
1 株芬芳镰刀菌的筛选鉴定及其提硒降镉效应
2024-01-01 00:00:00陈娥杜兴媛张朝阳马世龙黄子艺陈永波杨宝红
华中农业大学学报 2024年2期
关键词:真菌





摘要 为解决农作物硒镉共积累难题,采用选择性培养基从高硒镉背景值的植物根际土壤(土壤硒总含量50.187 mg/kg,土壤镉总含量50.311 mg/kg)中分离、筛选获得了1 株功能真菌,经鉴定为芬芳镰刀菌(Fusariumredolens),利用土壤模拟试验和盆栽试验测定土壤中有效硒、有效镉的变化和植物可食用部位Se 含量、Cd 含量,根据硒/镉富集和转运系数来评价真菌活硒抑镉的效果。土壤模拟试验结果显示,接种真菌后,活硒率与培养时间呈正相关,抑镉率与之相反;活硒率在第5 天达到最高,为60.9%;抑镉率在第3 天最高,为41.2%,第5 天最低,为27.3%。盆栽试验结果显示,添加108 cfu/mL 真菌,土壤pH 值比对照显著降低了0.35 个单位;对碎米荠幼苗生长无不良影响,平均株高为7.4 cm,平均根长为9.6 cm,平均生物量为2.9 g,均高于对照;土壤中有效硒的含量显著提高,活硒率最高达到24.8%,而有效镉的含量显著降低,抑镉率达到24.7%;碎米荠地上/地下部分总硒含量分别增加25.6% 和36.8%,总镉地上/地下部分含量分别降低32.7% 和11.6%。结果表明,真菌对天然高硒镉背景值土壤具有活硒抑镉能力,且添加浓度为108 cfu/mL 时效果最好,有助于碎米荠的富硒降镉。
关键词 硒; 硒镉伴生; 碎米荠; 真菌; 硒镉有效性
中图分类号 Q939.9 文献标识码 A 文章编号 1000-2421(2024)02-0099-10
中国富硒土壤中硒元素主要来源于黑色岩系,硒通常与重金属镉等伴生;而在一些非黑色岩系分布地区,如碳酸岩地区,土壤中的硒含量也达到富硒标准,也同时伴有重金属镉等元素[1-2]。相关性分析显示,镉-硒相互作用较强,存在显著的伴生现象[3],致使一些富硒地区的重金属含量超标,在南方一些富硒稻田中,土壤硒与汞、镉、铅和砷已呈现极显著或显著正相关[4]。还有研究显示,在较高镉含量(12mg/kg)情况下,土壤施硒反而促进植物根系对镉的吸收和积累[5]。在安徽池州石台县、海南富硒土壤区和陕南富硒区,均发现富硒土壤镉伴生现象,潜在生态危害系数达到98.89,属于强生态危害[6-8]。宋明义等[9]发现在浙西富硒地区中土壤及农产品富硒而镉含量超标是由母岩黑色岩系的硒、镉含量较高所导致。王锐等[10]对恩施市沙地乡的主要农作物(玉米、辣椒、土豆等)进行分析,发现各类农作物中硒含量均较高,但部分作物存在一定程度的镉超标情况。土壤中的镉可对土壤质量、农作物土壤生态功能产生一定程度的危害和影响,且土壤是农作物富集吸收硒、重金属等微量元素的主要来源,土壤镉污染可造成农产品食用健康风险,从而制约了富硒土壤资源的开发利用[11]。
我国的天然富硒区域主要集中在湖北恩施、陕西安康、广西等地。湖北省具有非常丰富的硒资源,特别是恩施地区,拥有全世界唯一探明的独立沉积型硒矿床,并被誉为“世界硒之都”[12]。堇叶碎米荠(Cardamine violifolia)属于十字花科多年生草本植物,在湖南省西部及湖北西部等地都有分布,无毒且营养价值高,是一种药食同源植物,由于其富集作用,尤其是堇叶碎米荠的超级富硒能力已成为近年来研究硒资源的热点与焦点之一[13]。为了降低碎米荠植株中的镉含量,湖北省恩施土家族苗族自治州农业科学院(以下简称恩施州农科院)成功研发野转家栽培技术,采用无土栽培,为企业提供了部分原料,但无土栽培生产成本居高不下,规模十分有限,且抛弃了恩施州天然富硒土壤资源的优势,浪费了宝贵的土壤硒资源。如恩施市新塘乡双河鱼塘坝硒矿区土地(土壤硒总含量50.187 mg/kg,土壤镉总含量50.311 mg/kg)在自然条件下生长的碎米荠叶片总硒含量为10.392 mg/kg,而叶片总镉含量高达84.900 mg/kg,碎米荠在自然高硒高镉伴生土壤上生长,富硒的同时也存在着重金属镉污染的现象,严重制约了硒资源的开发利用。
植物的根际是一个特殊的生态环境,其中聚居着大量的微生物,土壤中硒与镉的生物有效性、形态及迁移均受到土壤微生物的影响。目前,有关硒-镉相互关系的研究,主要关注外源硒肥料对水稻中镉累积的拮抗作用机制,以及外源硒药剂干预条件下人体内镉、硒的相互关系[14-15]。关于天然硒镉高背景区土壤中功能微生物的筛选鉴定和评价应用研究报道还不多见。因此,本研究从恩施州硒矿区附近的蒲儿草根际土壤中筛选到1 株能够耐受一定浓度硒镉的真菌菌株,通过室内评价真菌的活硒抑镉能力并结合盆栽试验,最终挖掘具有活硒抑镉功能的微生物资源,以期为开发利用硒资源、后续活硒抑镉机制研究和菌剂的开发提供基础资料。
1 材料与方法
1.1 供试材料
1)供试植物。堇叶碎米荠(Cardamine violifo⁃lia)穴盘幼苗由恩施州农科院提供,经测定,幼苗地上部分总硒含量为1.470 mg/kg,总镉含量为1.944mg/kg;幼苗地下部分总硒含量为2.404 mg/kg,总镉含量为0.996 mg/kg。
2)供试土样。筛选真菌的土样Pec,于2020 年5月12 日采集于恩施市新塘乡双河鱼塘坝硒矿区,土壤总硒、土壤有效硒分别为2.218、0.092 mg/kg,土壤总镉、土壤有效镉分别为1.620、0.665 mg/kg,Pec 对应植物蒲儿草叶片总硒、总镉分别为3.107、0.925mg/kg。用于土壤模拟试验、盆栽试验的土样MF 于2020 年7 月9 日采集于恩施市沐抚办事处,土壤总硒、土壤有效硒分别为10.28、0.26 mg/kg,土壤总镉、土壤有效镉分别为6.02、1.98 mg/kg。
3)试验主要试剂。引物、真菌基因组DNA 提取试剂盒和无毒核酸染料均购于上海生物工程股份有限公司,其他化学试剂均为国产分析纯或生物试剂级,购于湖北华圩科学器材有限公司。10% 吡虫啉可湿性粉剂购于江苏丰山集团股份有限公司,双面粘虫黄板购于山东曲阜市圣邦机械设备有限公司。
1.2 真菌筛选
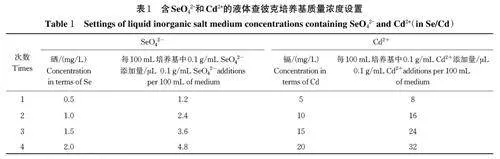
称取Pec 土样2 g ,接种于新鲜的100 mL 液体查彼克培养基中,28 ℃、150 r/min 振荡培养3~5 d。含SeO42-和Cd2+的液体查彼克培养基浓度设置见表1。吸取1.0 mL(带菌丝或3~5 个菌球)培养液按照表1 质量浓度设置依次以同样的培养条件转接培养,连续共转接富集培养4 次。采用稀释平板分离法在马丁孟加拉红培养基上涂布分离培养(真菌可以取10-1、10-2、10-3、10-4),培养3~5 d,肉眼初步观察菌落形态再显微观察菌丝形态,标记出所有不同的菌种并编号、描述和拍照。挑取单菌落,继续分离纯化培养,多次分离直到菌落大小、形态和颜色一致且无杂菌污染时,用马丁孟加拉红斜面培养基保存二级真菌菌株。
1.3 真菌分子生物学鉴定
利用真菌基因组DNA 提取试剂盒说明书,从培养3 d 的菌丝体中提取菌株总 DNA,采用真菌 ITS通用引物序列ITS1(5′-TCCGTAGGTGAACCT‐GCGG-3′) 和ITS4 (5′-TCCTCCGCTTATTGATATGC-3′),按照常规设置 PCR 反应体系进行扩增,将PCR 产物送广东美格基因科技有限公司测序,测序结果整理后提交NCBI,选取同源性较高和形态相近的菌株序列,用 MEGA7.0 中的邻接法构建系统发育树。
1.4 室内土壤模拟试验
1)灭菌。将10 g MF 土样分装于250 mL 三角锥形瓶中,在126 ℃下灭菌1 h,灭菌3 次,备用。
2)接种。将本文“1.2”分离保存的二级真菌菌株进行2 次活化后,在无菌操作条件下,用直径为5 mm的打孔器在活化培养的菌落周围生长旺盛的部分打取菌饼,接种到灭菌马丁孟加拉红培养液中,500 mL三角锥形装液量为150 mL/瓶,菌饼接种量为 3 个/瓶,将其置于恒温振荡仪中,于25 ℃、120 r/min 振荡培养3 d。
3)查彼克液体培养基制备。取50 mL 查彼克液体培养基添加至步骤1)锥形瓶中,备用。
4)将步骤2)中的种子液摇匀后,按照2% 的接种量,取10 mL 菌液(含有少量菌丝或2~3 个菌球)接种至步骤3)锥形瓶中,于28 ℃、150 r/min 下培养3~5 d,设置3 次重复,以不接菌液为对照。分别在第3天和第5 天摇匀后各取样30 mL,离心取上清液5 mL进行消化,用原子荧光光度计测定水溶态硒含量,剩余上清液通过中速定量滤纸过滤后,用ICP-MS 测定水溶态镉含量。因元素水溶态属于元素有效态的一部分,因此,试验在液体培养基中进行,测定上清液水溶态硒/镉量换算出土壤有效态硒/镉量。
1.5 盆栽试验
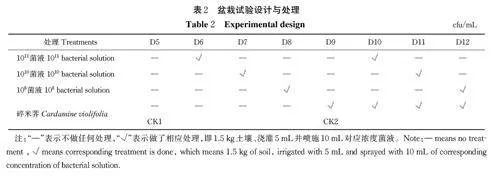
选用高12 cm、直径15 cm 的圆形塑料花盆,将已准备好的MF 土壤装盆,每盆装1.5 kg。试验设8 个处理(表2)。每个处理3 个重复,共计24 盆。使土壤全部浸湿,置于旱棚中平衡2 周,期间定期补水,将含水量保持在田间最大持水量的 60%。种植碎米荠前,将微生物均按照原始菌液(1011)、1010、108 cfu/mL 的浓度梯度,每个浓度按照5 mL/盆的添加量一次性浇灌到盆栽土壤中并喷施10 mL/盆(同浓度),不添加真菌的原始土壤作为对照1(CK1),只种植碎米荠的作为对照2(CK2)。每盆栽种5 株碎米荠穴盘幼苗,在碎米荠生长过程中定期补充水分,使土壤水分一直保持在田间持水量的60%,且每2 周更换盆栽位置,使每个盆栽光照均匀且充足。在碎米荠生长30 d 时第2 次施菌,施菌方式和浓度与种植前相同,种植 63 d 后植株成熟,收获碎米荠,并分成地上部分和地下部分,用自来水冲洗后再用去离子水冲洗干净,测量碎米荠根长、株高,碎米荠地上部分和地下部分在105 ℃下杀青30 min,然后在 70 ℃下烘至恒质量并测定碎米荠生物量,用于地上部分和地下部分总硒、总镉测定。另外,从每个盆中采集混合均匀的土壤样本约100 g,经自然风干、研磨并分别过孔径0.85、0.25和 0.15 mm 尼龙筛后,用于土壤pH、总硒、总镉、有效硒、有效镉测定。
土壤pH 测定:准确称量经过预处理的土壤样品(2.0±0.000 5) g 于50 mL 离心管中,加入20 mL 去离子水(土水质量比为1∶10),振荡30 min,取出静置30 min,测定上清液的pH。
碎米荠生物量测定:将碎米荠在70 ℃下烘干至恒质量,称取干质量。植株总硒测定参照GB5009.93—2017《食品安全国家标准 食品中硒的测定》进行。植株总镉测定参照标准GB5009.15—2014《食品安全国家标准食品中镉的测定进行。土壤总硒参照尹明等[16]的方法进行。土壤总镉测参照文献[17]进行。土壤有效硒测定参照NY/T 3420—2019《土壤有效硒的测定-氢化物发生原子荧光光谱法》进行。土壤有效镉测定参照HJ 804—2016《土壤8 种有效态元素的测定二乙烯三胺五乙酸浸提-电感耦合等离子体发射光谱法》进行。
1.6 数据处理
采用Microsoft Excel 2010 软件处理数据,用GraphPad Prism 8 软件制作图表,用SPSS17.0 软件进行差异显著性分析。
2 结果与分析
2.1 真菌筛选与分离
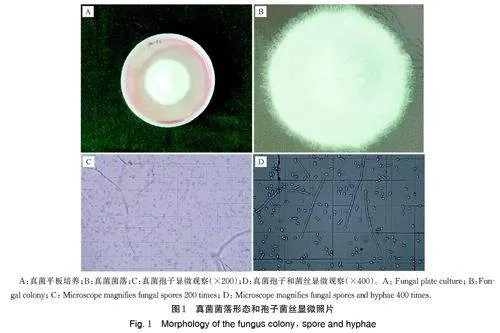
本研究从Pec 根际土壤中分离获得1 株真菌,并编号为Pec-z3,用于后续试验。在马丁孟加拉红固体培养基上,菌落致密呈白色绒毛状, 分生孢子镰刀型、多胞、无色(图1)。
2.2 真菌分子生物学鉴定
真菌菌株Pec-z3 经 rDNA-ITS 基因测序,在NCBI 中进行BLAST 比对分析,选取同源性较高的序列与 Pec-z3 构建系统发育树(图2),发现菌株Pecz3与芬芳镰刀菌(Fusarium redolens) X94169.1 聚在同一个分支,序列同源性为98%;另外,菌株Pec-z3形态特征与芬芳镰刀菌(F. redolens) 一致。因此,Pec-z3 菌株为芬芳镰刀菌(F. edolens) 。
2.3 真菌室内土壤模拟试验结果
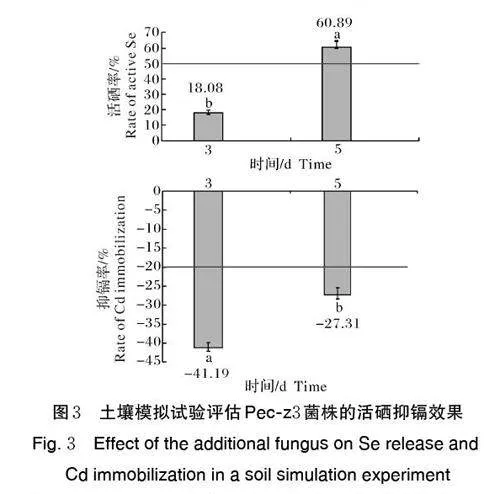
在含土培养基中接种真菌,结果(图3)显示,在培养第3 天时,真菌菌株Pec-z3 对土壤活硒率达到18.08%,抑镉率为41.19%;在培养第5 天时,活硒率达到60.89%,抑镉率为27.31%。第5 天活硒率显著高于第3 天,活硒率增加了42.81 百分点;与活硒率相反,第3 天抑镉率显著高于第5 天,抑镉率第3 天达到最大,但第3 天和第5 天抑镉率均超过20%。
2.4 盆栽试验结果
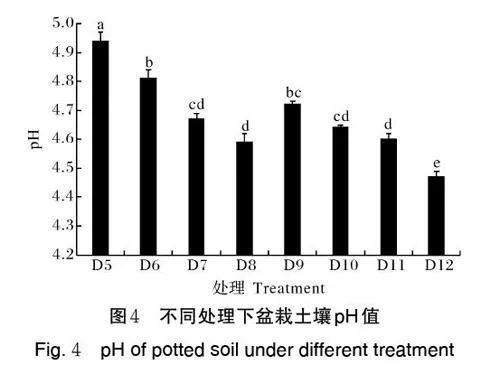
1)Pec-z3 菌株对盆栽土壤pH 值的影响。盆栽试验进行63 d 后,采集不同处理的土样测定土壤pH值,由图4 可见,处理组D6、D7、D8 与D5(CK1)pH 值差异显著,分别比对照低0.13、0.27、0.35 个单位,说明添加不同浓度的真菌菌液不同程度地降低了土壤pH 值。CK2(D9) pH 值显著低于D5(CK1),降低0.22 个单位,说明种植碎米荠也可以降低土壤pH值。处理组D10、D11、D12 与D9 存在显著性差异且均低于D9,其中D12 显著低于D5 和D9,pH 分别降低0.47 和0.25 个单位,说明种植碎米荠并添加108cfu/mL 的真菌菌液,土壤pH 降低程度最大。
2)Pec-z3菌株对碎米荠幼苗生长的影响。由图5可见,添加不同浓度的真菌对碎米荠的株高没有显著影响;添加108 cfu/mL 的真菌菌液处理(D12),碎米荠根长显著高于(D9),生物量与D9(CK2)无显著性差异。表明Pec-z3 菌株对碎米荠生长无不良影响。
从碎米荠盆栽表型结果(图6)可见,碎米荠在整个种植周期内正常生长,叶片颜色鲜绿,均无病害发生。虫害主要为蚜虫和潜叶蝇,种植期间分别采用叶面喷施70% 吡虫啉水分散粒剂(施药1 次,稀释2 000 倍)和黄板诱杀,喷施吡虫啉未影响碎米荠的生长。
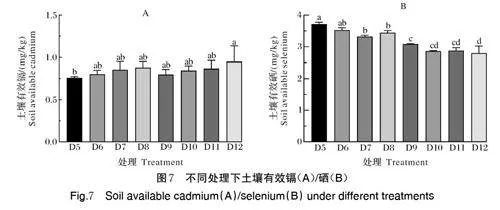
3)Pec-z3 菌株对土壤有效硒、有效镉的影响。由图7A 可以看出,处理D6、D7、D8 与D5(CK1)相比,土壤有效硒含量呈逐渐升高趋势,处理D10、D11、D12 与D9(CK2)相比,土壤有效硒含量同样为逐渐升高趋势。处理D9、D10、D11、D12 与处理D5(CK1)、D6、D7、D8 比较,各组之间差异不显著。处理D9 与D5 差异不显著但高于D5,与D5 相比,活硒率为4.82%。D12 中的土壤有效硒显著地高于处理D5,与D5 相比,活硒率为24.84%,D12 与其他处理差异不显著但高于其他处理,与D9 相比,活硒率为19.05%。由图7B 可以看出,处理D6、D7、D8 与对照D5 相比,土壤有效镉含量呈逐渐降低趋势,处理D10、D11、D12 与对照D9 相比,土壤有效镉含量同样呈逐渐降低趋势。处理D7、D8 显著地低于D5,与D5 相比,抑镉率分别为7.35%、10.65%。处理D9、D10、D11、D12 与处理D5、D6、D7、D8 比较,抑镉率分别为16.92%、18.85%、16.42%、15.78%。处理D12 显著地低于D9,与D9 相比,抑镉率为9.42%,与D5 相比,抑镉率为24.74%。
4)Pec-z3 菌株对碎米荠地上和地下部分总硒、总镉的影响。由图8A 可以看出,不同浓度的Pec-z3菌株处理后,D12 碎米荠地上部分的硒总含量为3.715 mg/kg,显著高于D9(CK2),硒总含量比D9 高出25.64%;与D9 相比,碎米荠地下部分硒总含量呈向上升趋势且均高于对照D9,其中D12 最高,比D9(CK2)高出36.82%。由图8B 可以看出,不同浓度的真菌处理后,碎米荠地上部分的镉总含量均低于对照D9,其中D12 显著地低于D9,镉总含量比D9 低32.72%;不同浓度的真菌处理后,与对照D9 相比,碎米荠地下部分镉总含量呈下降趋势,且均低于对照D9,其中D12 最低,比对照D9 低11.63%。
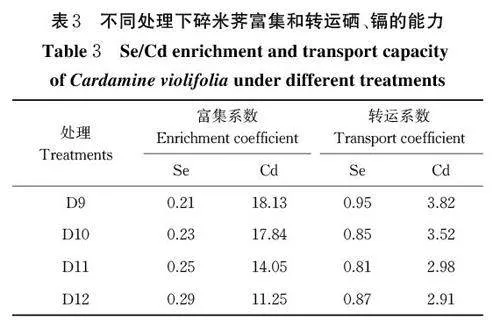
5) Pec-z3 菌株对碎米荠富集和转运硒、镉能力的影响。由表3 可知,加入真菌后,碎米荠对 Se 的富集系数均低于临界值(1.00),对 Cd 的富集系数均高于1,表明相较Se 而言,碎米荠对 Cd 具有更好的富集能力。不同浓度的真菌处理后,碎米荠对 Se 的富集系数各处理组均大于对照组D9,碎米荠对Cd 的富集系数各处理组均小于对照组,从侧面说明真菌对土壤中的硒具有活化作用、对镉具有抑制作用,添加108 cfu/mL 真菌对土壤的活硒抑镉能力影响最大。向土壤中施加不同浓度的真菌后,碎米荠对Se 的转运系数均小于1 且均低于对照组,说明Se 元素主要富集在碎米荠根部,添加不同浓度的真菌后抑制了Se 从地下部分到地上部分的转移能力;碎米荠对Cd的转运系数均大于1 且均低于对照组,说明Cd 元素主要富集在碎米荠叶部,添加不同浓度的真菌后抑制Cd 元素从地下部分到地上部分的转移能力,且影响碎米荠对Cd 的转运能力强于Se ,说明在天然高硒高镉的土壤上,碎米荠地上和地下部分的富镉能力要强于富硒能力。
3 讨论
本研究从恩施州硒矿区附近的蒲儿草根际土壤中筛选出1 株能够耐受一定硒镉浓度的真菌,经过菌落形态和分子生物学鉴定该真菌为芬芳镰刀菌(Fu⁃sarium redolens) 。Booth[18]及张素轩[19]认为芬芳镰刀菌(F. redolens ) 是镰刀菌属美丽组(section elegans)中 1 个具有重要经济意义的种。Zhang 等[20]从结香中分离到多株具有抗菌活性的内生真菌,如F.oxysporum var. redolens,属于芬芳镰刀菌属真菌。研究表明,芬芳镰刀菌的多种次生代谢产物都表现出较好的抗菌活性[21-22],徐利剑等[23]从盾叶薯蓣中分离获得了内生芬芳镰刀菌并研究了Dzf2 中的2 个抗菌活性成分,其中镰刀菌酸是芬芳镰刀菌的活性物质之一。Son 等[24]发现镰刀菌酸具有温和的毒性,在体外和体内均显示出对假单胞菌的抗卵菌活性,可以抑制植物病原卵菌和真菌的菌丝体生长,同时还可以有效抑制植物致病菌细胞的生长。刘奇[25]发现芳香镰孢属真菌HNU066 可以提高生物滴滤塔反应器活性填料对含硫化氢废气的清除效率,与对照组相比可提高34.5%。
本研究通过室内土壤模拟试验发现,芬芳镰刀菌具有活硒抑镉能力。添加108 cfu/mL 真菌并种植碎米荠,土壤中有效硒含量可提高24.84%,只添加108 cfu/mL 真菌,土壤中有效硒含量可提高15.64%。只种植碎米荠,土壤中有效硒可提高4.82%,活硒率均低于二者共同处理的活硒率24.84%;添加108 cfu/mL 的真菌并种植碎米荠,土壤中有效镉含量可降低24.74%,只添加108 cfu/mL 的真菌,土壤中有效镉含量可降低10.65%。只种植碎米荠,土壤中有效镉可降低16.92%,抑镉率均低于二者共同处理的抑镉率24.74%,说明土壤中有效硒/镉的含量既受真菌的影响又受碎米荠的影响,且添加108 cfu/mL的真菌后土壤中有效硒含量最高,有效镉含量最低,即活硒率和抑镉率均达到最高。综上,碎米荠和真菌都对土壤有效硒/镉有影响,同时碎米荠对土壤有效硒与有效镉影响低于真菌的影响,说明在硒镉伴生土壤中,真菌对土壤活硒抑镉的能力强于碎米荠。
真菌是否具有活硒抑镉能力以及对硒镉伴生土壤修复的效果最终体现在指示植物中硒/镉含量的变化情况,植物中的硒/镉含量是评估硒/镉生物利用度/毒性的直接指标之一。本研究中,不同浓度Pec-z3 处理后碎米荠地上部分总硒含量增加3.19%~25.64%,地下部分总硒含量增加14.58%~36.82%。碎米荠地上部分总镉含量降低9.82%~32.72%,地下部分总镉含量降低2.36%~11.63%。表明不同浓度的真菌有增加碎米荠地上和地下部分总硒含量、降低总镉含量的作用。添加108 cfu/mL 菌液对碎米荠地上和地下部分总硒、总镉含量影响最大,其中地上部分总硒含量增加25.64%,总镉含量降低32.72%,地下部分总硒含量增加36.82%,总镉含量降低11.63%。
富集系数与转运系数的大小和植物种类、土壤类型、土壤环境、重金属种类、重金属生物可利用性、强化剂种类等多种因素有关。富集系数和转运系数的大小可以间接反映土壤重金属污染植物修复的效果。研究发现,碎米荠是一种极具潜力的重金属污染的指示和修复植物[26],相较硒而言,碎米荠对镉具有更好的富集能力,硒元素主要富集在碎米荠根部,镉元素主要富集在碎米荠叶部。本研究中碎米荠地上部分对于镉的积累量最高为122.573 mg/kg(gt;100 mg/kg),碎米荠地下部分对于镉的积累量最高为32.117 mg/kg,说明碎米荠具有潜在的土壤镉修复价值。
目前,已有细菌或植物对土壤中Cd、Cr、Pb、Zn、Cu、Ni、Mn、As、Hg 等重金属进行单独或联合修复的研究[27-28],但暂时还没有真菌或植物单独或联合作用后能够提高植物可食用部位有益元素Se 降低有害元素Cd 含量相关的研究。本研究主要进行了小规模的盆栽试验,还处于初步模拟研究阶段,实际应用还有待进一步的田间试验验证。本研究没有涉及芬芳镰刀菌活硒抑镉的机制,只研究了芬芳镰刀菌单一使用于1 种指示植物的效果,今后将继续筛选出具有活硒抑镉能力的细菌、放线菌等,组合使用或联合其他植物来解决不同土壤和植物硒镉伴生的问题。
参考文献 References
[1] 张泽洲. 典型农作物中硒形态分析及其硒-镉相互作用研究
[D]. 武汉:中国地质大学,2019.ZHANG Z Z.Selenium speciation
and selenium-cadmium interaction in staple crops[D].
Wuhan:China University of Geosciences,2019 (in Chinese
with English abstract).
[2] 韦雪姬,王磊,卓小雄. 桂中典型碳酸盐岩区水稻及根系土硒
含量特征[J]. 地球与环境,2020,48(3):369-376.WEI X J,
WANG L,ZHUO X X.Characteristics of selenium contents in
rice and rhizosphere soil in a typical carbonate region of central
Guangxi[J].Earth and environment,2020,48(3):369-376 (in
Chinese with English abstract).
[3] 沈燕春. 贵池富硒区伴生重金属及其生物效应研究[D]. 合
肥:安徽农业大学,2011.SHEN Y C.Research of the selenium-
rich area in Guichi associated with heavy metals and its biological
effects[D].Hefei:Anhui Agricultural University,2011
(in Chinese with English abstract).
[4] 孙协平,周广文,罗友进,等. 硒镉伴生对富硒土壤柑橘生长
及生理代谢的影响[J]. 核农学报,2020,34(3):661-668.
SUN X P,ZHOU G W,LUO Y J,et al. Effect of selenium
cadmium associated on growth and physiological metabolism
of Citrus in Se-enriched soil[J].Journal of nuclear agricultural
sciences,2020,34(3):661-668 (in Chinese with English abstract).
[5] 李荣林,王胜兵,李优琴. 锌硒对油菜吸收和累积镉的影响
[J]. 江苏农业学报,2008,24(3):274-278.LI R L,WANG S
B,LI Y Q.Effects of zinc and selenium on absorption of cadmium
in rapeseed[J]. Jiangsu journal of agricultural sciences,
2008,24(3):274-278( in Chinese with English abstract).
[6] YANG B B,YANG C,SHAO Z Y,et al.Selenium( Se) does
not reduce cadmium (Cd) uptake and translocation in rice
(Oryza sativa L.) in naturally occurred Se-rich paddy fields
with a high geological background of Cd[J]. Bulletin of environmental
contamination and toxicology,2019,103(1):
127-132.
[7] 耿建梅,王文斌,温翠萍,等. 海南稻田土壤硒与重金属的含
量、分布及其安全性[J]. 生态学报,2012,32(11):3477-3486.
GENG J M,WANG W B,WEN C P,et al. Concentrations
and distributions of selenium and heavy metals in Hainan paddy
soil and assessment of ecological security[J].Acta ecologica
sinica,2012,32(11):3477-3486 (in Chinese with English
abstract).
[8] DU Y J,LUO K L,NI R X,et al.Selenium and hazardous elements
distribution in plant-soil-water system and human
health risk assessment of Lower Cambrian,Southern Shaanxi,
China[J]. Environmental geochemistry and health,2018,40
(5):2049-2069.
[9] 宋明义,岑静,胡艳华,等. 高镉地质环境富硒土壤特征及其
生态效应[J]. 地球与环境,2012,40(3):354-360.SONG M
Y,CEN J,HU Y H,et al.Study on the characteristics of selenium-
rich soil under geological environment with high contents
of cadmium and its ecological effects[J]. Earth and environment,
2012,40(3):354-360( in Chinese with English abstract).
[10] 王锐,侯宛苓,李雨潼,等. 高硒高镉区土地安全区划方法[J].
环境科学,2019,40(12):5524-5530.WANG R,HOU W L,
LI Y T,et al.Land safety zoning method in high-selenium and
high-cadmium areas[J].Environmental science,2019,40(12):
5524-5530( in Chinese with English abstract).
[11] 安凤秋. 外源重金属铅和镉对土壤生物活性及微生物群落多
样性的影响研究[D]. 杨凌:西北农林科技大学,2018.AN F
Q.Influence on soil biological activity and bacteria community
diversity by exogenous lead and cadmium[D]. Yangling:
Northwest A amp; F University,2018 (in Chinese with English
abstract).
[12] 杨礼茂. 鄂西南地区硒资源及其综合开发[J]. 地域研究与开
发,1998,17(4):72-76. YANG L M. Selenium resources in
southwest Hubei and its comprehensive development[J].Areal
research and development,1998,17(4):72-76(in Chinese).
[13] 林樾. 堇叶碎米荠硒形态分析及其富硒多肽对肝癌细胞作用
研究[D]. 无锡:江南大学,2021.LIN Y.Speciation analysis of
selenium in Cardamine violifolia and its selenium enriched peptides
activity in hepatoma cells[D].Wuxi:Jiangnan University,
2021( in Chinese with English abstract).
[14] RIAZ M,KAMRAN M,RIZWAN M,et al.Cadmium uptake
and translocation:selenium and silicon roles in Cd detoxification
for the production of low Cd crops:a critical review[J/
OL].Chemosphere,2021,273:129690[2023-02-06].https://
doi.org/10.1016/j.chemosphere.2021.129690.
[15] ZWOLAK I.The role of selenium in arsenic and cadmium toxicity:
an updated review of scientific literature[J]. Biological
trace element research,2020,193(1):44-63.
[16] 尹明,李家熙. 岩石矿物分析[M].4 版. 北京: 地质出版社,
2011: 845-846.YIN M,L J X. Rock mineral analysis[ M].4th
ed.Beijing:Geological Publishing House,2011: 845-846.
[17] 徐聪,赵婷,池海涛,等. 微波消解-ICP-MS 法测定土壤及耕
作物小麦中的8 种重金属元素[J]. 中国测试,2019,45(5):
85-92. XU C,ZHAO T,CHI H T,et al. Determination of
eight kinds of heavy metal elements in cultivated soil and the
wheat by microwave digestion-ICP-MS method[J]. China
measurement amp; test,2019,45(5):85-92 (in Chinese with
English abstract).
[18] BOOTH C.The genus Fusarium[M].Farnham Royal:Commonwealth
Agricultural Bureaux for the Commonwealth Mycological
Institute,1971.
[19] 张素轩. 镰刀菌属分类进展[J]. 真菌学报,1991,10(2): 85-
94.ZHANG S X. Advances in the taxonomy of the genus Fusarium
[J]. Acta mycological sinica,1991,10(2): 85-94( in
Chinese).
[20] ZHANG H W,RUAN C F,BAI X L.Isolation and antimicrobial
effects of endophytic fungi from Edgeworthia chrysantha
[J/OL]. Bangladesh journal of pharmacology,2015,10(3):
529[2023-02-06].https://doi.org/10.3329/bjp.v10i3.23575.
[21] XU L J,LIU Y S,ZHOU L G,et al. Enhanced beauvericin
production with in situ adsorption in mycelial liquid culture of
Fusarium redolens Dzf2[J]. Process biochemistry,2009,44
(10):1063-1067.
[22] 李静. 滇重楼内生真菌及其抗菌活性检测和甾体化合物分析
[D]. 雅安:四川农业大学,2007.LI J.Antimicrobial activity
and steroid analysis of the endophytic fungi from Paris poly⁃
phylla var. yunnanensis[D]. Yaan:Sichuan Agricultural University,
2007( in Chinese with English abstract).
[23] 徐利剑,李培琴,赵江林,等. 内生芬芳镰刀菌Dzf2 中两个抗
菌活性成分[J]. 天然产物研究与开发,2010,22: 564-567.
XU L J,L P Q,Z J L,et al.Two antimicrobial active components
in endophytic Fusarium fumigatus Dzf2[J].Natural products
research and development,2010,22: 564-567.
[24] SON S W,KIM H Y,CHOI G J,et al.Bikaverin and fusaric
acid from Fusarium oxysporum show antioomycete activity
against Phytophthora infestans[J].Journal of applied microbiology,
2008,104(3):692-698.
[25] 刘奇. 芳香镰孢属真菌HNU066 及其在含硫化氢废气降解中
的应用: 201910876615.3[P]. 2020-02-21. LIU Q. Fusarium
aromaticum fungus HNU066 and its application in the degradation
of hydrogen sulphide-containing exhaust gases:
201910876615.3[P].2020-02-21.
[26] 黄东华,麦淑华,仇曙,等. 镉对堇叶碎米荠生长生理特性的
影响[J]. 湖北农业科学.2022,61(5): 87-90.HUANG D H,
MAI S H,QIU S,et al.Effect of cadmium on the growth and
physiological characteristics of the corydalis capensis[J].Hubei
agricultural science.2022,61(5): 87-90.
[27] 刘新亮,刘梦茹,杨亚东,等. 重金属污染土壤修复技术研究
现状及发展方向[J]. 化工矿物与加工,2021,50(10): 52-57.
LIU X L,LIU M R,YANG Y D,et al.Research status and development
direction of heavy metal contaminated soil remediation
technology[J].Chemical minerals and processing,2021,50
(10):52-57.
[28] MAHAR A,WANG P,ALI A,et al.Challenges and opportunities
in the phytoremediation of heavy metals contaminated
soils:a review[J]. Ecotoxicology and environmental safety,
2016,126:111-121.
(责任编辑:张志钰)
猜你喜欢
学苑创造·A版(2022年11期)2022-05-30 14:29:21
食品安全导刊(2021年21期)2021-08-30 08:21:48
科学(2020年2期)2020-08-24 07:56:56
小学科学(学生版)(2018年8期)2018-08-21 12:05:50
现代园艺(2017年23期)2018-01-18 06:58:18
新农业(2016年21期)2016-08-16 11:52:07
天然产物研究与开发(2016年11期)2016-06-15 20:29:17
微生物与感染(2015年5期)2015-12-08 07:03:20
大豆科技(2015年1期)2015-03-23 08:27:26
营销界(2015年23期)2015-02-28 22:06:21
