
祁连山草地生态系统优势物种生长季的生态化学计量特征
2024-01-01 00:00:00张婷朱晓鹏徐海燕吴晓东刘桂民甘子鹏毛楠李莉莎薛守业康国慧殊秋丽陈卓
草地学报 2024年6期


摘要:植物生态化学计量是许多过程模型的重要参数,为了解高海拔地区优势物种的生态化学计量特征及其与土壤养分的关系,本文测定祁连山高寒草地生态系统5种优势物种叶片和根系的生态化学计量,分析了其与根际土壤养分的关系。结果表明:随着生长季推移,优势物种叶片总有机碳(Total organic carbon,TOC)含量相对稳定,全氮(Total nitrogen,TN)含量下降,而C∶N和C∶P则增加;大部分物候期下,青藏大戟(Euphorbia altotibetica Paulsen)的TN和全磷(Total phosphorus,TP)含量高于其他优势物种,且C∶N和C∶P低于其他优势物种,而垂穗披碱草(Elymus nutans Griseb.)的TN含量低于其他优势物种,且N∶P和C∶P高于其他优势物种;祁连山地区植物生态化学计量受到土壤化学计量的影响,植物生长主要受到N元素的限制。研究结果为相关植被模型和陆面过程模型的参数化提供了重要的基础数据。
关键词:草地生态系统;生态化学计量;优势物种;生长季;植物器官
中图分类号:S812.2""" 文献标识码:A"""" 文章编号:1007-0435(2024)06-1810-09
Ecological Stoichiometric Characteristics of Dominant Species in the
Grassland Ecosystem of Qilian Mountains During Growing Season
ZHANG Ting1,2, ZHU Xiao-peng1, XU Hai-yan1*, WU Xiao-dong2, LIU Gui-min1,
GAN Zi-peng1, MAO Nan3, LI Li-sha1, XUE Shou-ye1, KANG Guo-hui1,
SHU Qiu-li1, CHEN Zhuo1
(1. School of Environmental and Municipal Engineering, Lanzhou Jiaotong University, Lanzhou, Gansu Province 730070, China;
2. Cryosphere Research Station on the Qinghai-Tibet Plateau, State Key Laboratory of Cryospheric Science, Northwest Institute
of the Eco-Environment and Resources, Chinese Academy of Sciences, Lanzhou, Gansu Province 730000, China;
3. China Agricultural University, Beijing 100193, China)
Abstract:Plant ecological stoichiometry is an important parameter in many process models. To understand the ecological stoichiometric characteristics of the dominant species at high altitude region and their relationship with soil nutrients,we determined the ecological stoichiometry of leaves and roots of five dominant species in alpine grassland ecosystems in the Qilian Mountains,and analyzed their relationship with soil nutrients. The results showed that:with the growing season process,the total organic carbon (TOC) content in leaves of the dominant species was relatively stable,while the total nitrogen (TN) content was decreased,while the C∶N and C∶P was increased. In most phenological periods,the contents of TN and total phosphorus (TP) of Euphorbia altotibetica Paulsen were higher than those of other dominant species,and the contents of C:N and C:P were lower than those of other dominant species,while the TN content of Elymus nutans Griseb was lower than that of other dominant species,and N:P and C:P were higher than those of other dominant species. The ecological stoichiometry of plants in the Qilian Mountains is affected by soil stoichiometry,and plant growth is mainly limited by N. Our results provide important data the parameterization of many vegetation models and land-surface process models.
Key words:Grassland ecosystem;Ecological stoichiometry;Dominant species;Phenological periods;Plant organs
生态化学计量(Ecological stoichiometry)是表征植物养分平衡的一个重要指标,它对于理解物种相互作用、植物养分限制、利用效率和整体生活史策略具有重要意义,同时也是许多生物地球化学模型的重要参数,可用来预测植物对气候和环境条件变化的响应[1-3]。碳(Carbon,C)是生命骨架的基本成分[4],氮(Nitrogen,N)对蛋白质和有机物的合成至关重要,磷(Phosphorus,P)是各种酶、磷脂和核苷酸的基本成分[5],它们共同影响植物的光合作用、呼吸作用、初级生产力、群落组成、寿命和物种丰富度[6-8]。研究表明,植物的C∶N和C∶P是生物量与养分含量的比值,一般可以用来反映植物的养分利用效率[3],植物的N∶P比率则通常用于确定环境养分的可用性[9-10]。
目前植物生态化学计量已经在区域和全球尺度上对进行了广泛研究[8,11],主要集中在海拔[12]、坡度和坡向[13]、植物器官[14]、物候期[15]、放牧[16]以及气候变化,如变暖[17]、干旱[18]和CO2浓度增加[19]等对植物生态化学计量平衡的影响。可见,植物生态化学计量与植物本身和多个环境因子相关,但大多数研究侧重分析单一物种的植物生态化学计量,揭示其与生长、发育和新陈代谢的关系[20-22],这对于认识由多个物种组成的植被群落对环境因子变化的响应还远远不够。
青藏高原分布着世界上最大的天然高寒草地[23],其面积达1.28×108 hm2,约占青藏高原总面积的50%。整体而言,青藏高原海拔高、辐射强、气温低、土壤发育较差[24]。这些自然环境形成了独特的植被群落,且植被对这些环境因子表现出了较好的适应策略。其中,在生态化学计量方面,已有研究发现植物N含量和N∶P比值较高,P含量较低,可能导致高寒地区植物生长速率较低[25]。然而,目前高海拔地区植被群落生态化学计量特征及其与环境因子的关系还不清楚。
祁连山位于青藏高原北缘,该地区为典型的草地生态系统脆弱地区[26],植物对环境变化敏感[27]。该地区广泛分布着青藏高原典型的草甸、草原和荒漠草原等植被类型,物种也有很好的代表性。因此,研究该地区植物的生态化学计量有助于认识青藏高原草地植物对环境变化的响应和适应策略。本研究依照植物生长季节(返青期、草盛中期、草盛后期、枯黄期),对祁连山高寒草地生态系统5个优势物种的C,N,P化学计量特征进行了研究,并分析了与土壤生态化学计量的关系,研究旨在分析祁连山不同优势物种的C,N,P含量及其生态化学计量特征以及祁连山优势物种与土壤生态化学计量特征的关系。研究结果有助于了解祁连山优势物种在整个生活史周期中的生长状况、营养状况、限制性因素以及与土壤环境的关系,揭示祁连山地区优势物种对寒旱环境的适应机制,为青藏高原植物的研究及生态环境保护提供科学依据,从而促进对该地区草地生态系统多样性的保护。
1 材料与方法
1.1 研究区概况
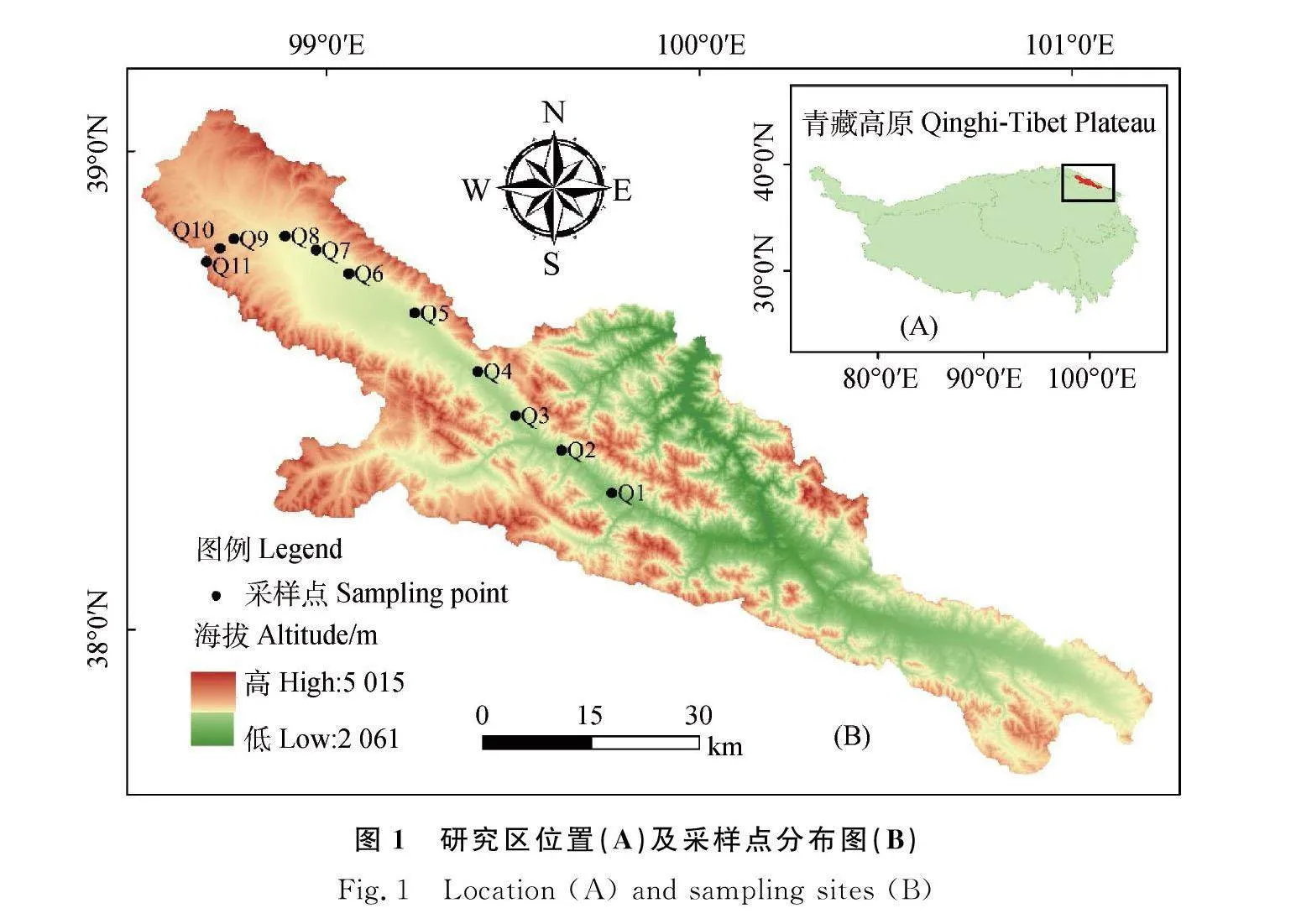
本研究位于青藏高原东北部的祁连山黑河上游野牛沟流域(98°44′E~99°46′E,38°18′N~38°50′N),具体研究区位置及采样点分布如图1所示。该地区海拔范围为2 800~4 500 m[28],年平均降水量为200~700 mm,年平均气温为-2.6~2.6℃[29]。该区域草地类型主要有高寒沼泽草甸、高寒草甸和高寒草原等[30]。主要优势物种为紫花针茅(Stipa purpurea Griseb.)、鸢尾(Iris tectorum Maxim.)、青藏大戟(Euphorbia altotibetica Paulsen.)、黑褐穗薹草(Carex atrofuscoides subsp. Minor(Boott) T. koyama)和垂穗披碱草(Elymus nutans Griseb.)等。
1.2 样品采集
依照植物生长季节(返青期、草盛中期、草盛后期、枯黄期)[31-32],本研究分别于6月、7月、8月、9月中旬在青藏高原东北部祁连山黑河上游野牛沟流域,对11个样点的5种优势植物的叶片和根系进行了采集,具体信息见表1所示。同时采集植物根际土壤,海拔梯度以100 m为间隔,共计11个不同海拔,分别编号为Q1~Q11。采样点坡度和坡向基本一致,并且受到人为因素干扰较小,采样的同时用GPS测定经纬度、海拔等样地信息,并记录草地类型及主要优势种等状况(表1)。
土壤样品采集:采用抖落法[33-34]与五点取样法(样方大小为10 m×10 m)相结合的方法,将优势物种整株挖起,抖动脱落土,混匀后装入自封袋并做好标记,置于4℃车载冰箱运送回实验室,用于土壤总有机碳(Total organic carbon,TOC)、全氮(Total nitrogen,TN)和全磷(Total phosphorus,TP)含量的测定(样品数量:11个样点×4个物候期 = 44个样品)。
植物样品采集:采用直接收获法与五点取样法相结合的方法,在同一样地的5个样点分别用剪刀与地面平齐剪下约10 g健康的植物叶片,用土钻挖取0~20 cm 地下部分植物根系约10 g,然后将每个样地5个样点内采集的叶片和根系分别混合均匀,用去离子水清洗干净后用吸水纸迅速擦干,装入信封袋中并做好标记,运回实验室用于植物TOC,TN和TP含量的测定(样品数量:4个物候期×11个样点×2个植物器官= 88个样品)。
1.3 测定方法
将采集好的植物样品于65℃烘箱烘干,研磨并过100目筛后,用于植物TOC,TN,TP含量的测定;土壤样品放置于室内自然风干,研磨并过100目筛后,用于土壤TOC,TN和TP含量的测定,具体方法参考鲍士旦的《土壤农化分析》[35],TOC含量采用高温燃烧法(Elementar VarioEL,Hanau,Germany)测定[36],TN含量采用半微量凯氏定氮法(Foss-8400全自动凯氏定氮仪)测定,TP含量采用全自动间断化学分析仪(Smart Chem 200,AMS-System,Rome,Italy)测定[37]。
1.4 数据处理
本研究中土壤、优势物种叶片和根系的C∶N∶P比均为元素质量比,数据以均值±标准偏差(SD)表示,对不同优势物种的生态化学计量特征进行LSD(Least-significant different)多重比较分析和单因素方差分析(One-Way ANOVA)。优势物种叶片、根系与土壤生态化学计量的相关性分析采用皮尔森(Pearson)双尾检验法。
2 结果与分析
2.1 优势物种根际土壤的生态化学计量特征
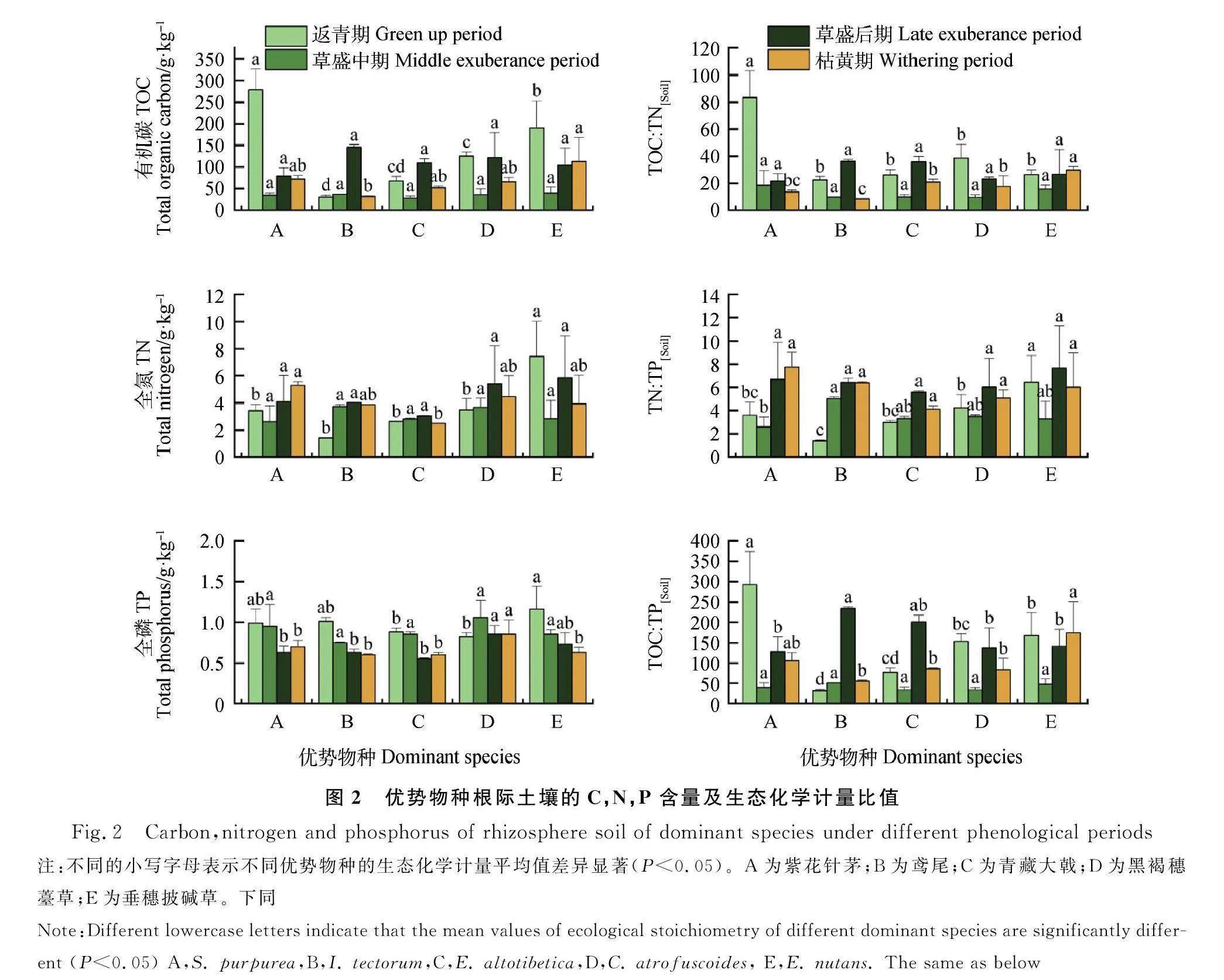
5个优势物种根际土壤的C,N,P 含量及生态化学计量特征如图2所示。在返青期,紫花针茅根际土壤中的TOC含量、C∶N,C∶P最高,垂穗披碱草根际土壤中的TN含量和N∶P最高;在草盛中期,N∶P在各个优势物种根际土壤间差异显著,其余生态化学计量均不显著;在草盛后期,各个优势物种根际土壤的TOC含量、TN含量、C∶N,C∶P均无显著差异;在枯黄期,黑褐穗薹草根际土壤的TP含量显著高于其他优势物种。
2.2 优势物种叶片和根系的生态化学计量特征
随着生长季的推移,所有优势物种的叶片TOC相对稳定,TN整体上呈现出下降的趋势,而C∶N和C∶P整体上呈现出上升的趋势(图3)。在整个生长季节,青藏大戟的TN均显著高于其他优势物种,而C∶N和C∶P均低于其他优势物种,垂穗披碱草的TN低于其他优势物种,但未达到显著水平,N∶P的最小值大部分出现在黑褐穗薹草。在草盛中期和枯黄期,青藏大戟的TP显著高于其他优势物种,在草盛后期,鸢尾的N∶P和C∶P显著高于其他优势物种。
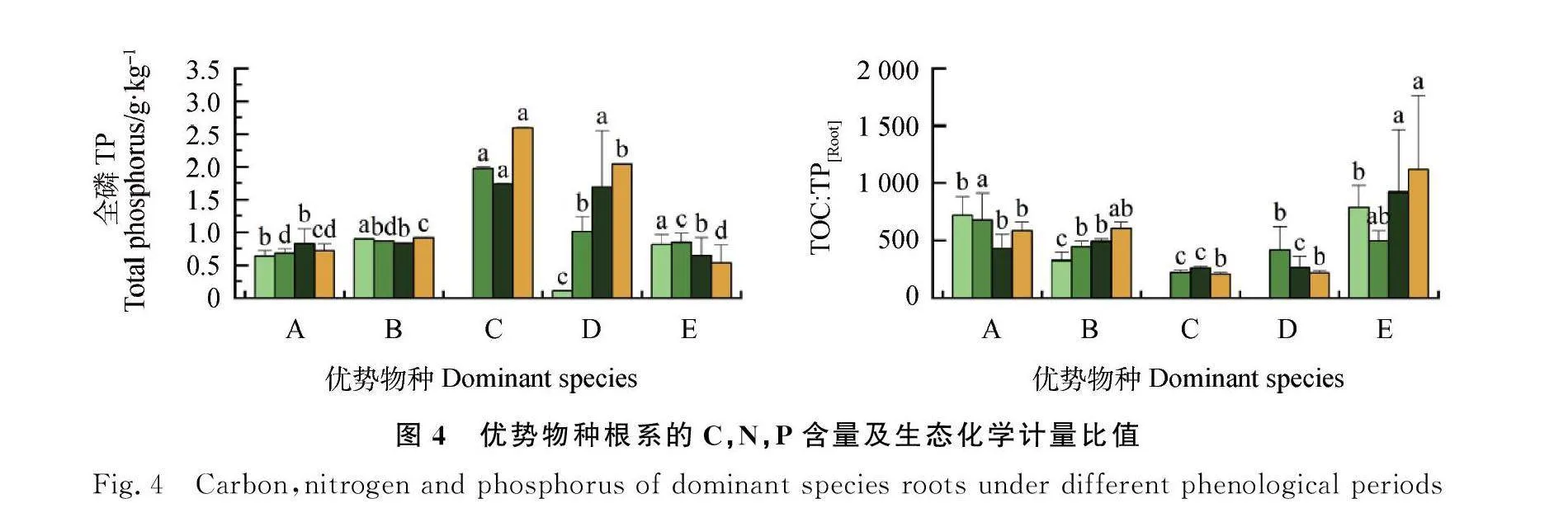
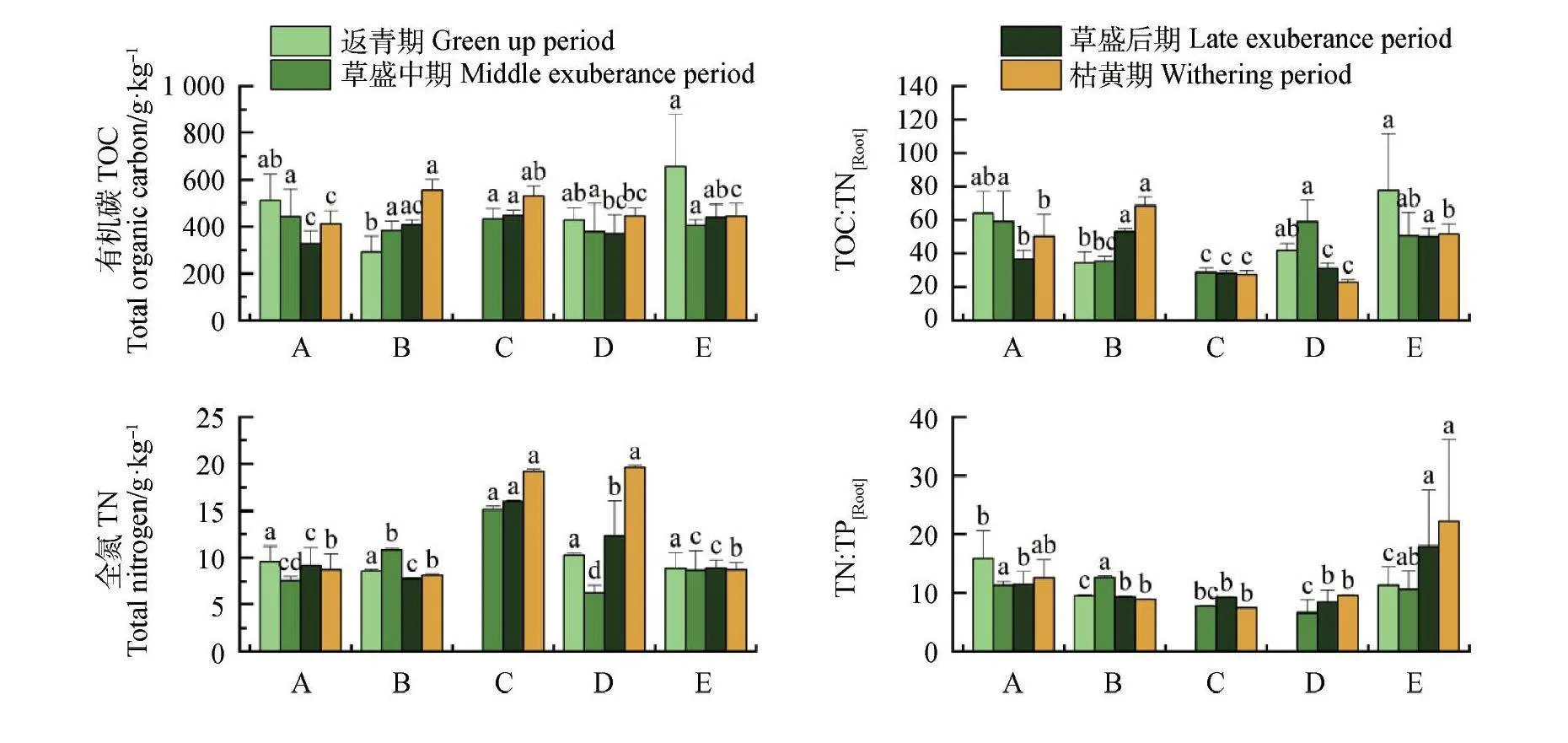
在返青期,根系TOC,C∶N和C∶P的最大值均出现在垂穗披碱草,最小值均出现在鸢尾;在草盛中期,青藏大戟的TN和TP显著高于其他优势物种,紫花针茅的C∶N和C∶P高于其他优势物种,但未达到显著水平;在草盛后期,青藏大戟的TN和TP显著高于其他优势物种,垂穗披碱草的TOC,N∶P和C∶P均显著高于其优势物种;在枯黄期,TOC和C∶N的最大值出现在鸢尾,N∶P和C∶P的最大值出现在垂穗披碱草(图4)。
祁连山优势物种的TOC,TN,TP含量整体上叶片高于根系,而C∶N和C∶P则根系高于叶片。
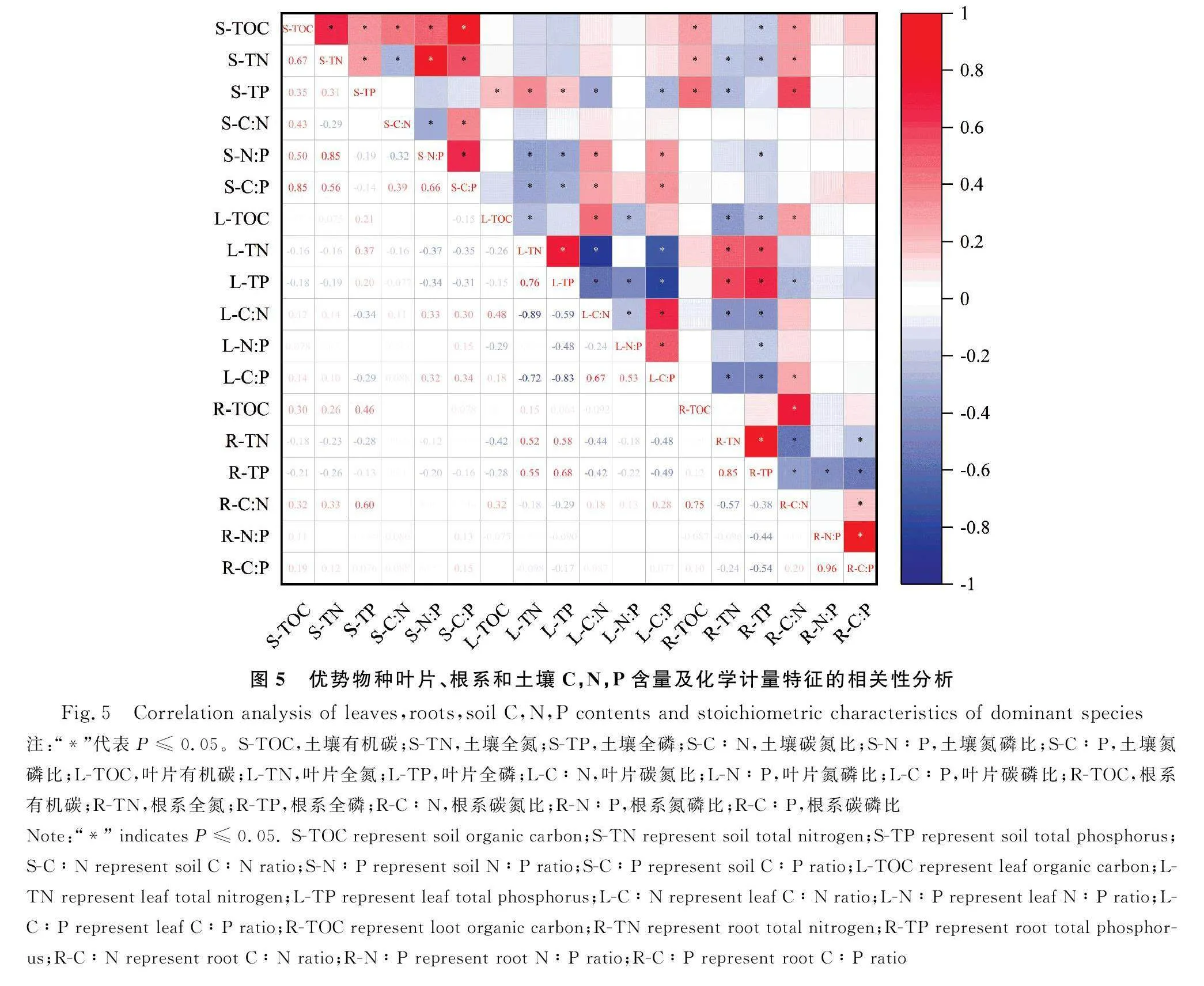
2.3 优势物种与土壤生态化学计量特征的相关性分析
土壤TOC,TN,C∶N与优势物种叶片的生态化学计量无显著相关性(图5);土壤TP与优势物种叶片的TOC,TN,TP均显著正相关(P≤0.05),与C∶N和C∶P显著负相关(P≤0.05);土壤N∶P,C∶P与优势物种叶片的TN,TP显著负相关(P≤0.05),与C∶N,C∶P显著正相关(P≤0.05)。土壤C∶N,C∶P与优势物种根系的生态化学计量无显著相关性;土壤TOC,TN,TP与优势物种根系的TOC,C∶N显著正相关(P≤0.05);土壤TOC与优势物种根系的TP显著负相关(P≤0.05),土壤TN与优势物种根系的TN,TP均显著负相关(P≤0.05),土壤TP与优势物种根系的TN显著负相关(P≤0.05);土壤N∶P与优势物种根系的TP显著负相关(P≤0.05)(图5)。
3 讨论
3.1 祁连山优势物种与中国陆生植物生态化学计量比较
祁连山高寒草地生态系统优势物种叶片(467.40 g·kg-1)和根系(436.63 g·kg-1)的TOC含量均高于中国陆生植物(叶片为436.50 g·kg-1,根系为417.80 g·kg-1)[38-40],表明祁连山的优势物种固碳能力较强。总体上,本研究中5种高寒草地优势物种叶片和根系的TN和TP含量(叶片TN为19.68 g·kg-1,叶片TP为1.66 g·kg-1,根系TN为9.71 g·kg-1,根系TP为0.92 g·kg-1)远高于中国陆生植物(叶片TN为14.16 g·kg-1,叶片TP为1.11 g·kg-1,根系TN为4.87 g·kg-1,根系TP为0.47 g·kg-1),这符合“温度-植物生理假说”(Temperature-Plant physiology hypothesis),即植物生长代谢率受温度调节[41-42]。在寒冷地区,植物的生理过程受到温度的限制,因此相应的温度敏感调节机制被激活,以增加植物的氮和磷含量来补偿生长代谢速率的降低。
植物的生态化学计量比可以反映植物的养分供应情况[10]。本研究的结果表明,植物叶片(30.74和368.00)和根系(48.92和703.31)的C∶N和C∶P均低于中国陆生植物(叶片为30.80和393.20,根系为85.80和888.90)[38-40],这表明祁连山优势物种利用氮磷养分的能力相对较弱。植物N∶P可以用于研究植物物种组成、群落结构、植被生产力等方面的功能特征,通常被用来指示环境的养分供应状况[6,9]。1996年,Koerselman和Meuleman提出了N∶P阈值假说,该假说认为当N∶Plt;14 时,氮是植物生长的限制因子;当N∶P gt; 16 时,磷是植物生长的限制因子;当14lt;N∶Plt;16 时,植物生长受到氮和磷的共同限制[43]。本研究结果表明,祁连山优势物种N 限制比例最高,为76%(N和P共同限制为5%,P 限制为18%),这可能是由于该地区的低温环境直接或间接导致的。在自然条件下,土壤元素的生物地球化学循环过程与生物代谢紧密相关。土壤N的矿化遵循Q10法则,而土壤P的风化却是化学过程,土壤N对温度的敏感度显著高于土壤P。因此,祁连山地区的低温环境可能是导致该地区N限制的原因。其次,“土壤基质年龄假说”也可以解释祁连山地区N限制这一现象,该假说认为高纬度地区的土壤较为年轻,从母岩中释放P的能力较强,而母岩中N含量较低,所以高纬度地区通常为N限制[38]。
3.2 物候期、物种、器官变化对优势物种生态化学计量的影响
植物的养分含量与植物的生存环境密切相关,为了适应不同的环境条件,植被通常会通过生理调控和养分分配过程来调节相应元素的含量[44-45]。本研究表明,随着生长季推移,植物叶片TOC相对稳定,这符合动态平衡理论(内稳态理论),该理论认为植物在长期的进化过程中已经形成了一定的稳态机制,在不断变化的环境中可以基本将体内元素的组成和含量保持在一定水平[46]。随着生长季的推移,植物TN整体上呈现出下降的趋势,这与对盐湖植物的研究结果相似[47]。植物的物质类型包括结构性物质、功能性物质和贮藏性物质,植物在不同的物候条件下生长速度不同,不同的生长需求导致这三种物质的比例不同[48]。在返青期,植物快速生长,其功能性物质被转移到幼嫩组织中,以保证对养分的需求;在草盛中后期,生物量迅速增加,由于养分“稀释效应”[49],植物中的营养物质被稀释,从而减少;在枯黄期,植物的各种功能随着衰老而下降,如酶活性降低、光合和蒸腾等生理作用减弱、营养吸收和同化能力减弱,这些功能的下降均可能导致TN降低[50]。与此同时,优势物种的C∶N和C∶P随着生长季的推移而增加,表明N和P的利用效率逐渐提高[15],这可能是由于与C固定相比,N和P的积累在早期较快,但在后期变慢。
不同的物种具有不同的生态化学计量特征,为了适应不同的外界环境条件,各个物种逐渐进化出适合自身的营养元素吸收和重新分配能力,导致不同优势物种的元素组成和化学计量不同[51]。生长速率假说(GRS)表明生物体的生长速率与其C∶N∶P比率密切相关。一般来说,随着植物生长速率的增加,其体内N∶P和C∶P趋于下降,而P趋于增加[46]。此外,C∶N和C∶P可以表征植物吸收和利用N和P养分同化C的能力,可以通过其大小判断植物对土壤营养元素的利用效率。各个优势物种之间,在大部分物候期下,青藏大戟的TN和TP高于其他优势物种,且C∶N和C∶P低于其他优势物种,而垂穗披碱草的TN低于其他优势物种,且N∶P和C∶P高于其他优势物种,该结果表明青藏大戟(高约20~30 cm)的生长速率相对较快,垂穗披碱草(高约50~70 cm)的生长速率相对较慢。这也符合“异速生长模型”,该模型认为不同生物体在相同的环境条件下,其生长速率是不同的,并且生物体的生长速率与其体积大小成反比,这是因为植物生长需要消耗能量,而能量的消耗与体型大小成正比。因此,体型越大的植物需要消耗更多的能量来维持生命活动,从而导致其生长速率变慢,反之亦然。然而,本研究还缺乏直接数据来支持这些物种的生长速度,未来的研究需要具体分析化学计量对植物生长速率的指示意义。
为了应对不同的胁迫环境,植物经过长期的自然选择,优化了自身资源配置,所以造成同一种植物所含的营养元素在不同器官之间存在差异[52]。祁连山优势物种的TOC,TN,TP含量整体上呈现出叶片高于根系,一方面可能是由于叶片光合作用需大量N和P元素合成叶绿素a和b和蛋白质、核酸及酶[53],因此叶片的N和P元素得到优先分配,另一方面是由于研究区寒冷干旱,抑制了硝酸还原酶活性,导致植物根部对N的吸收降低[54]。
3.3 祁连山优势物种叶片、根系与土壤生态化学计量的关系
植物与土壤是一个相互联系的整体,植物生长所需要的营养元素主要来源于土壤,其含量的高低与土壤中相对应元素的含量高低密切相关[55]。本研究对祁连山地区优势物种和土壤的C∶N∶P生态化学计量特征进行了Person相关性分析,发现其在植物叶片、根系和土壤之间具有相关关系,这表明祁连山地区植物生态化学计量受到土壤化学计量的制约,与以往大多数研究的结果一致,例如通过对委陵菜的研究认为海拔通过影响土壤C和P来影响委陵菜的生态化学计量[56]。
4 结论
本研究采集祁连山高寒草地生态系统的5种优势物种作为研究对象,测定了其叶片和根系的生态化学计量特征,并分析了其与根际土壤生态化学计量特征的关系。结果表明随着生长季推移,优势物种叶片TOC含量相对稳定,TN含量整体上下降,而C∶N和C∶P则增加;叶片的生态化学计量特征结果表明,青藏大戟的生长速率相对较快,垂穗披碱草的生长速率相对较慢;祁连山优势物种的TOC,TN,TP含量整体上表现为叶片高于根系;祁连山地区植物生态化学计量受到土壤化学计量的影响,主要受到N元素的限制。综上,在研究区内,植物的生态化学计量特征不仅与物种本身、物候期、植物器官等息息相关,并且会受到土壤环境的影响。
参考文献
[1] MOE S J,STELZER R S,FORMAN M R,et al. Recent advances in ecological stoichiometry:insights for population and community ecology[J]. Oikos,2005,109(1):29-39
[2] SARDANS J,RIVAS-UBACH A,PEUELAS J. The C∶N∶P stoichiometry of organisms and ecosystems in a changing world:A review and perspectives[J]. Perspectives in Plant Ecology,Evolution and Systematics,2012,14(1):33-47
[3] RONG Q,LIU J,CAI Y,et al. Leaf carbon,nitrogen and phosphorus stoichiometry of Tamarix chinensis Lour. in the Laizhou Bay coastal wetland,China[J]. Ecological Engineering,2015,76:57-65
[4] LIU R,ZHAO H,ZHAO X,et al. Facilitative effects of shrubs in shifting sand on soil macro-faunal community in Horqin Sand Land of Inner Mongolia,Northern China[J]. European Journal of Soil Biology.2011,47(5):316-321
[5] MAO R,CHEN H,ZHANG X,et al. Effects of P addition on plant C∶N∶P stoichiometry in an N-limited temperate wetland of Northeast China[J]. Science of The Total Environment,2016,559:1-6
[6] GSEWELL S. N∶P ratios in terrestrial plants:variation and functional significance[J]. New Phytologist,2004,164(2):243-266
[7] WRIGHT I J,REICH P B,Cornelissen J H C,et al. Assessing the generality of global leaf trait relationships[J]. New Phytologist,2005,166(2):485-496
[8] XIA C,YU D,WANG Z,et al. Stoichiometry patterns of leaf carbon,nitrogen and phosphorous in aquatic macrophytes in eastern China[J]. Ecological Engineering,2014,70:406-413
[9] HE J,WANG L,FLYNN D F B,et al. Leaf nitrogen∶phosphorus stoichiometry across Chinese grassland biomes[J]. Oecologia,2008,155(2):301-310
[10]YAN W,ZHONG Y,ZHENG S,et al. Linking plant leaf nutrients/stoichiometry to water use efficiency on the Loess Plateau in China[J]. Ecological Engineering,2016,87:124-131
[11]GREN G I,WEIH M. Plant stoichiometry at different scales:element concentration patterns reflect environment more than genotype[J]. New Phytologist,2012,194(4):944-952
[12]YU H,FAN J,HARRIS W,et al. Relationships between below-ground biomass and foliar N∶P stoichiometry along climatic and altitudinal gradients of the Chinese grassland transect[J]. Plant Ecology,2017,218(6):661-671
[13]AI Z,HE L,XIN Q,et al. Slope aspect affects the non-structural carbohydrates and C∶N∶P stoichiometry of Artemisia sacrorum on the Loess Plateau in China[J]. Catena,2017,152:9-17
[14]YIN H,ZHENG H,ZHANG B,et al. Stoichiometry of C∶N∶P in the roots of Alhagi sparsifolia is more sensitive to soil nutrients than aboveground organs[J]. Frontiers in Plant Science,2021,12:698961
[15]刘敏国,王士嘉,陆姣云,等. 河西走廊藜麦C、N、P生态化学计量学特征对物候期的响应[J]. 干旱区研究,2018,35(1):192-198
[16]ZHENG S,REN H,LI W,et al. Scale-dependent effects of grazing on plant C∶N∶P stoichiometry and linkages to ecosystem functioning in the Inner Mongolia grassland[J]. Plos One,2012,7(12):e51750
[17]WU T,QU C,LI Y,et al. Warming effects on leaf nutrients and plant growth in tropical forests[J]. Plant Ecology,2019,220(7):663-674
[18]HE M,DIJKSTRA F A. Drought effect on plant nitrogen and phosphorus:a meta-analysis[J]. New Phytologist,2014,204(4):924-931
[19]DU C,WANG X,ZHANG M,et al. Effects of elevated CO2 on plant C-N-P stoichiometry in terrestrial ecosystems:A meta-analysis[J]. Science of The Total Environment,2019,650:697-708
[20]GSEWELL S. Nutrient resorption of wetland graminoids is related to the type of nutrient limitation[J]. Functional Ecology,2005,19(2):344-354
[21]GOTELLI N J,MOUSER P J,HUDMAN S P,et al. Geographic variation in nutrient availability,stoichiometry,and metal concentrations of plants and pore-water in ombrotrophic bogs in New England,USA[J]. Wetlands,2008,28(3):827-840
[22]JAENIKE J,MARKOW T A. Comparative elemental stoichiometry of ecologically diverse Drosophila[J]. Functional Ecology,2003,17(1):115-120
[23]王静,魏小红,龙瑞军. 东祁连山高寒草甸植物抗寒性研究[J]. 草地学报,2007,15(6):537-542
[24]赵林. 青藏高原多年冻土及变化[M]. 北京:科学出版社,2019:7-12
[25]方玉琢. 青藏高原主要草地群落叶氮磷化学计量特征的环境响应[D]. 兰州:兰州大学,2019:26
[26]姚檀栋,刘晓东,王宁练. 青藏高原地区的气候变化幅度问题[J]. 科学通报,2000(1):98-106
[27]ZHANG Y,WANG G,WANG Y. Response of biomass spatial pattern of alpine vegetation to climate change in permafrost region of the Qinghai-Tibet Plateau,China[J]. Journal of Mountain Science,2010,7(4):301-314
[28]阳勇,陈仁升,吉喜斌. 近几十年来黑河野牛沟流域的冰川变化[J]. 冰川冻土,2007(1):100-106
[29]刘婵,刘冰,赵文智,等. 黑河流域植被水分利用效率时空分异及其对降水和气温的响应[J]. 生态学报,2020,40(3):888-899
[30]梁冰妍,徐海燕,吴晓东,等. 祁连山不同草地类型区土壤有机碳组份的差异[J]. 地球科学,2024,49(4):1487-1497
[31]李莉莎,徐海燕,吴晓东,等. 青藏高原高山嵩草叶、根抗寒性生理特征[J]. 草地学报,2020,28(6):1544-1551
[32]韩发,岳向国,师生波,等. 青藏高原几种高寒植物的抗寒生理特性[J].西北植物学报,2005,25(12):2502-2509
[33]杜坤,李金萍,王婷,等. 转抗草甘膦基因甘蓝型油菜根际土壤理化性质及真菌群落多样性[J]. 生态学杂志,2024,43(4):1082-1091
[34]杨琼,谭凤仪,吴苑玲,等. 不同林龄海桑林和无瓣海桑林根际微生物特征[J]. 生态学杂志,2014,33(2):296-302
[35]鲍士旦. 土壤农化分析[M]. 北京:中国农业出版社,2000:25-97
[36]NELSON D W,SOMMERS L E. Total Carbon,Organic Carbon,and Organic Matter[M]. Methods of Soil Analysis,1996:961-1010
[37]OLSEN S R. Estimation of available phosphorus in soils by extraction with sodium bicarbonate[M]. Washington:USDA Circular,1954:18-19
[38]REN S,YU G,TAO B,et al. Leaf nitrogen and phosphorus stoichiometry across 654 terrestrial plant species in NSTEC[J]. New Phytologist,2007,28(12):2665-2673
[39]HAN W,FANG J,GUO D,et al. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China[J]. New Phytologist,2005,168(2):377-385
[40]TANG Z,XU W,ZHOU G,et al. Patterns of plant carbon,nitrogen,and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems[J]. Proceedings of the National Academy of Sciences,2018,115(16):4033-4038
[41]REICH P B,OLEKSYN J. Global patterns of plant leaf N and P in relation to temperature and latitude[J]. Proceedings of the National Academy of Sciences,2004,101(30):11001-11006
[42]WOODS H A,MAKINO W,COTNER J B,et al. Temperature and the chemical composition of poikilothermic organisms[J]. Functional Ecology,2003,17(2):237-245
[43]KOERSELMAN W,ARTHUR F M M. The vegetation N∶P ratio:a new tool to detect the nature of nutrient limitation[J]. Journal of Applied Ecology,1996,33(6):1441-1450
[44]MLLER M,OELMANN Y,SCHICKHOFF U,et al. Himalayan treeline soil and foliar C∶N∶P stoichiometry indicate nutrient shortage with elevation[J]. Geoderma,2017,291:21-32
[45]XU X,QIN Y,CAO J J,et al. Elevational variations of leaf stochiometry in Leontopodium leontopodioides on the Qinghai-Tibetan Plateau,China[J]. Chinese Journal of Applied Ecology,2018,29(12):3934-3940
[46]STERNER R W,ELSER J J. Ecological stoichiometry:the biology of elements from molecules to the biosphere[M]. New Jersey:Princeton University Press,2002:262
[47]WANG X,L X,DIJKSTRA F A,et al. Changes of plant N∶P stoichiometry across a 3000 km aridity transect in grasslands of northern China[J]. Plant and Soil,2019,443(1):107-119
[48]牛得草,董晓玉,傅华. 长芒草不同季节碳氮磷生态化学计量特征[J]. 草业科学,2011,28(6):915-920
[49]STRIEBEL M,SPRL G,STIBOR H. Light-induced changes of plankton growth and stoichiometry:Experiments with natural phytoplankton communities[J]. Limnology and Oceanography,2008,53(2):513-522
[50]孙书存,陈灵芝. 东灵山地区辽东栎叶养分的季节动态与回收效率[J]. 植物生态学报,2001,25(1):76-82
[51]SARDANS J,PEUELAS J. The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system[J]. Plant Physiology,2012,160(4):1741-1761
[52]CAO Y B,WANG B T,WEI T T,et al. Ecological stoichiometric characteristics and element reserves of three stands in a closed forest on the Chinese loess plateau [J]. Environmental Monitoring and Assessment,2016,188(2):80
[53]刘万德,苏建荣,李帅锋,等. 云南普洱季风常绿阔叶林优势物种不同生长阶段叶片碳、氮、磷化学计量特征[J]. 植物生态学报,2015,39(1):52-62
[54]罗海斌,黄诚梅,朱慧明,等. 干旱胁迫对甘蔗根系碳氮代谢的影响[J]. 南方农业学报,2020,51(6):1332-1338
[55]HERBERT D A,WILLIAMS M,RASTETTER E B. A model analysis of N and P limitation on carbon accumulation in Amazonian secondary forest after alternate land-use abandonment[J]. Biogeochemistry,2003,65(1):121-150
[56]QIN Y Y,LIU W,ZHANG X F,et al. Leaf stoichiometry of Potentilla Fruticosa across elevations ranging from 2400 m to 3800 m in China’s Qilian Mountains (Northeast Qinghai-Tibetan Plateau)[J]. Frontiers in Plant Science,2022,13:814059
(责任编辑 闵芝智)
