
忍冬愈伤组织形成过程转录组分析*
2024-01-02 11:13陈宏宇孙庆文
贵州科学 2023年6期
陈宏宇,杨 烨,于 莹,黄 园,孙庆文,冉 飞
(1贵州中医药大学药学院,贵州 贵阳 550025;2贵州中医药大学基础医学院,贵州 贵阳 550025)
忍冬LonicerajaponicaThunb.是常用的药用植物,其干燥花蕾或带初开的花即金银花,具有清热解毒、疏散风热等功效。临床上用于痈肿疔疮、喉痹、丹毒、热毒血痢、风热感冒、温病发热等症。现代药理学研究表明,金银花具有抗炎、抗病毒、降血糖、抗氧化、保肝利胆、增强免疫、抗肿瘤细胞、神经保护等功能[1-2]。
组织培养技术在药用植物资源保护、快速繁殖、遗传改良和药效物质提取等方面有广泛、深入的研究,可用于解决诸如品质退化、种子病菌污染及农药残留等问题[3-4]。高通量测序技术用于大规模分析基因表达并识别基因调控通路[5-6]。利用转录组测序中的数字表达谱技术,对玉米幼胚胚性愈伤组织诱导过程中3个典型阶段样本进行了分析,识别了大量愈伤组织形成相关基因[7]。通过比较玉米幼胚胚性愈伤组织诱导过程中4个时期样本转录组,并识别microRNA,预测了靶基因与miRNA关系[8]。在小麦胚性愈伤组织形成的3个重要时期取样,构建转录组测序和miRNA测序文库,筛选了差异表达基因和差异表达miRNA[9]。
植物愈伤组织形成与很多类型的基因有关。利用分子生物学方法,发现ARF10基因的互作蛋白HBI1能促进愈伤组织起始及形成过程,并揭示其与WOX12、TIR1和AXR2等基因的调控关系[10]。利用基因工程验证了基因改变植物再生和转化效率[11]。这些研究为揭示植物组织培养技术中愈伤组织形成过程机制提供了重要基础。
忍冬分子生物学研究取得了较大进展[12],但是忍冬愈伤组织形成过程的分子机制研究相对较少。本实验对忍冬愈伤组织形成的3个时期分别进行转录组测序并比较分析,识别愈伤组织形成相关重要基因,分析调控通路,为进一步研究忍冬愈伤组织形成分子机制提供基础和参考。
1 材料与方法
1.1 材料
忍冬6年树苗购买于山东平邑,经贵州中医药大学杨烨鉴定为忍冬(LonicerajaponicaThunb.),取新生茎作为外植体。外植体用75%乙醇消毒30 s,10%次氯酸钠消毒15 min,无菌水冲洗3次,切成1 cm小段放入培养基(MS+0.5 mg/L 6BA+0.1 mg/L IAA),在恒温光照培养箱(25 ℃,14 h光照/天)中培养。从出现愈伤组织开始,至第5天(5d)、第10天(10d)和第15天(15d),分别取愈伤组织(图1),称取相同的质量,混样后经液氮冷冻提取总RNA。

图1 忍冬愈伤组织
1.2 RNA提取与测序
利用Trizol提取RNA,去除DNA,检查RNA质量和纯度后反转录,利用Illumina平台进行测序。原始测序数据利用软件SOAPnuke[13]去除接头序列(即接头污染)、含N碱基大于5%以及低质量读序(reads)(即质量值低于15的碱基占该reads总碱基数比例大于20%)。利用Trinity程序(v2.0.6)[14]从头拼接组装,聚类、拼接和去冗余处理后,得到非冗余单基因(unigene)序列。利用Nr(Non-Redundant Protein Database)、Nt、SwissProt、KEGG(Kyoto Encyclopedia of Genes and Genomes)、COG(Cluster of Orthologous Groups of proteins)、Pfam和GO(Gene Ontology)等数据库进行基因注释。
1.3 基因表达与调控通路分析
利用RSEM程序(v1.2.8)[15]的FPKM(Fragment Per Kilo Bases per Million Reads)方法计算基因表达量。利用DESeq[16]计算差异基因,以2个样本间基因表达差异log2FC大于2倍,且FDR≤0.001定义为显著差异表达基因(DEG)。利用GO数据库(Gene Ontology)对差异表达基因进行聚类分析,Qvalue ≤0.05定义为显著富集。利用KEGG数据库(Kyoto Encyclopedia of Genes and Genomes)对基因进行调控通路注释,推测基因的生物功能。
2 结果
2.1 测序与注释
对忍冬3个样本测序,去除低质量数据后,分别获得6.29Gb、6.40Gb和6.34Gb的clean data数据,所有样本Q20均大于97.98%,Q30均大于93.05%。从头组装后在3个组织共获得83 120条长的、非冗余单基因(unigene),其中长度为200~300 bp的单基因数量最多,有14 639个(图2)。在单基因中识别编码区,共得到47 263个CDS序列,长度分布在297~13 122 bp之间。利用Nr、Nt、SwissProt、KEGG、COG、Pfam和GO数据库分别注释了56 145、46 550、42 628、43 695、44 129、43 410和44 057条单基因,共58 668条单基因,其中7个数据库均能注释的基因有23 638个。

图2 组装基因长度分布
2.2 差异表达基因分析
由于发育程度不同,愈伤组织形成过程中3个阶段样本的基因表达预计有一定差异。经过计算,与5d样本相比,10d样本共有差异表达基因7 026个,其中上调表达3 829个基因,下调表达3 197个基因。类似的,与5d样本相比,15d样本共有差异表达基因12 218个,其中上调表达4 726个基因,下调表达7 492个基因。结果说明,随着忍冬愈伤组织的形成,基因表达模式发生较大变化,而且随着发育时间的延长,下调基因的数量明显较多。SERK、LEC2、WUS、AGL15和HBI1等基因与植物愈伤组织形成有关,基因表达量变化见表1。
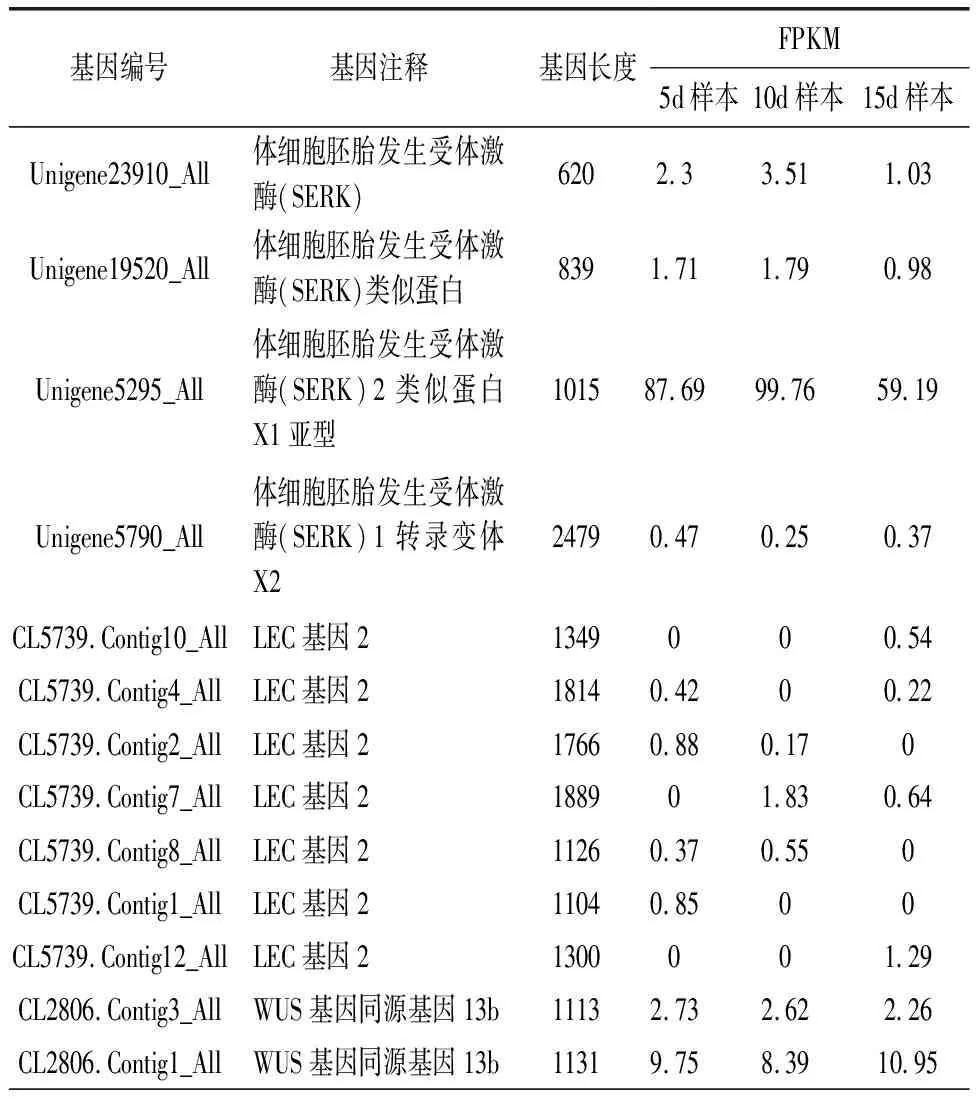
表1 忍冬愈伤组织形成相关基因的表达量
2.3 基因调控通路分析
利用GO聚类分析对显著差异表达基因进行功能分类,研究具有相同代谢过程或细胞途径的基因类群。结果表明,5d样本和10d样本之间差异表达基因有39个显著富集功能聚类,分为3大类:生物过程(biological process)、细胞组分(cellular component)和分子功能(molecular function),分别有22、2和15个功能聚类。如生物过程中的生物调控有包括GTPase活性激活、生长素分解代谢过程、生长素内稳态、碳水化合物稳态和细胞氧化还原稳态等差异表达基因614个。5d样本和15d样本之间差异表达基因有42个显著富集功能聚类,3大类分别为23、2和17个。如分子功能中的催化活性有包括酯键水解酶活性、鸟氨酸氨甲酰转移酶活性、β-葡萄糖苷酶活性、谷胱甘肽水解酶活性和核苷二磷酸酶活性等差异表达基因4427个。表2列举了每个类别显著富集程度最高的5个组。
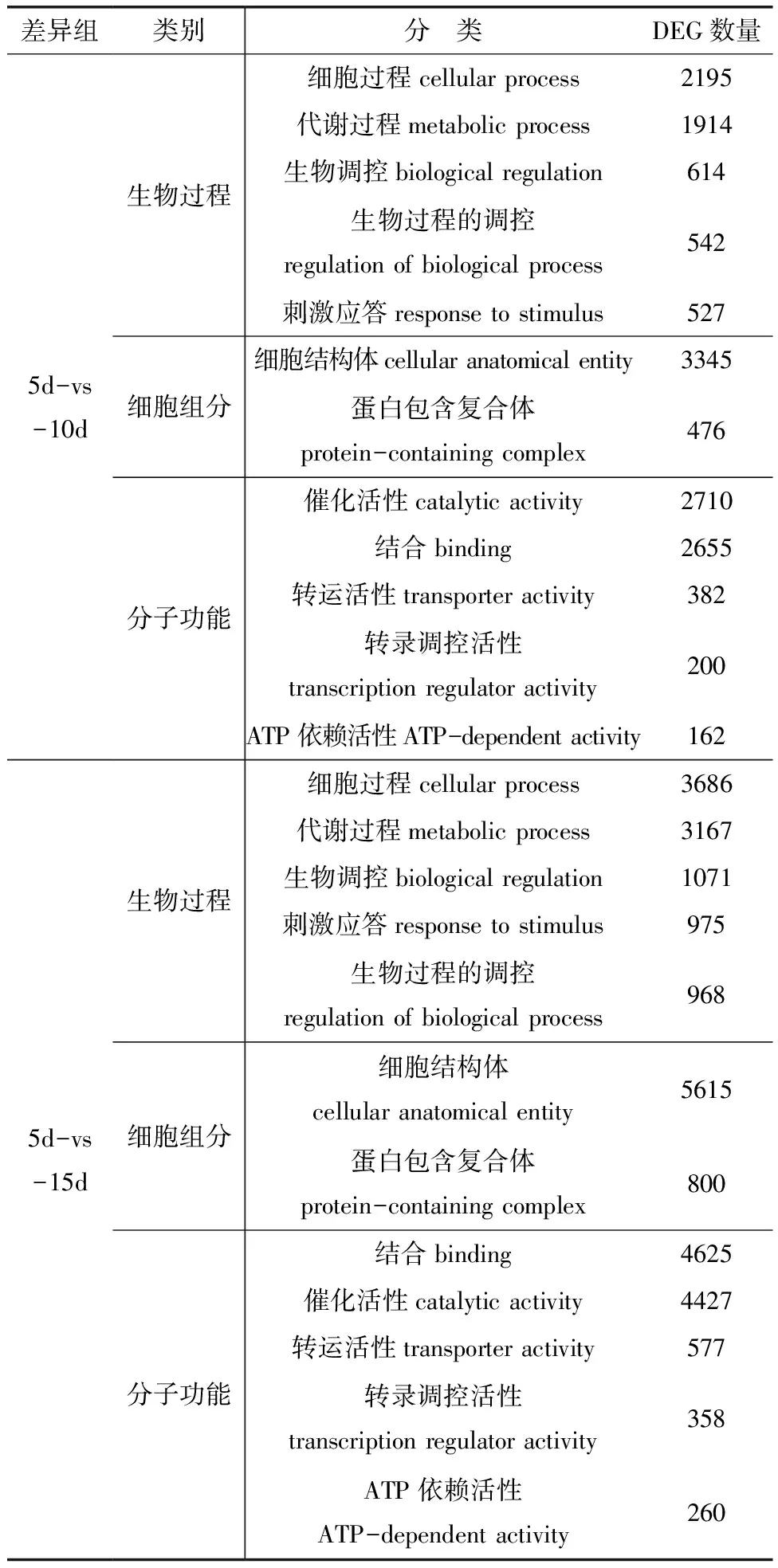
表2 忍冬愈伤组织形成过程GO聚类分析
利用KEGG数据库对差异表达基因进行生物学功能分析,推测参与生化代谢或信号转导途径基因的相互协调特征。结果表明,5d样本和10d样本之间差异表达基因归属通路分为5大类:细胞过程、环境信息过程、遗传信息过程、代谢和生物系统分别有1、2、4、11和1个通路。如“转运和分解代谢”“信号转导”“折叠、分类和降解”“定位”“碳水化合物代谢”和“环境适应”等通路富集程度较高。5d样本和15d样本之间差异表达基因与前者差异程度类似,基因总数更多。表3为显著富集的KEGG通路。

表3 忍冬愈伤组织形成过程KEGG通路分析
3 讨论
体细胞胚性愈伤组织的形成与发育是一个复杂的过程,包括诱导细胞分裂、胚性潜力诱导和重组体细胞等,其过程可能涉及很多类型的基因表达或抑制。随着分子生物学快速发展,很多基因的结构和功能逐渐被揭示出来。体细胞胚胎发生受体激酶(Somatic embryogenesis receptor kinase,SERK),富含亮氨酸重复序列,是体细胞胚发生相关类受体蛋白激酶基因。SERK基因最早在胡萝卜中发现,在胚性愈伤组织感受态阶段到胚性愈伤组织球形胚阶段均有较高表达,而在非胚性愈伤组织中表达量较低。拟南芥过表达SERK基因明显提高了胚性愈伤组织的诱导能力。赵玲玲等人克隆了芒属植物南获的SERK基因,利用荧光定量PCR证明基因在胚性愈伤组织中的表达量更高,并且随着愈伤组织的发育过程,其表达量先升高后下降[17]。本实验利用转录组分析了忍冬愈伤组织形成过程的基因表达信息,3个样本中共识别了4个可能的SERK基因,其中Unigene5295_All在3个样本中表达量均较高,但在样本之间差异较小(表1)。LEC基因(Leafy Cotyledon),包括LEC1和LEC2,最早发现于拟南芥。研究表明,当LEC基因发生突变时,拟南芥胚胎无法形成。本实验识别了7个可能的LEC基因,但表达量均相对较低(表1)。WUS基因(WUSCHEL)也与植物愈伤组织形成有关,研究表明WUS功能获得性突变能促进拟南芥体细胞转变为胚性细胞。本实验识别了7个可能的WUS基因,Unigene1500_All和CL5416.Contig1_All表达量相对较高,CL5416.Contig2_All在愈伤组织形成第3个时期表达量下降(表1)。AGL15基因(AGAMOUS-LIKE15),最初是在拟南芥用于鉴定胚发育的表达基因,在胚发育中表达且蛋白积累达很高,发芽后表达降低。本实验识别了5个可能的AGL15基因,但表达量均相对较低(表1)。HBI1基因(HOMOLOG OF BEE2 INTERACTING WITH IBH1)与愈伤组织起始有关。王龙等发现HBI1可以调控WOX12等基因,能促进愈伤组织起始及形成过程[10]。本实验识别了1个可能的HBI1基因,但表达量均相对较低,且在样本之间差异较小(表1)。
基于高通量测序技术的转录组分析在植物愈伤组织形成机制研究中有一定基础。罗旭等对玉米幼胚胚性愈伤组织诱导过程中3个典型阶段样本进行了分析,与幼胚对照相比,分别识别了4 825、5 119和5 463个差异表达基因,根据KEGG分析,玉米愈伤组织诱导相关基因可能分别集中在脂肪酸生物合成、细胞色素代谢外源性物质等途径[7]。江舟等比较了玉米幼胚胚性愈伤组织诱导过程中4个时期miRNA与靶基因,并按照调控关系建立了关联。这个过程中,有78个miRNA可能参与调控,并预测213个靶基因中的14个基因可能参与愈伤组织形成调控[8]。楚宗丽等比较了小麦胚性愈伤组织形成的3个重要时期转录组,在3、6和15时间阶段,识别了3 181、2 085和1 468个差异表达基因,同时也分别识别了30、33和18个差异表达miRNA[9]。本实验利用忍冬愈伤组织形成的3个时期,分别进行转录组测序,计算获取差异表达基因的数量和类别,推测差异基因的生物调控通路,为进一步研究忍冬愈伤组织形成分子机制提供基础和参考。
猜你喜欢
新民周刊(2022年27期)2022-08-01
西部林业科学(2021年4期)2021-08-11
传染病信息(2021年6期)2021-02-12
中南林业科技大学学报(2020年10期)2020-11-03
天津农学院学报(2020年3期)2020-10-14
学苑创造·A版(2020年12期)2020-01-07
中国外汇(2019年15期)2019-10-14
作文教学研究(2016年1期)2016-07-05
医学研究杂志(2015年8期)2015-06-22
生物医学工程学进展(2015年1期)2015-02-28
