每次5 min;Flag抗体按1∶200(v/v)稀释,室温孵育2 h或4 ℃过夜孵育,1×PBS漂洗3次,每次10 min;Alexa Fluor 594标记的山羊抗兔IgG(Goat Anti-Rabbit IgG,Jackson ImmunoResearch)按1∶2 000(v/v)稀释,室温孵育1 h;使用DAPI染色(1∶10 000稀释),室温孵育30 min,最后将爬片取出倒扣于有适量抗荧光猝灭剂的载玻片上,使用指甲油封片,通过荧光显微镜进行观察拍照。为了进一步探究Bmttv的过表达是否会影响翅发育关键基因,通过NCBI数据库下载Bmttv,Bmwg,BmHippo,BmDpp,BmHh和Bmsotv序列,使用Primer 3(https:∥primer3.ut.ee/)在线网址设计其定量引物,通过实验检测后选择溶解曲线单一的引物进行qRT-PCR实验(表1),提取BmE细胞RNA,反转录合成cDNA,qRT-PCR检测上述基因的表达量,方法同1.5节。
1.8 数据分析
qRT-PCR的数据分析均采用2-ΔCt方法计算各个基因的相对表达量(Livak and Schmittgen, 2001),并使用双尾t检验进行统计分析,使用GraphPad Prism8.0软件绘图。
2 结果
2.1 Bmttv CDS的克隆与序列
silk DB3.0数据库中显示Bmttv位于家蚕5号染色体,包含8个外显子,7个内含子,编码741个氨基酸。通过PCR实验成功克隆到大小约为2 000 bp的序列,经测序验证,克隆的序列与silk DB3.0数据库中提供的序列完全一致,并通过RACE实验进一步确定了Bmttv的CDS全长序列,长2 228 bp。
蛋白结构域预测表明BmTTV包含两个结构域:一个EXT蛋白家族特有的结构域exostosin位于第19-362位氨基酸,符合与智人EXT1是同源基因这一特征;一个具有糖基转移酶活性的蛋白家族结构域-Glyco-transf-64位于第472-722位氨基酸,是硫酸肝素生物合成所必需的。同源序列比对分析结果显示(图1),家蚕、黑腹果蝇和智人TTV均包含以上两个结构域,且家蚕与黑腹果蝇和智人TTV的氨基酸序列一致性均达45%,具有较高的保守性,暗示着它们可能具有相似的功能。
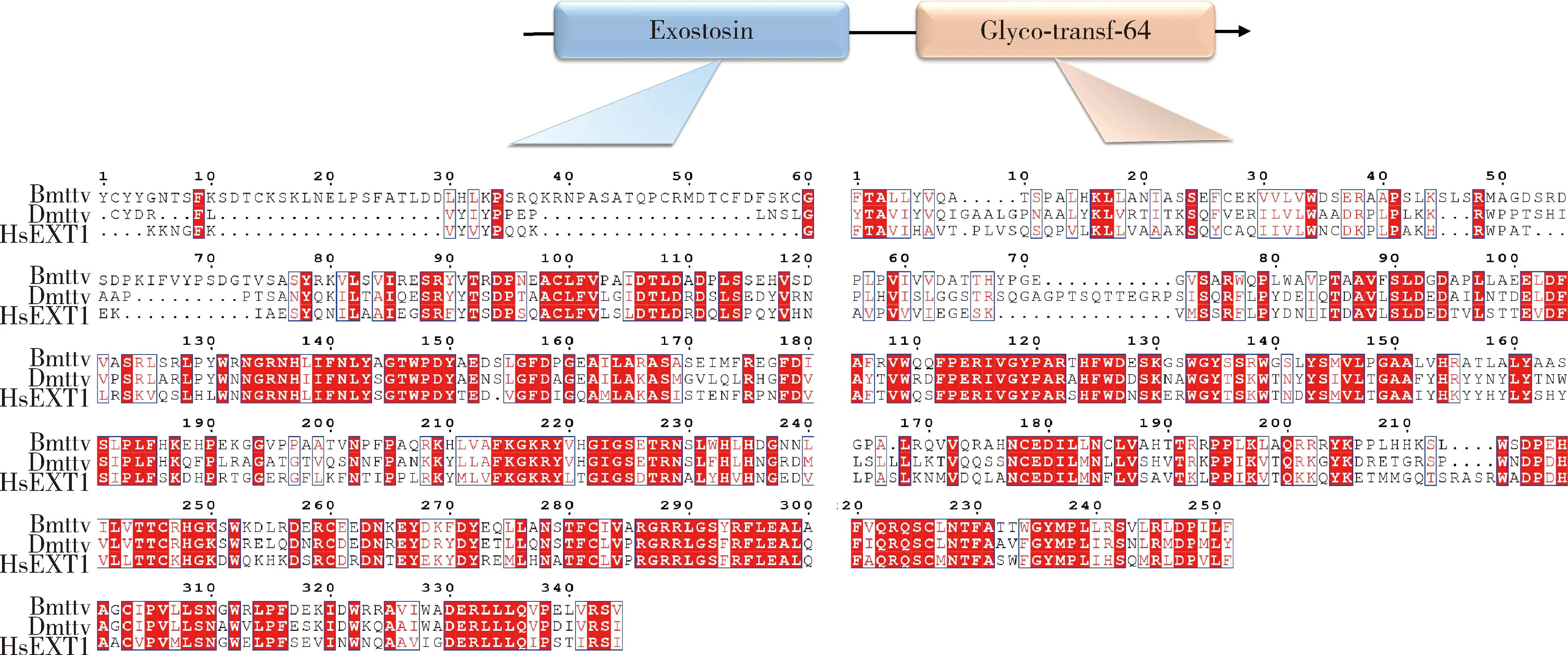
图1 TTV氨基酸序列比对
2.2 Bmttv的系统进化
系统发育树显示(图2)总体而言昆虫与哺乳动物的TTV各聚为一枝。在昆虫中家蚕BmTTV主要与亚洲玉米螟Ostriniafurnacalis、菜粉蝶Pierisrapae等鳞翅目昆虫TTV聚为一枝,以黑腹果蝇和冈比亚按蚊Anophelesgambiae为代表的双翅目以及蜜蜂代表的膜翅目昆虫TTV各自聚为一枝,暗示TTV在昆虫中广泛存在,保守性较高,且BmTTV与冈比亚按蚊和黑腹果蝇TTV的亲缘关系较近。
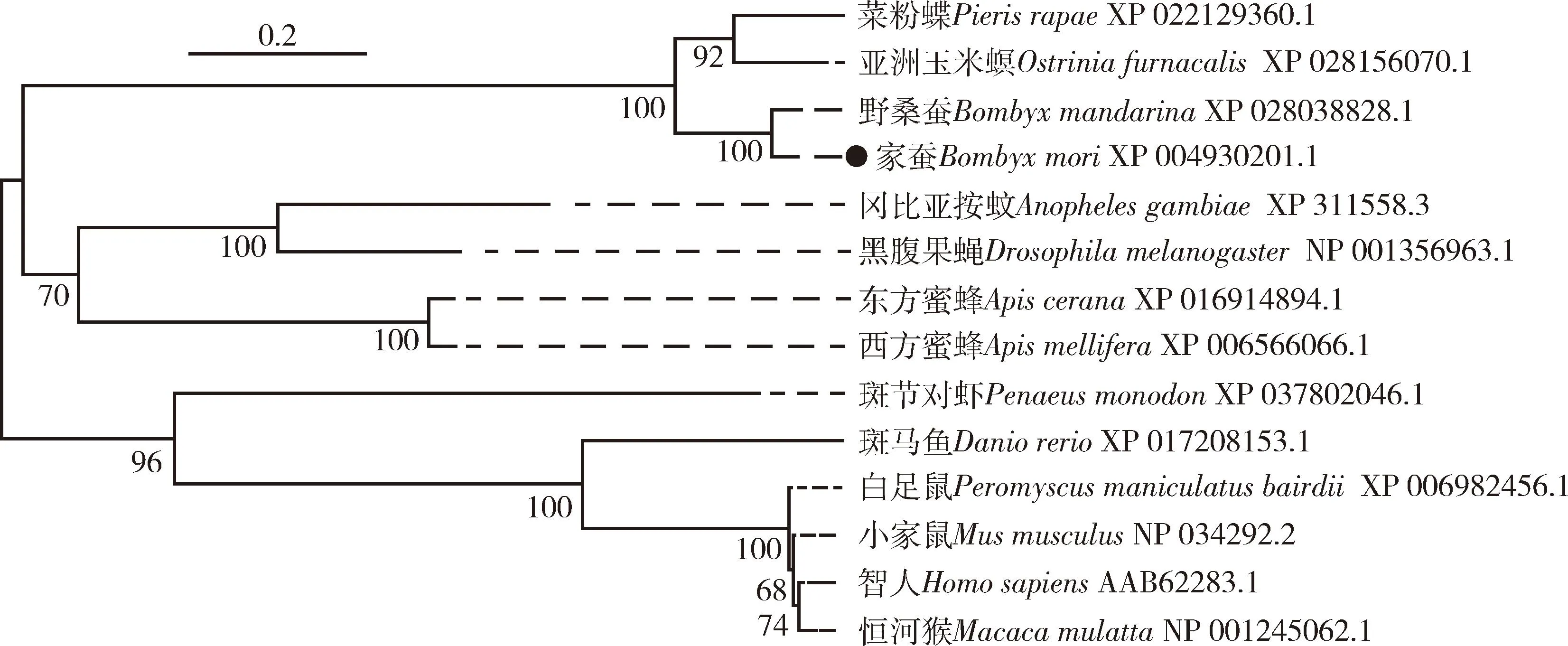
图2 基于氨基酸序列的家蚕BmTTV 和其他物种TTVs的系统进化树
2.3 Bmttv的时空表达特征
qRT-PCR检测结果显示,Bmttv在家蚕5龄幼虫期表达量较低,但从游走期直至成虫期表达量整体呈现上升趋势,且保持一个较高的表达水平,尤其在3日龄蛹出现一个峰值,随后在羽化前期再次出现一个高表达峰值,这暗示着Bmttv可能主要在家蚕蛹期发挥着重要的作用(图3: A)。为了进一步探究Bmttv在翅发育中的作用,进一步鉴定了其在家蚕翅原基发育过程中各个时期的表达情况(图3: B),发现Bmttv在翅原基中的表达趋势与在不同发育阶段表达谱较为一致,在5龄幼虫期表达量相对稳定,从游走期表达量开始上调,而游走期是家蚕翅发育的关键时期,因此,推测Bmttv在游走期高量表达可能与家蚕翅的发育相关。Bmttv在5幼虫中肠和精巢中表达量最高,在翅原基、马氏管和头中表达量次之,在丝腺表达量最低(图3: C),暗示Bmttv可能在家蚕大部分组织中都发挥比较重要的生物学功能。

图3 Bmttv在家蚕不同发育阶段整虫(A)和翅原基中(B)以及5龄幼虫不同组织中(C)的相对表达量
2.4 BmTTV的亚细胞定位
通过瞬时转染实验在细胞水平过表达Bmttv,免疫荧光结果显示BmTTV主要定位在BmE细胞质中(图4)。

图4 BmE细胞中BmTTV的亚细胞定位
2.5 BmE细胞中Bmttv过表达对翅发育关键基因表达量的影响
qRT-PCR检测结果显示,与过表达1180-Basic质粒对照组相比,过表达Bmttv后BmE细胞中Bmttv的表达量显著上调(P<0.001)(图5: A);Bmsotv(P<0.01)(图5: B)和BmHippo(P<0.05)(图5: C)的表达量显著下调,而Bmwg(图5: D),BmDpp(图5: E)和BmHh(图5: F)表达量无显著变化(P>0.05),暗示在家蚕中Bmttv和Bmsotv也存在着一定的相互联系,且Bmttv可能通过Hippo信号通路进而影响家蚕翅的发育。
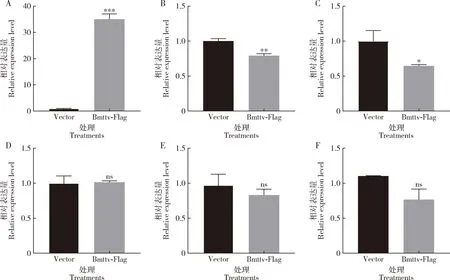
图5 BmE细胞中Bmttv过表达后家蚕翅发育关键基因的表达量
3 讨论
果蝇ttv基因作为HSPG生物合成中关键的糖基转移酶之一,在大多数物种中普遍存在。HSPG可作为多种细胞信号因子的共受体参与Hh, Wnt和Dpp信号通路的调控(Theetal., 1999; Busse-Wicheretal., 2014)。根据已有文献的研究表明,果蝇中ttv的敲除会导致其成虫期翅缘出现轻微损伤,部分翅脉出现杂乱无序的现象(Hanetal., 2004)。而在斑马鱼中ttv的缺失也会导致鱼鳍受损(Leeetal., 2004)。然而,Bmttv在家蚕中是否具有相似的功能仍然未知。本研究时空组织表达谱显示Bmttv在家蚕各个组织中普遍存在,在翅原基中表达量较高,且5龄幼虫期稳定低量表达,但发育至游走期表达量开始上升(图3),这与家蚕翅原基在游走期发育加快一致,暗示Bmttv与家蚕翅原基的发育相关。此外Bmttv在中肠、头部和精巢也高量表达(图3),可能还存在一些隐藏着的生理功能,有待进一步的探索。
本研究通过对BmTTV蛋白序列进行分析,其蛋白结构域包含EXT蛋白家族和一个糖基转移酶结构域,BmTTV与果蝇TTV氨基酸序列一致性达45%,说明该蛋白结构域序列具有一定的保守性。这暗示在家蚕中BmTTV极有可能存在着与果蝇TTV相似的功能,即作为关键的糖基转移酶参与合成HS,进而影响HSPG的合成,介导细胞信号因子的传递,最终影响个体的生理功能,比如翅发育,目前该部分实验仍在探索中。进化分析也表明ttv在鳞翅目昆虫中广泛存在,由此可推测若将ttv基因敲除,可能会影响这些害虫的飞行能力,从而减少农产品的虫害。
免疫荧光定位确定了BmTTV主要在细胞质中表达(图4),这也跟果蝇中ttv编码糖基转移酶,在内质网中产生,存在于细胞质内的报道相一致。此外,我们在细胞水平过表达Bmttv,通过qRT-PCR检测转录水平发现,Bmttv的表达量显著上调,而其同源基因Bmsotv表达量下调(图5),表明二者存在一定的联系,还有待进一步的探究。
本研究对部分已经有报道影响翅发育的关键基因BmHippo,BmDpp,Bmwg和BmHh进行检测,发现在Bmttv过表达的BmE细胞中,只有BmHippo是显著下调的,其余基因均无明显差异(图5)。而在家蚕中,已有研究表明Hippo的表达异常会导致家蚕翅的严重皱缩(Yinetal., 2020)。因此,我们推测Bmttv可能通过影响Hippo信号通路参与家蚕翅的发育。以上研究结果是解析Bmttv在家蚕变态发育过程中的功能的初步探究,为深入研究Bmttv在昆虫变态发育过程中的功能解析奠定了基础,不仅有助于了解昆虫翅的发育过程,也将为害虫的防治提供一定的理论基础与指导意义。
猜你喜欢
学苑创造·A版(2023年10期)2023-11-04 13:14:04
大自然探索(2023年11期)2023-03-01 09:04:36
四川蚕业(2022年2期)2022-11-19 02:09:52
学苑创造·A版(2022年3期)2022-03-29 23:32:16
四川蚕业(2021年2期)2021-03-09 03:15:30
四川蚕业(2021年1期)2021-02-12 02:03:20
学苑创造·A版(2019年6期)2019-07-11 01:07:39
计算机应用(2018年10期)2018-11-22 09:37:54
华南师范大学学报(自然科学版)(2017年4期)2017-09-11 09:16:23
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19 07:41:15
