
寒温带针叶林优势树种倒木分解真菌群落多样性及功能差异
2023-12-25 14:27:28崔福星杨立宾付晓宇朱道光
生态学报 2023年23期
潘 虹,崔福星,杨立宾,魏 丹,付晓宇,朱道光
黑龙江省科学院自然与生态研究所,哈尔滨 150040
倒木是林木生长发育过程中由于老龄化、种间竞争、自然或人为干扰造成死亡的一种粗木质残体(CWD)。倒木分解深刻影响了森林生态系统的碳库储存、群落更新、养分循环等关键过程,是林地重要的生态功能单元,在维持和稳定森林生态系统的生物地球化学循环方面具有重要作用。
近年来,倒木分解逐渐成为森林生态学研究热点领域之一,内容多集中于分解过程的生态功能[1],主要包括倒木养分的分解及释放动态[2]、木腐机制及功能菌群的降解机理[3]、分解过程的呼吸特征及呼吸贡献[4]、土壤养分输入中的碳循环对气候变化的响应[5—6]以及森林更新[7]等方面,而针对倒木分解过程中微生物群落组成及功能方面还缺乏深入探讨。树种的差异是如何影响倒木分解过程中真菌群落结构和生态功能的?前人研究发现,倒木分解是小型动物蚕食、微生物侵入、自然淋溶和风化破碎交互作用的结果,微生物作为倒木的分解者之一,可将木质中的大分子有机底物腐解,促使倒木结构和质量发生改变[8]。与此同时,倒木通过异养呼吸、生物转化与浸出等过程释放多种营养物质,成为微生物的养分来源[9]。另外,有研究表明,在森林小尺度生境内,不同树种倒木的化学结构和物理性状存在差异,随着腐殖化进程的加剧,这种差异不仅吸引着具有不同生态功能微生物的生长繁殖,而且与真菌群落生物量之间存在定向关系。王越[10]、蔡琳颖[11]、张铁等[12]在香樟、红楠、甜槠、三角槭、狭叶润楠等分解初期的研究中发现,倒木自身质量、含水率、孔隙结构、木质残体碳结构、含氮量等因素与真菌群落多样性之间存在密切关联,森林群落类型、树种影响了环境真菌的群落组成及分布规律。
大兴安岭北部地区是以寒温带针叶林作为地带性植被的北方森林生态系统,区内辅以兴安落叶松、白桦、樟子松为主要的建群种或优势种,是全球生态变化的重要敏感区。目前,全球范围有关寒温带中高纬度森林倒木分解过程中真菌群落变化的相关研究并不广泛,优势树种倒木的养分差异及其对微生物群落带来的动态影响方面的研究还不够深入。因此,本文在大兴安岭北部高山林区开展白桦(BetulaplatyphyllaSuk.)、兴安落叶松(Larixgmelinii(Rupr.)Kuzen)、樟子松(Pinussylvestrisvar.mongolicaLitv.)3种优势树种倒木分解过程中真菌群落多样性及功能差异等方面的研究,试图探究(1)不同优势树种倒木真菌群落的多样性及功能变化;(2)影响倒木真菌群落分布的驱动因子。研究结果为进一步阐述寒温带森林生态系统中微生物参与的倒木分解、物质返还及对土壤碳库的贡献提供理论依据。
1 材料和方法
1.1 试验区设置及样品采集
研究地点位于黑龙江省大兴安岭呼中国家自然保护区(51°49′01″—51°49′19″N,122°59′33″—123°00′03″E),该地点地形平缓,海拔847—974 m,年均气温-4℃,年均降水量458.3 mm,年均相对湿度71%,年均蒸发量911 mm[13]。该地点共有平均胸径>1 cm的木本植物41种,其中乔木4种,灌木37种,草本127种,隶属于21科39属[14]。
试验样地内设置20 m×20 m样方,样方内要求以白桦(下文简称Bp)、兴安落叶松(下文简称Lg)、樟子松(下文简称Ps)为主要的建群种或先锋树种 。 2018年6月分别在处于生长期的白桦、落叶松、樟子松上折取长约50 cm、直径约2 cm的健康枝条若干,将枝条均匀散布于样方内地表,一年后原位回收各倒木枝条样品3根,经封装、标记后带回实验室,刮取土壤接触面处枝条周皮5 g,用于真菌的DNA提取,每样品3次重复。
1.2 分解倒木中真菌DNA的提取、扩增及建库
利用E.Z.N.A.® Soil DNA Kit(D5625-01,Omega,美国)提取白桦、兴安落叶松及樟子松凋落枝条样品总DNA,采用1%的琼脂糖凝胶电泳检测DNA的提取质量,使用超微量分光光度计(ND-2000,Thermo,美国)检测DNA浓度约10 ng/μL即可,电泳条带均在20 kb以上,具有完整性,满足扩增要求。
利用GeneAmp® 9700型聚合酶链式反应基因扩增仪(PCR仪)(9700,ABI,美国),使用引物SSU0817F(5′-TTAGCATGGAATAA TRRAATA GGA-3′)和1196R(5′-TCTGGACCTGGTGAGTTTCC-3′)对真菌内源转录间隔区(ITS)基因序列进行扩增。反应体系:5×FastPfu Buffer 4 μL,dNTPs(2.5 mmol/L)2 μL,Forward Primer(5 μmol/L)0.8 μL,Reverse Primer(5 μmol/L)0.8 μL,FastPfu Polymerase 0.4 μL,BSA 0.2 μL,Template DNA 10 ng,补充ddH2O至反应体系20 μL。反应条件:95℃预变性3 min,30个循环(95℃变性30 s,55℃退火30 s,72℃延伸45 s),然后72℃稳定延伸10 min,最后10℃直至停止。产物4℃保存,每样本3次重复。
将同一倒木样品的3次PCR产物混合后,首先,使用2%琼脂糖凝胶3 μL上样电泳检测,切胶回收扩增产物,利用AxyPrep DNA Gel Extraction Kit(AP-GX-250,Axygen Biosciences,美国)对产物进行Tris-HCl洗脱纯化;其次,使用2%琼脂糖凝胶二次电泳检测,并使用QuantiFluorTM-ST蓝色荧光定量系统(Promega,美国)对回收产物进行检测定量;最后,使用NEXTflexTMRapid DNA-Seq Kit(5144-08,Bioo Scientific,美国)建立MiSeq文库。
1.3 高通量测序和数据处理
利用Illumina公司的MiSeq PE 300平台对9份样本的PCR扩增产物进行双端测序,下机后对序列数据进行质量控制,得到优化序列。优化序列的数据信息上传至美国国立生物技术信息中心序列阅读档案(NCBI SRA)数据库(序列号:SRP360510)。
使用Uparse(Version 11)[15],将得到的优化序列在97%的相似度水平下进行聚类,同时剔除嵌合体,根据最小样本序列数对样本序列进行抽平,获得OTU及代表序列。基于OTU分类水平样本表,首先,使用R(Version 3.3.1)[16],统计不同分组中OTU数目并制作Venn图,以展现不同树种倒木分解中真菌OTU的组成及重叠情况。其次,使用Mothur(Version 1.30.2),计算不同分组下Shannon指数,反映真菌群落的Alpha多样性;Sobs指数反映实际观测的物种数目;Chao1指数为估计的物种数目,反映真菌群落的丰富度。
基于Unite(Release 8.0)[17]ITS真菌分类学数据库,采用RDP classifier(Version 2.13)对OTU的代表序列进行分类学注释,设置序列比对的置信度阈值为0.7,得到每个OTU在各物种分类水平对应的物种分类信息。基于真菌群落的分类信息及丰度数据,使用R,采用One-way ANOVA方法检验各物种在不同分组中的丰度差异,分别在门、属水平上绘制柱形图,以展示不同倒木分组内的物种丰度分布及差异显著性;使用R Ternary包绘制三元相图,展示优势真菌在不同倒木分组间的分布比例。
使用R,基于不同分组下OTU、门、属水平上的真菌多样性数据,将物种的差异和丰度作为权重,基于Bray-Curtis距离算法、Adonis组间差异检验方法对分类数据进行主坐标分析(PCoA)[18]。此外,采用Pearson相关性算法、Bray-Curtis距离算法得到分组样本的距离矩阵。通过绘制PCoA图和组间距离矩阵图,比较分析不同倒木分解中真菌群落的Beta多样性。
使用R,在群落方面,通过Mantel tests分析检验倒木养分矩阵与门、属水平上真菌群落矩阵的相关关系;在物种方面,通过计算养分指标与真菌各物种之间的Pearson相关系数及差异显著性P值,获得相关性数值列表,以评估不同群落、物种与倒木养分变量之间的关系。
使用Fungal traits[19]真菌功能性状数据库,对样本中真菌群落进行功能注释,绘制条形图展示不同倒木的主要真菌生态类型。
1.4 养分指标的测定方法
采用微量凯氏定氮法测定倒木样品全氮含量;重铬酸钾容量法测定全碳含量;酸度计法检测样品pH值;烘干称重法(60℃,48 h)测定样品含水率;96孔板酶标法测定纤维素、半纤维素、木质素含量。
2 结果和分析
2.1 不同树种倒木分解中真菌群落多样性比较
序列经质控后得到356300条优化序列,经聚类后得到59个OTU。从Venn(图1)可以直观地表现出白桦(Bp)、兴安落叶松(Lg)、樟子松(Ps)倒木组间OTU的比例及重叠情况,其中,Bp、Lg、Ps的真菌OTU占总OTU比率分别为77.97%、83.05%、61.02%,三组共有OTU占比52.54%,独有OTU占比分别为15.25%、15.25%、0。可见,落叶松倒木分解过程中真菌种类最多,白桦次之,樟子松最少,仅存在与白桦或落叶松倒木上的独有真菌数量相同,樟子松没有独有真菌。

图1 基于OTU水平不同树种倒木分解的真菌Venn图
如表1,Coverage表示真菌物种文库的覆盖率,3种倒木样本中真菌序列被测出的概率均达到0.99以上,可以反应分解倒木上真菌的真实情况,在此前提下可开展不同树种倒木分解过程中真菌群落Alpha多样性指数变化及分析。

表1 不同树种倒木分解中真菌群落Alpha多样性指数
在分解的倒木中,Lg、Bp、Ps 树种真菌群落的Shannon指数逐渐降低,说明3种倒木样品中兴安落叶松分解倒木中真菌群落具有较高的物种丰度或均匀度,Alpha多样性最高,白桦次之,樟子松最低。另外,从实际观测到的物种数目来看(Sobs指数表征),Lg倒木真菌数量最多,Bp次之,Ps最少。与Sobs指数相比,Chao1指数对稀有物种更为敏感,可表征物种的丰富度,下表可见,Lg倒木Chao1指数最高,Bp次之,Ps最少,说明兴安落叶松分解倒木中不仅观测到的物种数目最多,同时也具有稀有物种的相对优势。差异性检验表明,Shannon、Sobs与Chao1指数在Lg与Ps组间均呈现显著差异,Bp与Lg组间、Bp与Ps组间均无显著差异,可见,在3种不同树种倒木分解过程中,兴安落叶松、樟子松上真菌群落的Alpha多样性差异显著,白桦与其它树种的Alpha多样性尽管存在一定程度差异,但并不显著。
2.2 主要真菌群落在不同树种倒木分解中的组成及分布
图2柱状图所示,门水平上,大兴安岭林地3种倒木上共检测出2个已知真菌门,分别为Ascomycota(子囊菌门)、Basidiomycota(担子菌门),其中Ascomycota为Lg组内优势菌门,Basidiomycota为Bp、Ps组优势菌门,且Ascomycota菌门的相对丰度在不同倒木分组中呈现显著差异,Basidiomycota呈极显著差异。

图2 基于门分类水平下倒木分解的真菌群落组成与分布
图3柱状图所示,属水平上,相对丰度0.01以上的真菌包括附毛菌属(Trichaptum)、Acidea、伞菌纲未分类属(Agaricomycetes unclassified)、子囊菌门未分类属(Ascomycota unclassified)、肉座菌目未分类属(Hypocreales unclassified)、散囊菌纲未分类属(Eurotiomycetes unclassified)、Cryptotrichosporon等10个菌属,其中Trichaptum为Bp、Ps优势菌属,Acidea为Lg优势菌属,且Trichaptum菌属的相对丰度在不同倒木分组中呈极显著差异,Acidea呈显著差异。

图3 基于属分类水平下倒木分解的真菌群落组成与分布
图2、图3中三元相图展示了白桦、兴安落叶松、樟子松倒木对主要真菌类群的富集特征。结果表明,不同倒木样本中真菌群落的组成和分布比例有所不同,其中,Ascomycota菌门,Acidea、Hypocreales unclassified、Basidiomycota unclassified菌属主要分布于Lg;Basidiomycota菌门,以及Trichaptum、Cryptotrichosporon菌属主要分布于Ps;Agaricomycetes unclassified、Ascomycota unclassified菌属主要分布于Bp,各主要真菌类群在优势组别的相对丰度在0.05—0.96之间,体现了微生物在不同分解倒木中的分布差异性。
2.3 不同树种倒木分解中真菌群落组间差异性分析
图4所示,在门水平上,Bp、Ps组间相对距离为0.907,群落相似程度较高,Bp、Lg组间、Lg、Ps组间相对距离分别为-0.569、-0.171,群落差异性较大,但整体来看,Bp、Lg、Ps 3组倒木真菌群落组间的置信椭圆重叠性相对较高,P值为0.068,组间无显著性差异。在属、OTU水平上,Bp、Lg、Ps组间置信椭圆重叠性较低,相对距离分别在0.194—0.513、0.134—0.339之间,P值分别为0.005、0.001,说明不同树种倒木在属和OTU分类水平上的真菌群落均具有较低的相似性,组间呈现极显著差异。可见,不同树种在属、OTU水平上极显著改变了倒木上真菌群落的Beta多样性,间接说明OTU这种假定的分类层次在一定程度上客观地反应了属水平上体现的物种群落信息。

图4 不同树种倒木分解的真菌群落PCoA分析及相关性矩阵
2.4 倒木分解中真菌群落主要生态类型
图5可见,大兴安岭林地3种树种倒木上真菌群落的主要生态类型为腐生型(Saprotroph)、病原型(Pathotroph)以及共生型(Symbiotroph),其中,腐生是Bp、Lg、Ps倒木上优势真菌的主要类型。同一生态类型下主要真菌类群存在不同的生活方式、营养特征及宿主,Trichaptum为木质腐生(Wood saprotroph),Acidea为土壤腐生(Soil saprotroph),Capnobotryella、Cystobasidiopsis等为未确定腐生(Undefined saprotroph),Cryptotrichosporon为植物病原(Plant pathogen),Anzia为地衣共生(Lichen symbiotroph)。此外,不同倒木上真菌群落的生态类型分布不同,Bp、Ps真菌主要以木质腐生为主,其次为土壤腐生、植物病原及地衣共生等;Lg真菌则以土壤腐生为主,其次为木质腐生及植物病原。综上所述,腐生型是大兴安岭白桦、兴安落叶松、樟子松倒木上真菌群落的主要生态类型,优势腐生真菌的生活方式及营养特征在不同树种间具有一定的差异性。

图5 不同树种倒木真菌群落的生态类型
2.5 不同树种倒木养分差异及其与真菌群落的关联分析
表2所示,养分指标中,TC、TN、纤维素、半纤维素、木质素的质量分数在3种倒木中由高到低依次为Ps、Lg、Bp,MC、pH则以Bp最高,Lg次之,Ps最低,可见,不同养分指标的含量在3种倒木间呈现出一定的规律性。另外,在统计分析上,倒木基质的pH、TC、TN、C/N、半纤维素、木质素在Bp、Lg、Ps不同树种间均存在显著性差异。
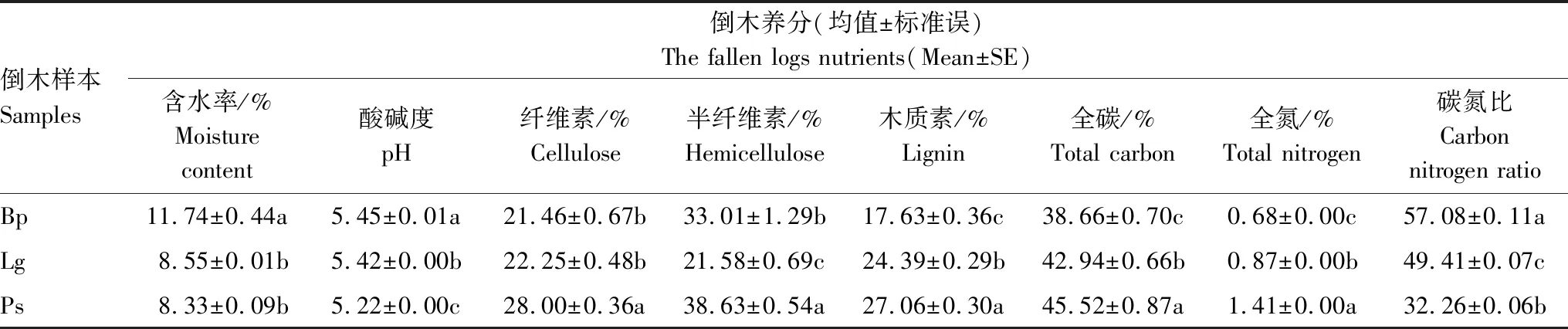
表2 不同树种倒木养分
表3中Mantel测试分析显示倒木养分指标与属水平上真菌群落之间相关性更高(r=0.432,P=0.002),说明腐朽过程中养分指标主要改变属水平各物种的组成和分布。其中,通过Pearson相关分析发现,MC与Ascomycota unclassified之间呈现极显著正相关;纤维素、半纤维素、TN与Trichaptum呈极显著或显著正相关;纤维素、TC、TN与Cryptotrichosporon呈显著正相关性。另外,尽管养分指标与门水平上真菌群落相关性较低(r=0.170,P=0.107),但纤维素、半纤维素与Basidiomycota呈现的正相关性验证了担子菌门物种在倒木腐解初期的重要作用。总之,通过多种相关分析,推测TN、TC、MC、纤维素、半纤维素是影响倒木上优势真菌群落分布的主要养分因素,对Basidiomycota、Trichaptum、Cryptotrichosporon、Ascomycota unclassified等菌群可能产生的富集作用,有待进一步证实。
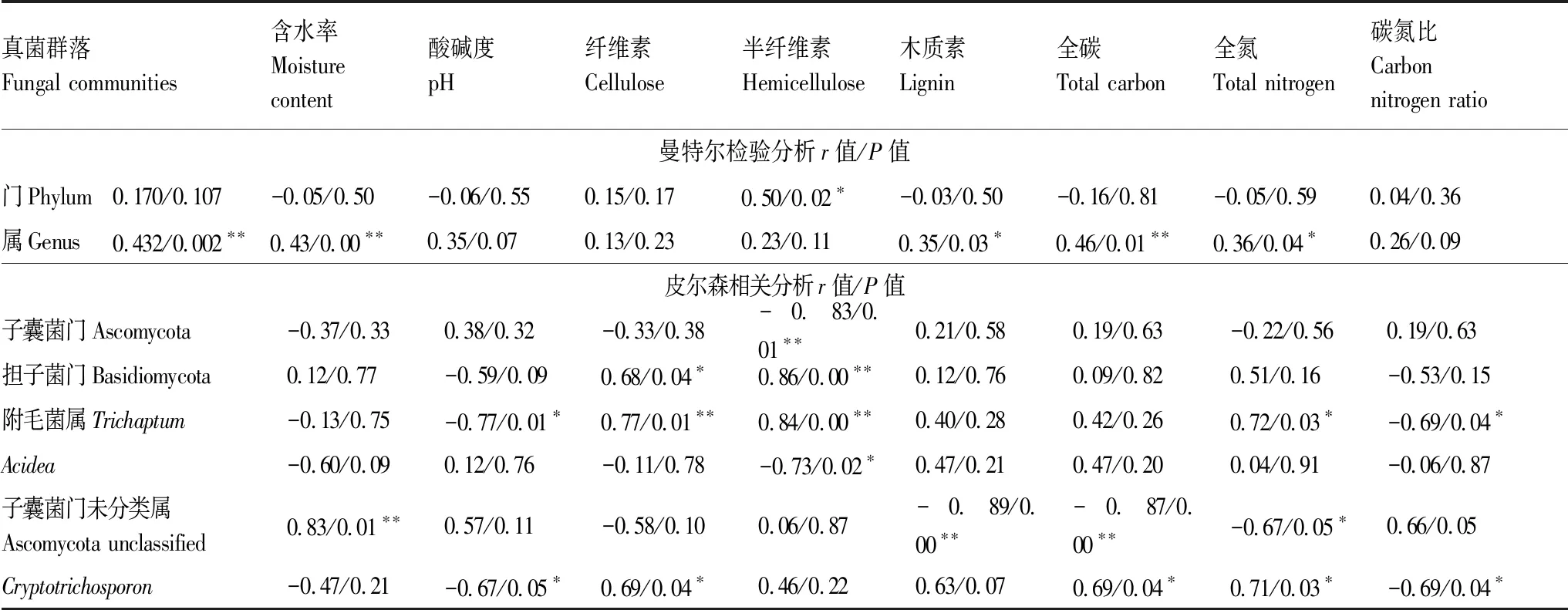
表3 倒木养分与真菌群落的相关性分析
3 讨论
3.1 不同树种倒木真菌群落多样性变化
众多研究表明,树种类型是影响倒木真菌群落结构的重要因素之一[11,20]。崔宝凯等对不同腐朽乔木真菌的聚类分析发现,大兴安岭落叶松属倒木上的真菌种类和数量明显高于本地云杉属和杉属[21]。王越等对亚热带典型树种与微生物群落相关研究得出,落叶阔叶树种枫香、三角槭以及常绿阔叶树种香樟、红楠倒木上真菌群落组分发生明显改变[10]。本研究对林地优势树种倒木的真菌群落多样性分析表明,与白桦、樟子松相比,兴安落叶松倒木真菌群落具有较高的物种丰富度和均匀度,多样性指数最高,树种的差异显著改变了倒木真菌群落的Alpha多样性。造成这一结果的原因,一方面是由树种自身不同的化学和结构特性对真菌群落的影响决定的,另一方面是由林分中树种的不同覆盖度导致的寄主差异决定的,即兴安落叶松属作为区域原始天然林的建群种,拥有富集真菌的相对优势。
在倒木真菌群落组成上,多项研究表明,在Ascomycota、 Basidiomycota、 Mortierellomycota(被孢霉门)等菌门,以及Trichaptum、Oligoporussericeomollis(柔丝干酪孔菌)、Daedaleopsistricolor(三色拟迷孔菌)等菌属为寒温带落叶松林、阔叶红松林、针阔混交林优势树种倒木上的主要真菌类群[22—23]。本研究中,Ascomycota、Basidiomycota菌门以及Trichaptum、Acidea菌属是白桦、兴安落叶松、樟子松倒木分解的优势菌群,这与上述他人研究结果部分一致。可见,在北方森林生态系统中,尽管林分不同,但Basidiomycota、Ascomycota、Trichaptum是主导倒木初期分解的主要真菌类群。此外,在林地小环境下,样地原位土壤真菌类型对倒木的群落组成产生重要的影响。杨立宾、隋心等在与本文同一样地进行土壤真菌群落的研究中认为,大兴安岭落叶松林样地土壤中存在Ascomycota、Basidiomycota、Zygomycota(接合菌门)、Chytridiomycota(壶菌门)、Mucoromycota(毛霉门)等优势菌群[13, 24],其中原位土壤中的Ascomycota、Basidiomycota菌门与腐朽倒木优势菌群一致,推测这些真菌类群在分解初期可能会突破土壤基质向倒木接触面延伸,成为了腐解过程的先驱者。
除此以外,本研究在呼中国家自然保护区监测样地3种倒木上共检测出2个已知真菌门,10个丰度0.01以上菌属,这与前人在丰林自然保护区针叶林倒木上检出的主要真菌70属[22]、官山自然保护区常绿阔叶林倒木真菌16种属[25]、喀斯特常绿落叶阔叶混交林倒木31属[12]相比甚少。究其原因可能为,本研究中倒木属于Renvall[26]粗木质残体分解等级中的腐烂Ⅱ级,木质处于腐烂初期,基质表面腐解程度低,残体较硬,溃孔较小,对多种木腐真菌未起到较好的富集作用。此外,我国森林生态系统气候多样,相较热带、亚热带和温带森林,本研究样地属于中国寒温带最高纬度林区,气温较低,树种稀少,从而造成林地倒木真菌种类相对较少。
3.2 倒木养分及腐殖化差异对真菌群落的影响
有研究表明,同一环境,不同树种间因自身理化特性及养分释放规律的差别导致倒木腐殖化存在较大差异[27—28]。尽管本研究不同树种倒木间的TC、TN、半纤维素、木质素等养分指标存在显著性差异,但因不同乔木基质的初始质量不同,养分含量无法作为判断倒木腐殖进程的依据。
有研究证实,N是真菌利用胞外酶分解倒木的重要元素[29],养分含量是衡量植物残体分解速率的重要计量指标[30],较高的TC、TN、纤维素、半纤维素、木质素以及较低的C/N更利于植物残体的分解[31]。本研究中樟子松倒木TC、TN、木质纤维素含量最高,C/N最低,据此推测樟子松倒木可能是寒温带森林优势乔木初期分解进程最快、腐殖化程度最高的树种。另外,康玲等学者对大兴安岭原始林中倒木分解的研究表明,兴安落叶松倒木中的C、N、木质纤维素等养分含量显著大于白桦,与本研究结果一致。该研究还认为,兴安落叶松倒木的分解常数为0.0121/a(R2=0.9715),白桦为0.2859/a(R2=0.8915),白桦倒木的分解速率明显高于兴安落叶松[32]。倘若本研究通过养分指标比对(表2所示)得出兴安落叶松倒木分解速率高于白桦,则与上述康玲的研究结果相反。除了上述TC、TN、木质纤维素等养分指标外,含水率也是影响倒木呼吸的重要因子[10],本研究中白桦倒木含水率显著高于兴安落叶松,在一定的阈值内较高的储水条件会增加倒木表面溃孔,提高透气性,从而加速白桦分解。可见,造成不同树种倒木腐殖化差异的因素复杂多样,学者应进行多角度比较,尽可能得到合理的综合分析结论。
真菌是腐朽倒木的重要分解者,倒木养分、腐殖化等级与真菌群落的变化息息相关[33]。有研究表明,N含量是影响倒木分解的重要指标,尽管可溶性有机C、N因自然作用不断淋失,但倒木的分解增大了微生物的N固持,从而导致分解初期N含量升高[34]。还有研究表明,较高的N 含量不仅可以加速倒木的分解进程,也有利于营腐生菌群的代谢繁殖,与真菌群落存在正相关关系[35]。此外,在倒木分解初期,自然淋溶及分解者的呼吸作用促进了部分纤维素的解聚,从而造成有机碳、纤维素、半纤维素含量逐渐下降,木质素含量逐渐上升[36—37]。本研究倒木养分与真菌群落的关联分析也证实,在门水平上,Basidiomycota与TC、TN、纤维素、半纤维素含量存在正相关性,这与崔福星等研究者在大兴安岭落叶松倒木研究中得出的结论一致[23]。
木腐真菌是主导倒木分解的重要微生物类群,包括偏好纤维素、半纤维素水解的褐腐真菌,以及偏好木质素降解的白腐真菌[38]。本研究中,Trichaptum为属水平上3种倒木的优势菌群,也是白腐真菌的主要类群之一,从热带到温带森林腐朽木质上均有不同种类发生,由于倒木样本属于腐烂Ⅱ级,处于分解初期,该时期木质残体中大部分菌群未经木腐条件的长期驯化,功能性分化不明显,Trichaptum并未展现出木质白腐特性。李俊凝等学者在木腐真菌多样性与寄主倒木关系的研究中,认为腐烂Ⅱ级倒木与白腐真菌多度具有极显著相关性[22],这与本研究中的Trichaptum为一年期倒木优势菌群结果一致,说明Trichaptum菌属可能是大兴安岭寒温带森林倒木腐朽初期的主要分解者。此外,在关联分析中,Trichaptum菌属展现了与纤维素、半纤维素极显著的正相关性。有研究表明,白腐真菌通过分泌多种水解酶降解木材中的部分纤维素、半纤维素和木质素等大分子多糖物质,促使木质丧失纤维结构而腐朽[28],反之,这些多糖物质对倒木优势真菌的影响如何?目前,树种特性对真菌的影响主要是基于群体数量进行统计,纤维素、半纤维素及木质素对优势菌群的影响研究相对匮乏,急需开展条件控制实验进行深入探讨,以补充倒木寄主特性对真菌群落分布及生态功能的影响。
4 结论
大兴安岭呼中国家自然保护区寒温带森林优势树种倒木上共检测出2个真菌门、10个真菌属(相对丰度≥0.01),优势门为Ascomycota、Basidiomycota,优势属为Trichaptum、Acidea,各优势菌群的相对丰度在不同倒木中差异显著;倒木组内真菌群落Alpha多样性由高到低依次为兴安落叶松、白桦、樟子松,树种差异极显著的改变了OTU、属水平上倒木组间真菌群落的Beta多样性;腐生型是3种倒木上优势真菌的主要生态类型,木质腐生、土壤腐生为真菌类群的主要生活方式;推测TN、TC、MC、纤维素、半纤维素可能是影响倒木真菌群落分布的主要因素,对Basidiomycota、Trichaptum、Cryptotrichosporon等菌群产生的富集作用有待进一步证实。
猜你喜欢
《学习方法报》小学语文四年级(2023年26期)2023-04-29 00:44:03
今日农业(2021年10期)2021-11-27 09:45:24
文苑(2019年20期)2019-11-20 02:12:33
文苑·经典美文(2019年10期)2019-10-16 02:47:16
山东林业科技(2019年2期)2019-06-03 10:10:54
现代营销(创富信息版)(2018年2期)2018-08-15 00:45:27
现代园艺(2018年2期)2018-03-15 08:00:09
现代园艺(2018年2期)2018-03-15 08:00:08
北极光(2018年12期)2018-03-07 01:01:58
河南科技(2014年5期)2014-02-27 14:08:50
