
极小种群闽桦生殖物候及种子萌发影响因素研究
2023-12-18 10:52陈爱平
中国农学通报 2023年32期
陈爱平
(福建省沙县官庄国有林场,福建三明 365050)
0 引言
闽桦是福建省特有乡土阔叶树种,发现于三明罗卜岩保护区桦木科(Betulaceae)桦木属(Betula)的新树种,该树种树干笔直,高可达40 m,少分枝,幼树树皮与亮叶桦(Betulaluminifera)相似,老树树皮呈纵裂[1],与光皮桦呈明显区别,2020年列入福建省特有树种极小种群目录。闽桦材性中等偏硬,具有较好的机械加工性能[2],为林区农民喜好用材树种,保护区成立前其天然资源被大量砍伐。目前仅在闽西北闽桦有少量零星分布,仅在罗卜岩保护区有小范围天然片林,且林分以大径阶老树为主,幼树极少,自然更新十分困难,属于极度濒危树种[1,3]。目前有关闽桦研究集中于分类学[1]、群落种间关系[3-4]及种子质量[5]等方面少数研究报道,导致其天然更新困难的原因是其自身生殖衰退还是生境限制,亦或是两者共同作用,原因尚未明晰。课题组前期对闽桦种子质量和萌发条件进行初步探索,发现黑色种子占比高,但无萌发能力[5],目前对于黑色种子形成的原因尚未探明。闽桦种群天然更新困难是黑色种子占比高导致还是某些环境因子对种子萌发限制导致,影响闽桦种子萌发的主要因素有哪些。
植物种子萌发受种子质量和环境因子共同作用。萌发率是衡量种子质量的重要指标,不同成熟期种子的萌发率有较大差异。通过对植物生殖物候规律的观测,了解物种的生殖对策,探究影响物种生殖的系统发育限制以及与非生物因子之间的关系[6],掌握植物种子成熟时间及生长适应机制[7-8]。金江群等[9]对极小种群崖柏生殖物候及胚胎发育过程进行研究,表明胚珠败育及雌配子体游离核时期至幼胚发育期间的败育是崖柏生殖障碍的主要原因。对濒危植物极小种群物种进行物候观测,一方面有助于了解植物的生物学和生态学特性,揭示其濒危机制;另一方面可以为种群复壮和迁地保护提供理论依据[9]。种子萌发过程需要经历一系列复杂的生理生化变化及形态结构改变,水分和温度是影响植物种子萌发主要环境因子,适宜的温度能确保这些变化顺利进行[10]。有关种子吸水性[11]、浸种时间[12]、温度[13]、土壤湿度[14]以及不同基质条件[15]对种子萌发率、萌发过程的生理指标的影响等已有大量研究[16]。本研究对闽桦生殖物候进行长期观测,比较不同成熟期种子萌发率,同时采用室内控制试验对不同成熟期闽桦种子萌发率的主要影响因子进行研究,探讨闽桦种子萌发生长的适宜条件,为闽桦种群扩繁和迁地保护提供理论依据和数据支撑。
1 研究区概况
福建省三明市罗卜岩自然保护区(117°34′19″E—117°35′44″E,26°25′57″N—26°27′25″N)位于三明市沙县西北部与明溪县交界处,坐落在武夷山东侧支脉。保护区地处中亚热带,气候温暖湿润,属中亚热带气候,年平均温度28.4℃,年平均湿度在80%以上。全区总面积约327.3 hm2,海拔250~683 m,为低山丘陵,山体呈东西走向,基岩以花岗岩为主,土壤为酸性红壤和红黄壤。区内地形复杂,富口溪蜿蜒横贯其间,溪岸多悬崖峭壁,形成独特峡谷景观。森林覆盖率达97.8%,属野生植物类型自然保护区,保护区以闽楠、闽桦等珍稀植物为主要保护对象,有维管束植物153 科517 属979 种,其中珍稀濒危植物23 种。闽桦与闽楠混交生长,分布于罗卜岩沟谷边和山坡上,为群落中的优势树种[17]。
2 研究方法
2.1 生殖物候观测
2017—2019 年连续3 年,随机选择5 株生长发育正常并已开花结实3年以上无病虫害发生的闽桦植株作为物候观测对象,所选植株皆处于开阔地带,便于观测,挂牌标识。借助单双筒望远镜、数码相机等工具,对闽桦的地上部分各器官的生长物候状况进行为期三年观测。选位于植株阳面的枝条作为观测对象,观察叶和雌雄花序等器官的形态变化,确定各器官的萌芽期、展叶期、开花期、果实发育期、叶片变色期和落叶期(见表1)。观测的时间,根据各器官不同生长期而定。(1)春季或物候变化较快的时期(如萌芽期和开花期),生长快,物候短暂,每日观测;(2)随着生长的进展,观察间隔时间延长,每3 d观测一次。生长后期7 d或更长时间观测一次。在开花结实季节(每年3—5月)每天观测、拍照、记录,其他月份每月固定时间观测并拍照。观测目标超过一半的器官到达目标形态,该时期为对应的物候期。

表1 闽桦生殖物候观测标准
2.2 采种与种子品质测定
为了探明闽桦种子的成熟时间及最佳采种时间,2018 年在雌花序成熟的时间段内,分早期、中期和晚期3 个时间(4 月25 日、30 日和5 月5 日)进行种穗采收,采收后的穗条均放在阴凉通风处晾晒1~2 d,待种子自然脱落,对3 个阶段种子的千粒重、含水率、黑色种子占比和发芽率等指标进行测定。
数相应的种子1000 粒各3 组,用万分之一的天平称取重量(g),测定千粒重;含水率参照李合生[18]的研究方法,每批种子随机取样0.2 g各3份,放入预先烘干的铝盒,置放到烘箱,80℃烘2~3 h,后调至105℃烘至恒重。种子含水率计算见公式(1)。
黑色种子占比采用每批种子随机抽取100粒种子各6组,黑色种子在每组中的比例,取平均值获得。同时将6 组种子放到培养皿(垫湿润滤纸)中,每天观测发芽率,连续3 d数据没有增加终止试验,发芽率计算见公式(2)[19]。
2.3 种子浸种处理方法
将不同时期采收的种子,随机称取3份,每份2 g,分别放入无纺布过滤袋中,采取常温无浸泡、常温浸泡5、10和20 h、恒温水浴锅30℃水温浸泡2、5、10和20 h共8 种处理。处理结束,将种子平铺于垫湿润滤纸培养皿中,每天记录发芽数,连续3天数据没有增加终止试验,不同浸泡处理发芽率计算见公式(3)。
2.4 种子储藏试验
将4月30日采收的种子,随机分成3袋,每袋随机取样2 g,采取常温、冷藏(0~4℃)和冷冻(0~-14℃)3种处理,分别于试验开始时、储藏1、2、3和4个月的时候,每处理3 个重复,进行发芽试验(同2.3),计算种子萌芽率,公式(4)。
2.5 基质对种子出苗率影响试验
将4 月30 日采收的种子,随机称取每份2 g,撒播于13 种不同苗圃基质条件中(见表2),每处理3 个重复,每天观测出苗数量。播种后第11天开始出苗并记录出苗率,记录后即拔除,连续3天数据没有增加终止试验。
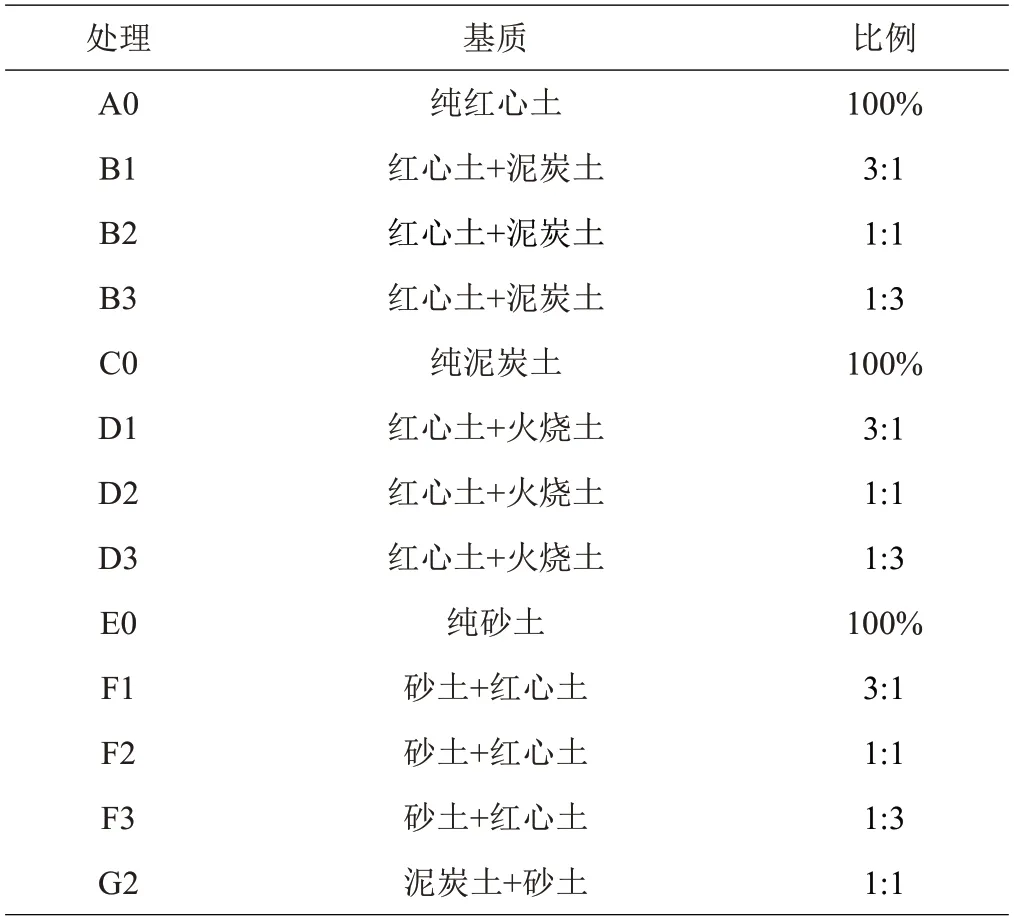
表2 13种不同基质
2.6 数据统计与分析
所有数据在Excel 2010 中进行整理,采用SPSS 22.0进行单因素方差分析,显著水平为α=0.05。
3 结果分析
3.1 闽桦生殖物候特征
闽桦主要器官物候变化时间在年际间有一定的差异(见表3)。闽桦是阔叶落叶树种,每年的3月中下旬新叶开始萌发并展开生长,直至秋季气温下降,开始落叶(10 月中旬—11 月中上旬),11 月下旬—12 月中旬,叶落尽(图1)。

图1 闽桦物候观测图

表3 2017—2019年闽桦生殖物候观测结果
闽桦是雌雄同株异熟落叶植物,雌花序生长期周期短,雄花序在前一年雌花序凋落后开始生长,生长时间较长。雄花序生长经历萌芽期、生长期、散粉期和凋落期4个时期,其中形成期和伸长期持续近10个月;散粉期和脱落期时间短,连接紧密,大部分在18~20 d内完成。
雌花序生长持续时间较短,和叶片生长时间部分重叠,从发芽到脱落,在56~58 d 内完成。经历萌芽期、生长期、成熟期和脱落期4 个时期,其中萌芽期和生长期持续时间大概32~36 d左右,授粉期11~13 d左右。3 月23 日—4 月10 日期间雌雄花絮完成授粉,雄花序脱落,雌花序进入成熟期,持续11~16 d左右。雌花序易被鸟取食或风折断,果实成熟后种子易脱落被风传播,坐果时间短,需及时采收,否则容易错过最佳采种时间。
3.2 采种时间对闽桦种子发芽率影响研究
不同采种时间闽桦种子的千粒重、含水率、颜色以及萌发率均有显著差异(表4)。随着成熟期的推进,黑色种子的占比逐渐增多,在早期采收的种子中,黑色种子仅占2.77%;在晚期采收的种子中,黑色颜色种子占比高达72.83%。成熟期的中期,闽桦种子的千粒重和含水率显著低于晚期种子,黑色种子占比33.3%,但发芽率显著高于早期和晚期。可见种子采收时间直接影响了种子的质量及发芽率,对育苗成效有极大的影响。
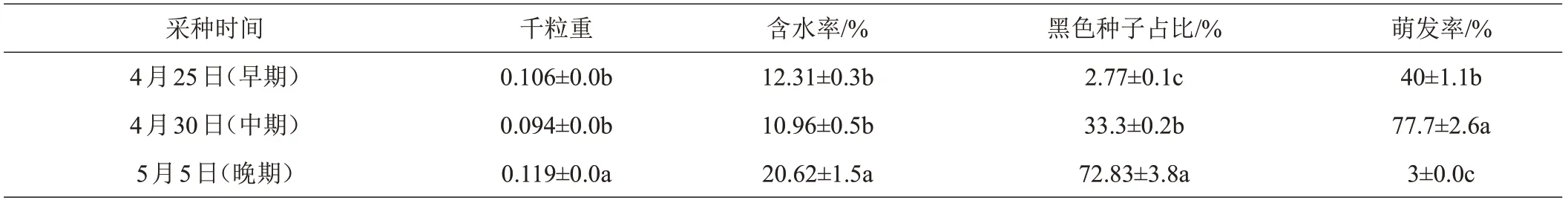
表4 不同颜色闽桦种子质量特征
3.3 不同浸种条件对闽桦种子发芽率影响研究
不同采种时间的种子采取常温和30℃水温下浸种时间发芽率比较研究,结果表明,成熟期中期采收的种子在各种条件下的发芽率均高于早期和晚期采收的种子(表5)。早期和中期采收的种子在常温下浸泡5 h的处理的发芽率显著高于其他处理,未浸泡种子的发芽率仅次于常温浸泡5 h的种子;晚期采收的种子在各种处理下的发芽率无显著差异,除常温浸泡5 h的外,发芽率均低于5%。可见种子采收时间对闽桦种子质量和发芽率有显著影响,闽桦种子在常温条件下浸泡5 h有利于提高其发芽率。
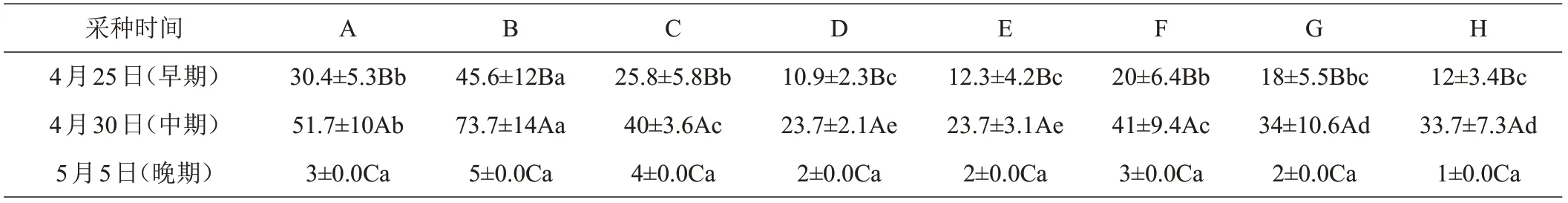
表5 不同采种时间不同浸泡条件下闽桦种子发芽率%
3.4 储藏条件及时间对闽桦种子萌发率影响研究
成熟期中期采收的闽桦种子在不同储藏条件下种子萌发时间集中于播种后第5~20 d,各处理种子萌发高峰期有差异(表6)。常温保存1 个月的种子萌发率接近0;冷藏2 个月发芽第10 天后萌发率显著高于其他处理;冷冻处理的种子随冷冻时间的延长种子萌发率降低。可见,闽桦种子不适宜常温下保存,宜随采随播或在冷藏或冷冻条件下进行短期储藏。
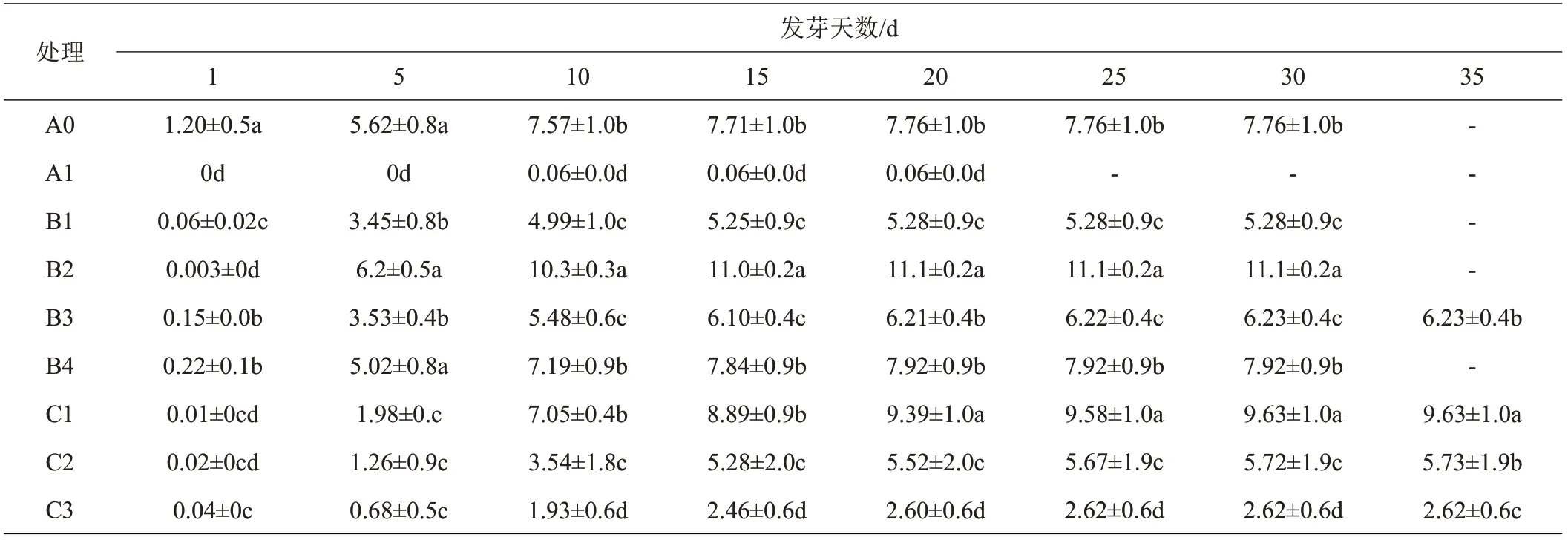
表6 不同储藏条件及时间闽桦种子萌发率 %
3.5 基质条件对闽桦出苗率影响研究
成熟期中期采收的种子在13 种不同基质和比例条件下出苗率随观测时间的延长呈波动变化趋势,出苗时间集中于播种后第11~25 天(见表7)。播种后第11天(观测第1天),A0处理纯红心土、B处理红心土+泥炭土各比例基质的出苗率显著高于其他基质条件的出苗率;播种后第15天,仅D2处理红心土+火烧土(1:1)基质条件下的萌发率显著低于其他基质条件,其他基质条件下出苗率无显著差异;播种后第25 天,各基质条件下的出苗率达峰值,F1 处理砂土+红心土(3:1)条件的总出苗率显著高于其他基质的出苗率。可见,闽桦大田播种苗床最适宜基质为砂土与红心土3:1混合。

表7 不同基质条件下闽桦种子出苗率 %
4 结论与讨论
4.1 闽桦生殖物候与种子质量
植物的生殖物候规律,是植物长期适应生长环境的进化策略。闽桦结实量大,与同科属植物亮叶桦相比黑色种子(败育种子)占比高[5],黑色种子的产生可能是闽桦种群衰退的内在因素之一。有性生殖过程是造成植物濒危至关重要的原因[20],传粉障碍、雌配子分化、胚发育过程异常等均可能导致败育[9,21-22]。闽桦种子成熟早期即有一定比例的黑色种子,可见在种子形成过程中,具备产生黑色种子的内在条件,建议对闽桦雌雄传粉过程及胚发育进行深入的研究,以期解决黑色种子产生的内在机制。气候等环境因子变化是导致种子败育的另一重要因素,有研究表明黄芩种子成熟期连续降雨导致瘪种率高,种子质量差[23];Owens等[24]对挪威云杉(Abies amabilis(Dougl.ex Loud.) Dougl.ex J.Forbes)的研究表明,其传粉期的霜冻是影响种子结实率的重要原因。闽桦黑色种子占比随着成熟期的推进逐渐增多,种子千粒重和含水率在成熟期晚期达到最大,可见除了内在因素外,局部小气候变化也许是影响黑色种子数量的关键因素。闽桦种子成熟季节为4月底雨季,黑色种子产生也许与降雨量或空气湿度有关,后期可将物候观测、气象数据结合闽桦种子产量质量进行连续观测研究,解决黑色种子产生的原因。
4.2 闽桦种子萌发主要影响因子
除种子质量外,温度、水分和光照等环境因子是影响种子萌发率的重要因素[25-26]。本研究结果表明,闽桦种子在常温条件下浸泡5 h显著提高萌发率。可见闽桦种子萌发对水分需求较高,闽桦种子千粒重仅0.1~0.2 g,种子本身的水分和养分含量极少,通过浸泡,种子吸收足够的水分,有利于提高萌发率。储藏条件影响了植物种子的萌发[27],不同储藏条件及时间发芽率试验结果表明冷藏2个月的闽桦种子萌发率显著高于常温和冷冻储藏种子萌发率,可见闽桦种子不宜常温下保存,宜在冷藏条件下进行短期储藏。闽桦种子于4月底成熟,春末夏初高温高湿天气居多,可能种子的呼吸作用等生理活动较强,自身养分和水分含量少,在常温条件下容易失活,需通过冷藏或冷冻来降低其生理活动,延长储藏时间。圃地基质条件是影响大田育苗成败的关键因素,本试验设置13种基质条件下闽桦出苗率影响研究,结果表明闽桦大田播种苗床最适宜基质为砂土与红心土3:1混合。据观察闽桦种子萌发前期胚根较短,砂土的比例较大,基质较疏松,便于其胚根向下延伸吸收养分和水分。除了基质条件外,苗期的水、肥、光照、虫害等管理也是影响出苗率的关键,有待进一步研究。
猜你喜欢
热带作物学报(2022年6期)2022-07-08
辽宁农业科学(2021年5期)2021-11-03
房地产导刊(2020年9期)2020-10-28
中国果树(2020年2期)2020-07-25
新农业(2018年6期)2018-04-18
西南农业学报(2017年5期)2017-06-23
河北林业科技(2016年5期)2016-11-08
西南农业学报(2016年5期)2016-05-17
浙江柑橘(2016年4期)2016-03-11
现代农业(2016年5期)2016-02-28
