
宁夏无芒稗对光周期的响应与适应
2023-12-11 10:03荆庆芳程云龙聂豪杰李林傲
草业科学 2023年11期
荆庆芳,朱 林,张 杨,程云龙,聂豪杰,李林傲
(1.西北土地退化与生态恢复省部共建国家重点实验室培育基地 / 西北退化生态系统恢复与重建教育部重点实验室 /宁夏大学生态环境学院, 宁夏 银川 750021;2.宁夏大学农学院, 宁夏 银川 750021)
光照是植物生命周期中最为重要的生态环境因素之一,为植物生长发育过程提供所需的能量[1]。植物在整个生育期中受到光周期的显著调控,使植物在生长发育过程中与特定环境条件相适应[2-3]。光周期不仅影响植物的光合作用,还作为信号因子直接影响植物整个生育期的发育和生理特性[4-5]。目前,国内外对光周期调控方面的研究发现,光周期不仅在植物从营养生长过渡到生殖生长阶中起着重要作用,还调节植物生长发育过程中的各个阶段[6-7],且在不同的生育阶段所需光照时间也不尽相同[8]。需长日照的生育阶段,短日照处理不利于植物生长,会降低光合色素的形成,在一定程度上抑制了叶片的光合作用,进而阻碍植物生长发育[9-10]。史莹华等[11]研究推测不适宜的光照时间可能作为一种胁迫因子影响苜蓿(Medicago sativa)生长。当植物遭受逆境胁迫时,抗氧化酶在抵御逆境胁迫产生过量的活性氧自由基(reactive oxyradical, ROS)中发挥重要的作用。目前,国内外关于不同光周期处理下植株抗氧化酶活性变化规律的研究鲜见报道,所以从抗氧化酶活性角度出发研究光周期影响稗属牧草生育期各指标改变的机理很有必要。
宁夏无芒稗(Echinochloa crusgalli)原为宁夏地区分布的野生无芒稗,后经人工驯化成为栽培品种,因其具备牧草种子产量和养分含量高等优良品质,现被作为重要的粮草兼用型作物[12]。目前,关于宁夏无芒稗的各项研究较少,大部分研究主要集中在耐盐碱性[13],而缺乏宁夏无芒稗对光周期响应的研究,且无法明确光周期对其生育期及生长生理特性的影响过程,这导致部分引种种植地区出现植株矮小,产量下降等情况,严重制约了区域土地资源合理利用及其畜牧业发展[14-15]。因此,本研究以宁夏无芒稗为研究对象,测定不同光周期处理下宁夏无芒稗拔节、抽穗开花、灌浆的生长指标,并分析不同光周期处理下各发育时期叶片的生理生化指标等,揭示不同光周期对宁夏无芒稗生长发育过程的调控作用,为我国耐盐碱优质牧草宁夏无芒稗的合理栽培及其异地引种提供重要依据,同时为实现盐碱土壤资源改良及可持续利用提供科学依据,能够更大程度实现盐碱土壤资源的合理利用,促进区域畜牧业的大力发展。
1 材料与方法
1.1 试验材料
试验材料为宁夏草原站提供的禾本科稗属牧草宁夏无芒稗。选取籽粒饱满、大小一致的种子用2%NaClO 溶液消毒20 min,蒸馏水冲洗4~5 次,最后用滤纸吸干表面水分后种植于营养土体积比为泥炭 土 ∶ 珍 珠 岩 ∶ 蛭 石 = 1 ∶ 1 ∶ 1 混 合 的500 g (干重)基质中。此外,每隔一周浇施一次霍格兰营养液(稀释50 倍),以保证全生育期植株正常生长。
1.2 试验设计
试验于2022 年3 月—6 月在宁夏大学人工气候室进行,设置光量子密度为180 μmol·(m2·s)-1,温度为(26 ± 1) ℃/(22 ± 1) ℃(昼/夜),湿 度 为35%。于2022 年3 月19 日在直径为23 cm、高度为13 cm 的花盆中播种,每盆播种20 粒宁夏无芒稗种子。待植株出苗一周后进行光周期处理,试验共设置5 个光周期处理,分别为8 (Ph8)、10 (Ph10)、12 (Ph12)、14(Ph14)、16 (Ph16) h·d-1,其中光照时间Ph8 为04:00—12:00,Ph10 为04:00—14:00,Ph12 为 04:00—16:00,Ph14 为04:00—18:00,Ph16 为04:00—20:00,其余时间均处于黑暗状态,每个处理20 株,重复3 次,共300 株。各光周期处理用遮光布分割成不同的试验区域,避免互相影响。不同处理之间除光周期不同外,其余管理措施均一致。此后,分别测定分析宁夏无芒稗在每个光周期处理下拔节期、抽穗期和灌浆期的生长指标及光合参数。之后,每株选取2~3 片功能叶片,用于后续生理生化指标的测定。
1.3 测定项目及方法
1.3.1生育期
拔节期:当不同光周期处理的盆栽中有1/2 的植株拔节时开始记录拔节期;抽穗期:当不同光周期处理的盆栽中有1/2 的植株抽穗时开始记录抽穗期;灌浆期:当不同光周期处理的盆栽中有1/2 的植株灌浆时开始记录灌浆期。
1.3.2 生长指标
不同生育时期测定植株的株高、叶长、茎粗、叶片数;在抽穗期测量旗叶的叶长、叶宽、叶面积,以及穗长、穗宽和穗重;并计算各生育期的地上生物量,每个光周期处理选取9 株(重复3 次)。
株高:由植株基部至主茎生长点的自然高度;茎粗:距土面2 cm 处的茎秆直径;叶长:由植株倒二叶的叶基部到叶尖的长度;叶片数:叶长大于1 cm的可见叶片数;采用叶面积仪(浙江托普云农科技股份有限公司,智能叶面积测量系统,YMJ-CH)测量旗叶的叶长、叶宽和叶面积;穗长:用直尺测量穗基部至穗顶部的长度;穗宽:用直尺测量植株主穗的最粗处为穗粗;穗重:用精度为0.01 g 电子天平称量其穗重;计算各生育期的地上生物量(拔节期:叶 +茎;抽穗期和灌浆期:叶 + 茎 + 穗)。
1.3.3 细胞膜透性
植株细胞膜透性采用相对电导率法,用DDS-307 型电导率仪测定[16]。
1.3.4 抗氧化酶活性
在宁夏无芒稗的不同生育期中,取不同光周期处理的新鲜叶片0.5 g,分2 次共加入7 mL 磷酸缓冲液,研磨成匀浆,用10 000 r·min-1冷冻离心20 min,取其上清液。其中,超氧化物歧化酶(Superoxide dismutase,SOD)活性采用NBT 光还原法[16];过氧化物酶(peroxidase,POD)活性采用愈创木酚法测定[16];过氧化氢酶(catalase,CAT)活性采用紫外吸收法测定[16]。
1.3.5 叶绿素含量
叶绿素a、叶绿素b 和类胡萝卜素含量采用95%的乙醇研磨法测定[16]。
1.3.6 光合气体交换参数
宁夏无芒稗的拔节、抽穗和灌浆期,每个处理随机选取9 株长势一致、叶片完好的植株,于09:30—11:30 采用 Li-6400 XT 便携式光合仪(Li-Cor Inc., Lincoln, NE, USA)测定。设定LED 红蓝光源,光强为1 000 μmol·(m2·s)-1,使用开放式气路,空气流速设定为500 μmol·s-1,选取从上至下数第3 个叶片进行净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)和蒸腾速率(Tr)的测定,并利用以下公式计算瞬时水分利用效率(WUE ):
1.4 数据分析
采用Excel 2010 整理数据;所有统计分析均使用SPSS 23.0 进行,用平均值和标准误表示测定结果,采用单因素方差分析(one-way ANOVA)和双因素方差分析(two-way ANOVA),最后采用Origin 2021 制图。
2 结果与分析
2.1 不同光周期处理对宁夏无芒稗生育进程的影响
不同光周期处理下,宁夏无芒稗完成生育期所需的时间不同(图1)。在光照时间10 h 处理下,宁夏无芒稗出苗到灌浆所需的天数最短,仅需60 d。在长日照(16 h)处理下,其所需的天数明显延长,需要89 d。说明较长的光照时间下,宁夏无芒稗的生长缓慢。
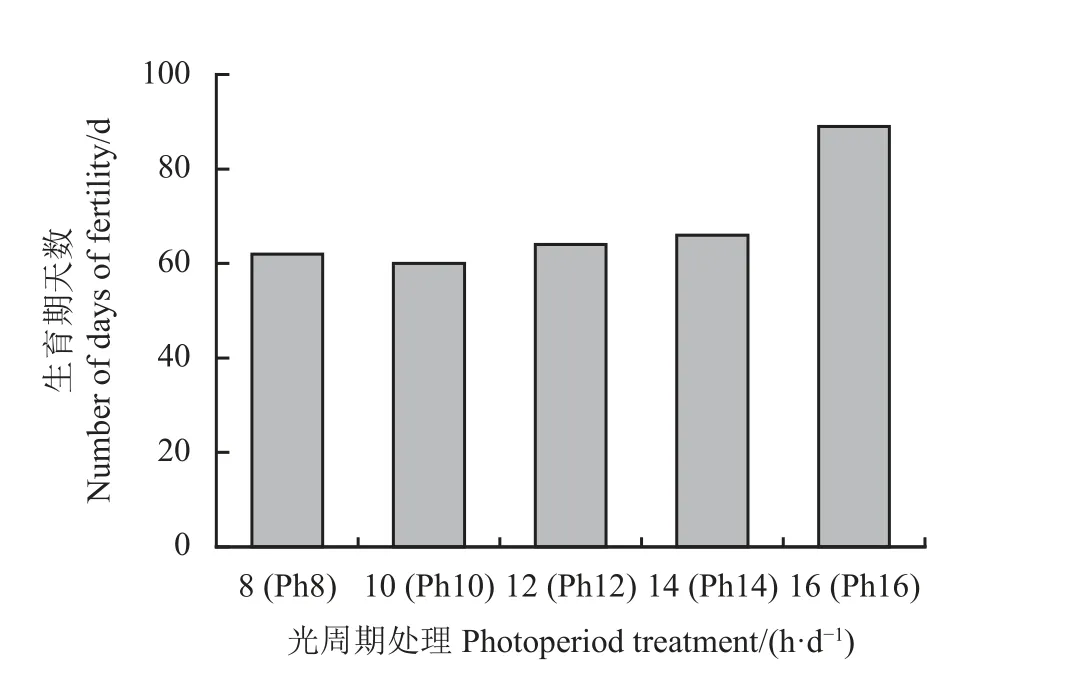
图1 光周期处理下宁夏无芒稗灌浆情况Figure 1 Grouting of Echinochloa crusgalli under photoperiod treatments
2.2 光周期对宁夏无芒稗不同生育时期生长指标的影响
不同光周期处理对宁夏无芒稗各生育期的生长状况均具有重要影响(表1)。其中,在Ph16 处理下宁夏无芒稗各生育期的株高、叶长和地上生物量均显著高于其他处理(P< 0.05),但叶片卷曲严重。随着光照时间的延长,在宁夏无芒稗的不同生育期中茎粗和叶片数均呈“先升后降”的变化趋势,在Ph12 处理下达到最大。随着生育期的推进,宁夏无芒稗的生长指标均表现为灌浆期 > 抽穗期 > 拔节期。且生育时期和光周期对各生长指标具有极显著影响(P< 0.01),生育时期和光周期交互作用对茎粗和地上生物量的影响极显著(P< 0.01),对株高、叶长和叶片数影响不显著(P> 0.05)。
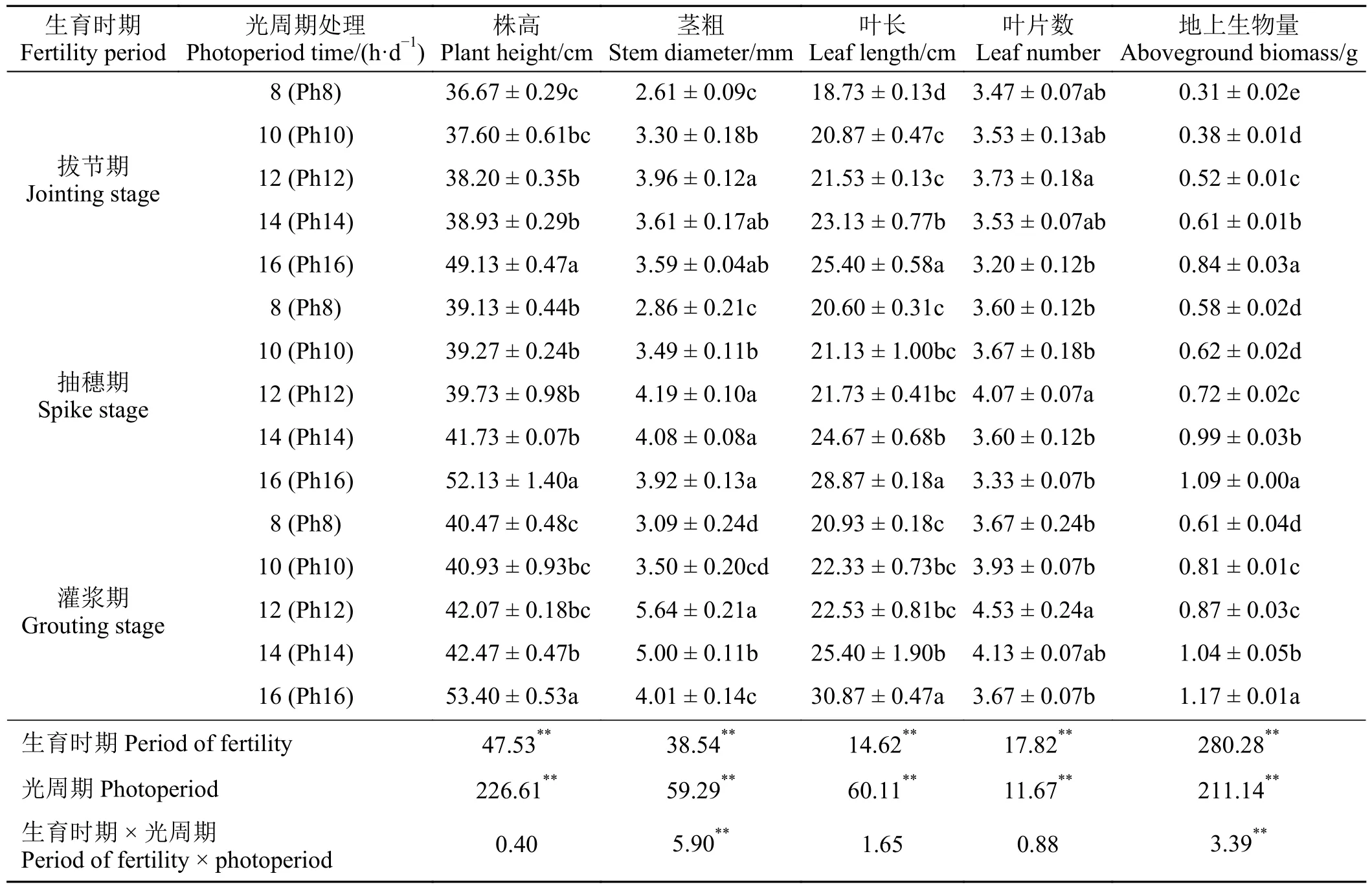
表1 不同生育时期和光周期对宁夏无芒稗生长指标的影响Table 1 Effects of different fertility periods and photoperiods on the growth index of Echinochloa crusgalli
不同光周期处理对宁夏无芒稗抽穗期的旗叶性状和穗部特征均产生重要影响(表2)。在Ph12 处理下,宁夏无芒稗的旗叶长和旗叶面积显著高于其他处理(P< 0.05),分别提高了31.91%、26.03%、39.86%、76.67%和38.83%、35.51%、56.89%、130.59%。而旗叶宽在Ph16 处理下显著低于Ph10 和Ph12 (P<0.05),分别下降17.40%和23.87%;较其他光周期处理无显著差异(P> 0.05)。与旗叶面积结果类似,宁夏无芒稗的穗部特征也均呈“倒V”型变化趋势,均在Ph12 处理下达到最大。穗重在Ph12 处理下显著高于其他处理(P< 0.05),分别上升67.50%、48.89%、34.00%和45.65%,而穗长和穗宽在不同光周期处理间无显著差异(P> 0.05)。这可能是较短或较长日照下,宁夏无芒稗的旗叶面积较小,不利于其进行光合作用,进而导致光合产物不能满足灌浆需求,最终体现在穗重下降。
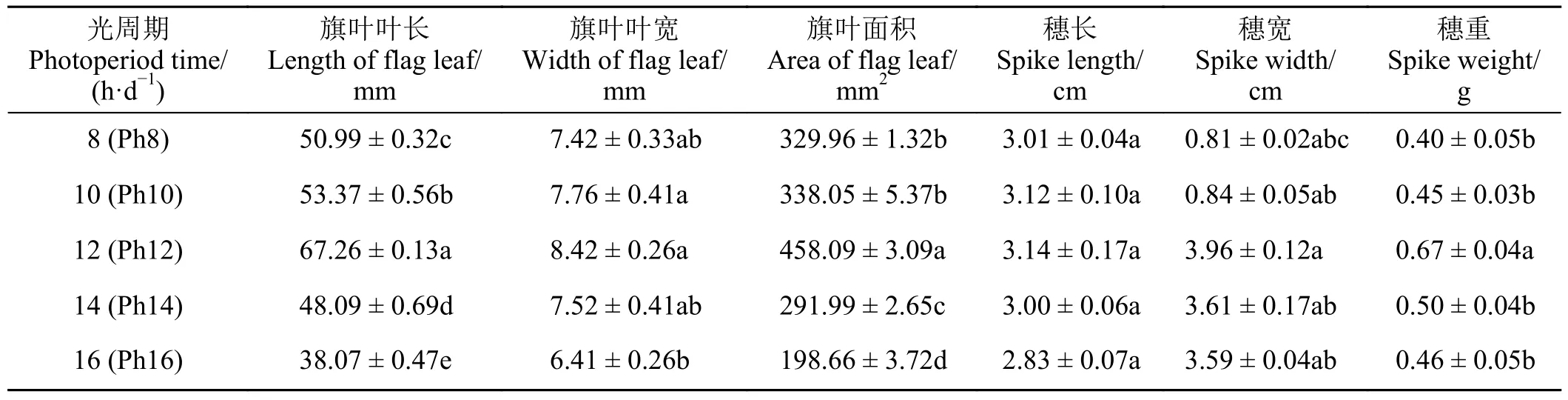
表2 光周期处理下宁夏无芒稗生长指标的比较Table 2 Comparison of growth indices of Echinochloa crusgalli under photoperiodic treatment
2.3 光周期对宁夏无芒稗不同生育时期生理生化指标的影响
2.3.1 细胞膜透性
细胞膜透性的大小可以直接反映光照时间对植株细胞膜透性的改变及细胞膜损坏的情况。不同光周期处理对宁夏无芒稗细胞膜透性产生重要的影响(图2)。随着生育进程的推进,宁夏无芒稗的细胞膜透性呈不断增加的变化趋势,表现为灌浆期 > 抽穗期 > 拔节期。且随着光照时间的延长,宁夏无芒稗各生育期的细胞膜透性呈“先降后升”的变化过程,均在Ph12 处理下降到最小值,分别为42.02%、45.28%和44.29%。且在Ph12 处理下的细胞膜透性显著低于Ph8 和Ph16 处理(P< 0.05)。

图2 光周期处理下宁夏无芒稗细胞膜透性的比较Figure 2 Comparison of cell membrane permeability of Echinochloa crusgalli under photoperiodic treatment
2.3.2 抗氧化酶活性
SOD 作为植物体内抗氧化酶体系的核心酶,能够通过催化对细胞有害活性氧和自由基发生歧化反应使活性氧生成H2O 和O2,在活性氧清除系统中起到关键作用。如图3 所示,随着光照时间的延长,宁夏无芒稗各生育时期的SOD 活性均呈“先降后升”的变化趋势。拔节期在Ph16 处理下的SOD 活性显著高于Ph8、Ph10 和Ph12 处理(P< 0.05);抽穗期在Ph16 处理下显著高于其他处理(P< 0.05);灌浆期在Ph8 处理下显著高于Ph12 和Ph14 处理(P<0.05),与其他处理无显著差异。说明较短和较长的日照长度激发SOD 的活性,从而使得植株叶片的SOD 活性升高。
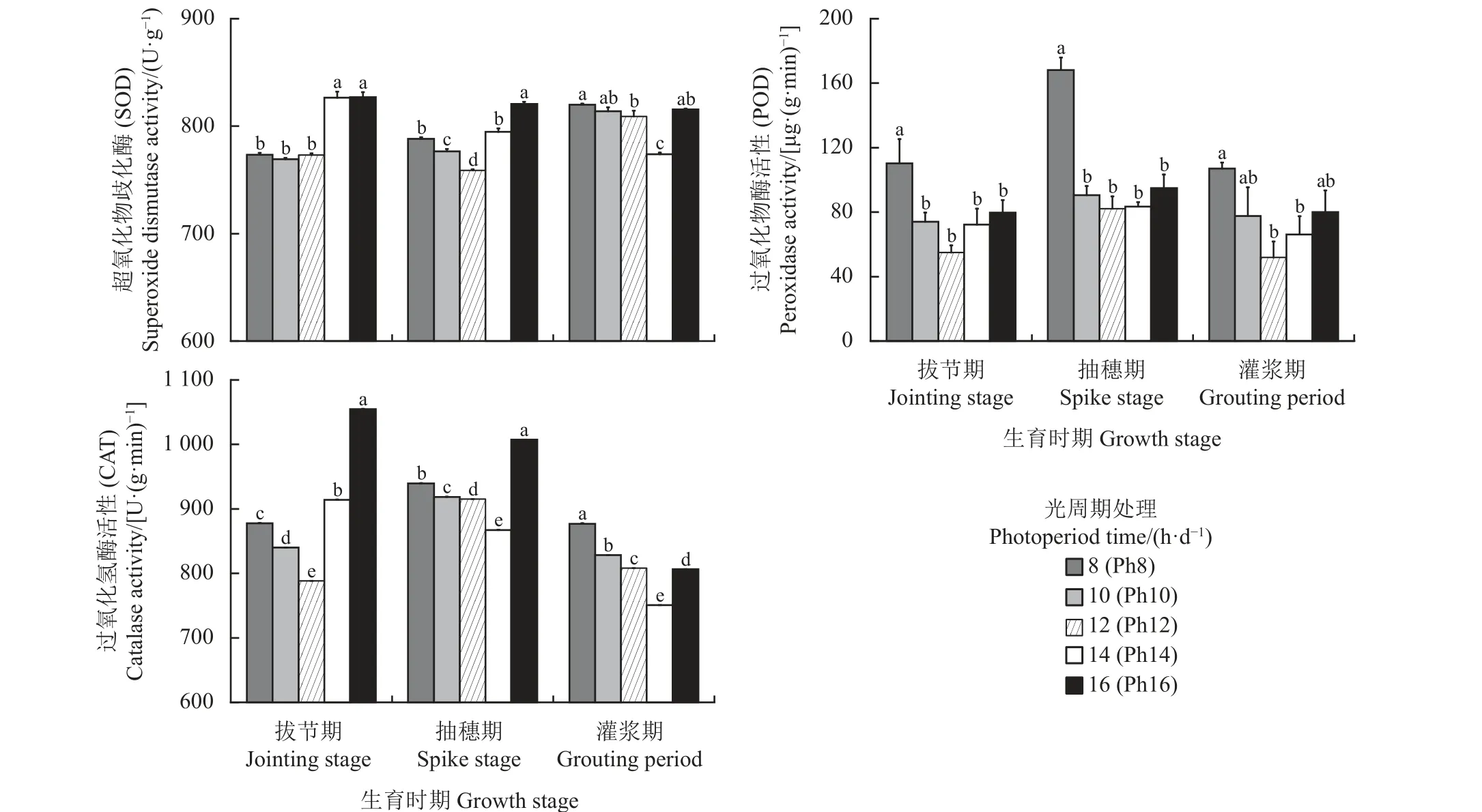
图3 光周期处理下宁夏无芒稗抗氧化保护酶活性的比较Figure 3 Comparison of antioxidant protective enzyme activities of Echinochloa crusgalli under photoperiodic treatment
POD 作为植物体内的保护酶之一,能够清除细胞内的自由基,其活性的高低可反映某时期植物的代谢及抗逆性。如图3 所示,随着生育时期的推进,宁夏无芒稗的POD 活性呈“先升后降”的变化趋势。而随着光照时间的延长,宁夏无芒稗各生育时期的POD 活性均呈“V”型变化趋势,均表现为Ph8 处理下POD 活性最高,Ph16 次之。说明光胁迫可能会刺激宁夏无芒稗叶片的POD 活性升高,从而减轻活性氧对植株细胞的损伤。
CAT 能够清除代谢中产生的H2O2,避免H2O2积累对膜系统造成伤害。随着光照时长的增加,宁夏无芒稗各生育期的CAT 活性呈“先降后升”的趋势(图3)。拔节期在Ph12 处理下CAT 活性显著低于其他处理(P< 0.05),分别下降10.19%、6.17%、13.77%和25.27%;抽穗期和灌浆期时,在Ph14 处理下显著低于其他处理(P< 0.05),分别下降7.71%、5.59%、5.25%、13.91%和14.37%、9.34%、7.09%、6.92%。说明较短或较长的光照时间可能会刺激宁夏无芒稗叶片的CAT 活性升高,以此清除细胞内多余的活性氧,进而保护植株细胞免受伤害(图3)。
2.3.3 叶绿素含量
叶绿素作为植物进行光合作用的重要色素,其含量高低直接影响植物的光合作用,还可作为衡量叶片衰老的重要生理指标,在一定程度上也能够反映植株的代谢能力和抗逆性[17]。随生育时期的不断推进,叶绿素b (Cb)含量在P14 和P16 处理下呈先升后降的趋势,在其他处理下均呈下降趋势(图4),说明作物随着生育期的不断推进,植株叶片的光合色素含量整体将逐渐下降。因此,宁夏无芒稗在各生育时期的叶绿素含量表现为拔节期 >抽穗期 > 灌浆期。随着光照时间的增加,宁夏无芒稗各生育期的叶绿素a、叶绿素b 和类胡萝卜素含量呈“先升后降”的变化趋势,均在Ph12 处理下达到最大。说明光照时间较短或较长可能导致植株叶片的叶绿素合成不足或抑制其合成。
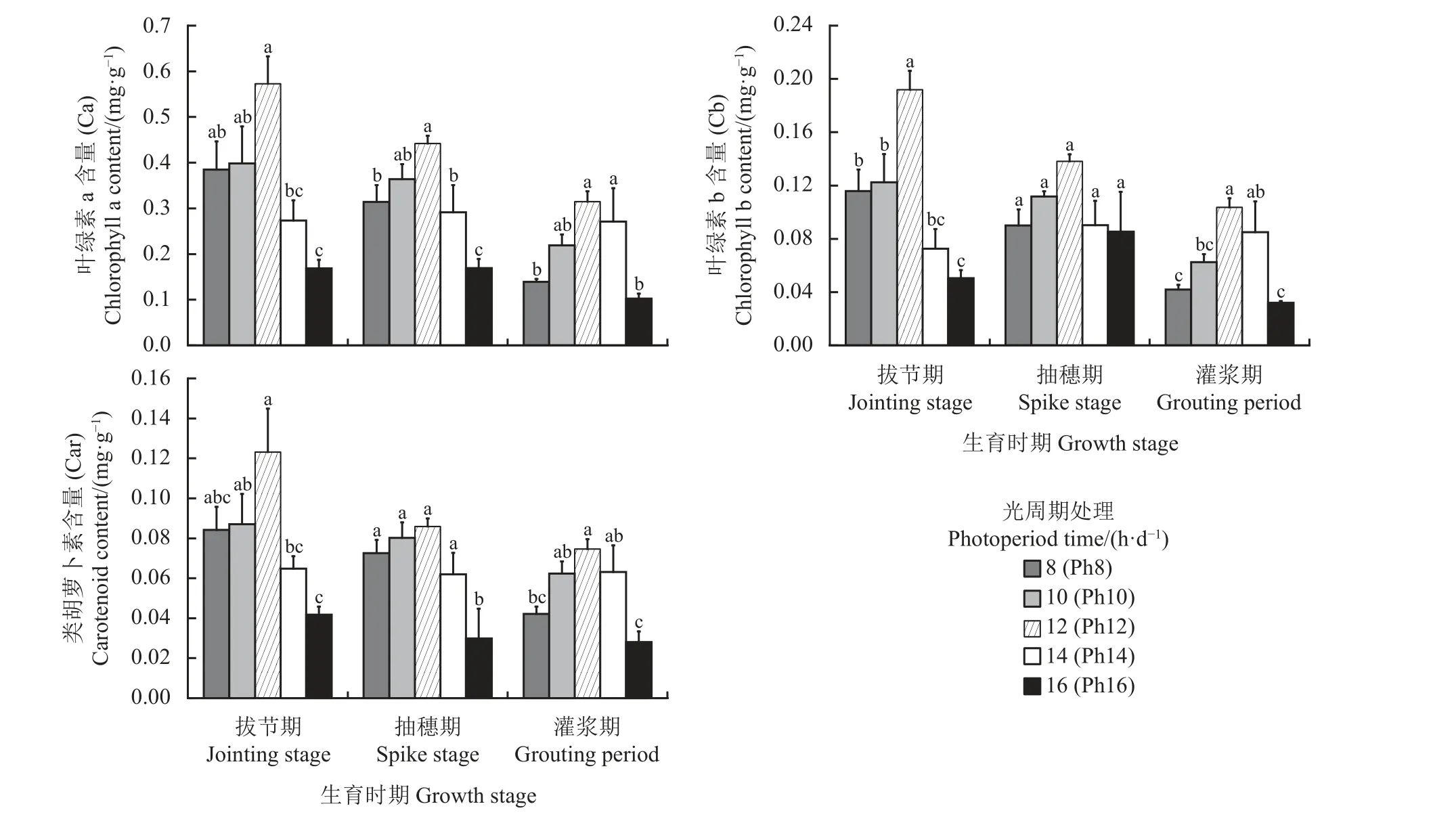
图4 光周期处理下宁夏无芒稗叶绿素含量的比较Figure 4 Comparison of chlorophyll content of Echinochloa crusgalli under photoperiodic treatment
2.3.4 光合参数
光合作用是植物生长发育的基础,也是植物对环境变化最为敏感的生理反应之一。不同光周期处理下,宁夏无芒稗在各生育期的光合参数发生变化(图5)。随着光照时间的延长,宁夏无芒稗各生育期的净光合速率整体呈“先升后降”的变化趋势,均在Ph16 处理下显著低于其他处理(P< 0.05)(图5)。可能是长日照条件下,宁夏无芒稗的叶片严重卷曲,不利于植物进行光合作用,进而导致其净光合速率下降。且气孔导度和瞬时水分利用效率也呈“倒V”型变化过程,蒸腾速率也出现类似规律,在Ph16 处理下宁夏无芒稗的蒸腾速率显著低于Ph12 处理(P< 0.05),分别降低了63.27%、74.85%和57.45%。
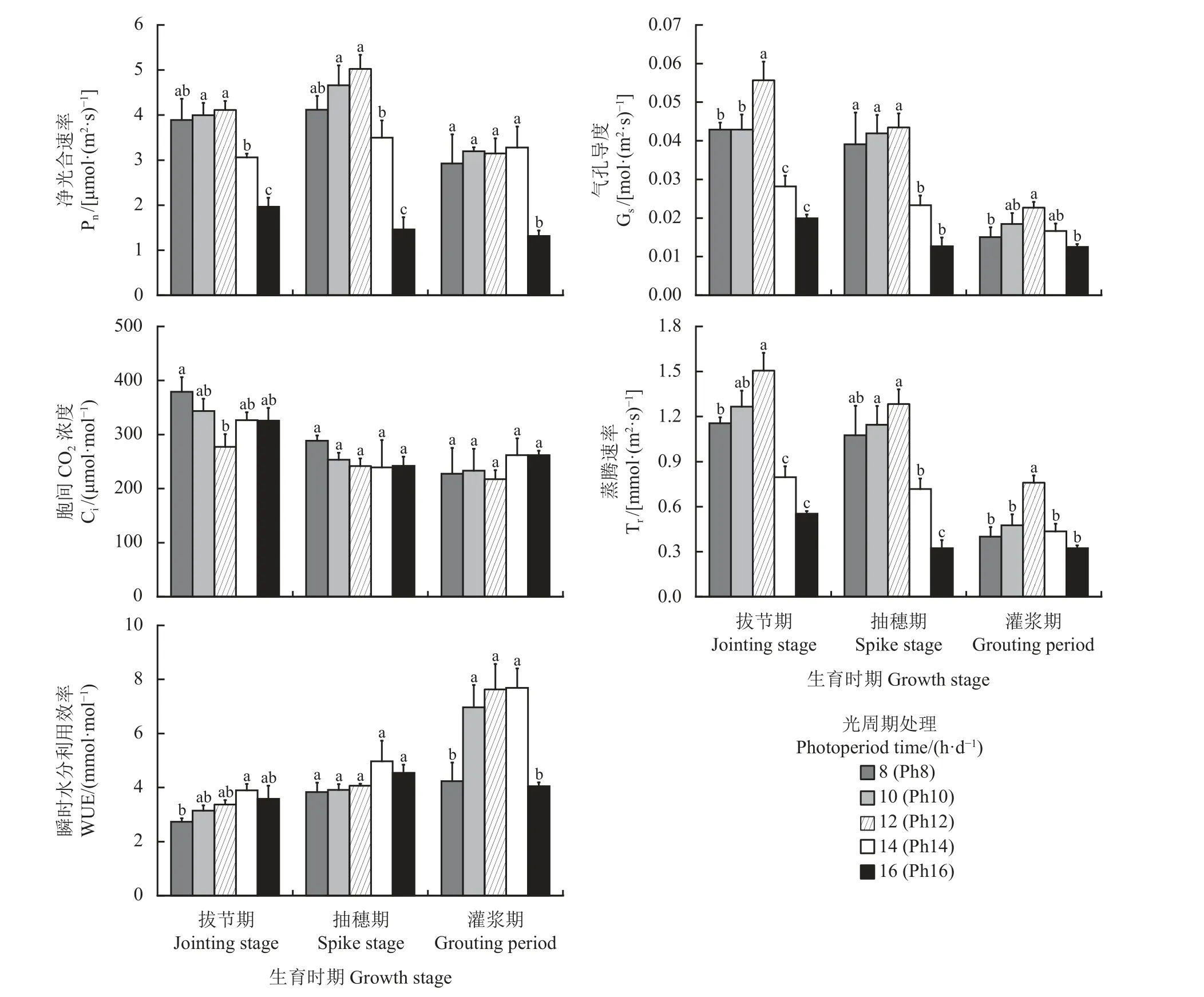
图5 光周期处理下宁夏无芒稗光合参数的比较Figure 5 Comparison of photosynthetic parameter of Echinochloa crusgalli under photoperiodic treatment
2.4 光周期处理下宁夏无芒稗不同指标间的相关性
如图6 所示,穗重(SW)和旗叶面积(LA)、Car呈显著正相关关系(P< 0.05),与Gs、Tr、Ca、Cb 呈极显著正相关关系(P< 0.01),而与Pn呈正相关关系;反之,SW和3 种抗氧化酶呈负相关关系。LA 与Pn、Gs、Tr、Ca、Cb、Car 之间呈极显著正相关关系(P< 0.01)。Pn与Ca、Cb、Car 之间呈极显著正相关关系(P< 0.01),而与Ci、SOD、POD、CAT 呈负相关关系。SOD 与CAT 呈极显著正相关关系(P<0.01),而与POD 呈正相关关系。通过对各指标的相关性分析发现,穗重与旗叶面积、光合、叶绿素含量等具有明显的相关性,说明叶绿素含量、光合等参数能够反映旗叶面积的大小,而旗叶面积又能够决定宁夏无芒稗种子产量的高低。
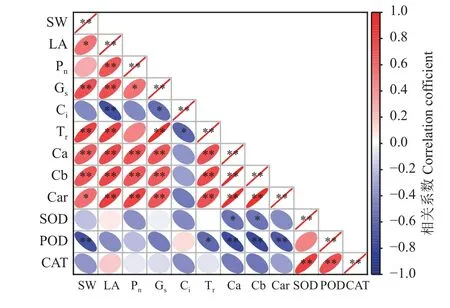
图6 光周期处理下宁夏无芒稗各指标的相关性Figure 6 Correlation of the indices of Echinochloa crusgalli under photoperiodic treatment
3 讨论与结论
光周期通过调控植物的生长发育进程,进而影响植物的产量和品质。本研究表明,随着光照时间的延长,宁夏无芒稗的整个生育期长短发生显著的变化。在长日照(16 h·d-1)条件下,宁夏无芒稗的生育期显著延长,生长缓慢。这与谢丽莉等[18]对玉米(Zea mays)和狗尾草(Setaria viridis)的光周期敏感性研究中结论相一致。这一发现表明过长的光照时间将延长宁夏无芒稗的生育期,可能会限制种子的发育或使植株不能避免恶劣的自然环境,并不利于牧草的生长发育。另外,光周期通过调控牧草的生长指标,进而影响牧草的产量[19]。本研究发现,宁夏无芒稗各生育期的株高均随着光照时间的延长呈不断上升的趋势,而茎粗和叶片数均呈先升后降的变化趋势,这与王玉卓等[20]的研究结果基本一致,说明适当延长光照时间有利于植物的生长。此外,还有研究表明,旗叶性状是抽穗开花至成熟期影响籽粒产量的重要因素[21]。本研究发现,随着光照时间延长,旗叶各性状均呈先升后降的趋势,与之相同,籽粒产量也呈现相似的变化规律,造成该现象的原因可能是光照时间太短或太长使得旗叶面积小且卷曲严重,不利于旗叶进行的光合,从而造成籽粒产量较低。但是发现随光照时间的延长,宁夏无芒稗各生育期的地上生物量则呈上升趋势,这与前人研究结果一致[22],该现象的可能原因是随着光照时间的延长,宁夏无芒稗的营养生长期显著延长,有利于植株干物质的积累。
当外界环境变化时,植物能够通过调节抗氧化酶活性及渗透调节物质含量以此来适应环境的变化[23]。已有研究表明,在光胁迫下植物体内的抗氧化酶能够减轻光胁迫对植株的伤害,其中SOD 主要作用是清除超氧物阴离子自由基,CAT 和POD的作用是清除细胞内过多的过氧化物,是活性氧清除酶系统的重要保护酶,能够有效清除自由基氧化损伤[24]。本研究发现,随着光照时间的延长,宁夏无芒稗各生育期的SOD、CAT 和POD 活性均呈先降后升的趋势,造成该现象的原因可能是中等日照长度下,宁夏无芒稗叶片的超氧阴离子自由基的产生和清除处于动态平衡,所以不同程度地降低了3 种酶的活性;相反,光照时间太短或太长会刺激宁夏无芒稗叶片3 种酶活性的升高,以此清除植株细胞内多余的活性氧,减轻对植株叶片的伤害,以确保植株光合作用持续正常的进行。这与赵宝平等[25]关于光周期对燕麦(Avena nuda)影响的研究结果相似。
另外,叶绿素不仅在一定程度上反映植物的光合能力,而且还能反映植物的生长情况[26]。光照时间的变化可影响植物体内叶绿素的含量。本研究表明,随着生育期的推进,叶绿素a 和类胡萝卜素含量均呈下降趋势,可能是随着生育期的推进,叶片逐渐黄化衰老,进而导致宁夏无芒稗叶片的叶绿素含量逐渐降低[27]。此外,随着光照时间的延长,宁夏无芒稗各生育期的光合色素含量均呈先升后降的变化趋势,在Ph12 处理下其叶绿素含量最高。由此说明适当延长光照时间有利于宁夏无芒稗叶片光合色素的积累,使叶片增绿。造成该现象的原因,可能是短日照限制植株的光合碳同化和光合酶等活力,进而抑制了叶绿素的合成,或延续的过长日照下,植株的光合产物达到饱和后将产生负反馈作用,进而抑制叶片中叶绿素的合成,使得叶片中叶绿素含量降低[28-29]。这与韦朝妹等[30]关于补光时长对多肉‘艳日辉’(Aeonium decorumf.variegata)形态及生理影响的研究结论一致。
光合作用是植物通过叶片中叶绿素将光能进行吸收传递最终转变成化学能的过程,植物体内叶绿素含量的多少直接影响植物的光合速率等[31]。此外,光周期对植物光合参数的影响受生育期等影响较大[3]。已有研究发现,造成植物光合速率降低有两种限制因素:一是气孔关闭导致的气孔限制因素;二是光合色素的降解及合成受阻等造成的非气孔限制因素[32]。本研究发现,随着生育时期的推进,宁夏无芒稗的光合参数呈下降趋势。这可能是随着生育期推进,宁夏无芒稗叶片的叶绿素含量下降,从而使得光合速率减弱。同时,随着光照时间的延长,宁夏无芒稗各生育期的Pn、Gs、Tr和WUE整体呈“倒V”型变化过程,Ci则呈先降后升的变化趋势。因此,宁夏无芒稗的Pn下降可能是受非气孔限制因素的影响。李孟洋等[33]关于缩短光周期对茅苍术(Atractylodes lancea)影响的研究中也发现了类似的现象。该现象的潜在机制可能是不利的光照条件使植株体内产生较多的活性氧,进而对植株的光合作用结构和功能造成损伤;也可能是短日照使得植株叶绿素合成不足,而长日照抑制叶绿素的合成,导致植株的Pn下降。这一发现说明太长或太短的日照时间均不利于宁夏无芒稗的光合作用,从而影响其正常的生长发育,该现象的机制可能是,适当延长光照时间,使植株进行光合作用的时间增加,合成积累更多的有机物以支持宁夏无芒稗的生长发育,反之较短的光照时间会降低有机物的合成,不利于植株的生长发育。
综上所述,本研究通过分析不同光周期对宁夏无芒稗拔节期、抽穗期和灌浆期3 个生育时期的生长和生理生化指标的影响。结果表明中等日照长度(12 h·d-1)条件下是宁夏无芒稗生长发育的适宜光照条件;而在长日照(16 h·d-1)下,宁夏无芒稗的生育期显著延长,生长缓慢。其潜在的调控机制是:1)适当延长光照时间有利于叶片中光合色素的积累,从而增强了宁夏无芒稗叶片对光能的传递和利用效率,提高光合效率,合成积累更多的光合产物。2)适宜延长光照时间下,植株通过抗氧化酶系统及时清除多余的活性氧,以维持体内自由基代谢的动态平衡,进而保护植株膜系统的稳定性,有利于植株的光合作用。值得注意的是,本研究设置在人工气候室中进行,削弱了其他环境因子对植物生长发育的影响,因而结果具有一定的局限性。因此,在未来的研究中需要通过田间试验进一步研究和验证,明确其适宜的光照长度,为其合理引种提高科学依据,以避免不适宜地区引种种植后其产量及品质下降,严重制约区域土地资源合理利用,从而造成土地和牧草资源浪费等问题。
猜你喜欢
遗传(2023年9期)2023-09-25
农业科技通讯(2023年1期)2023-02-12
中国机械工程(2022年8期)2022-05-09
中国土壤与肥料(2021年5期)2021-12-02
中国机械工程(2021年8期)2021-05-07
音乐教育与创作(2019年8期)2019-05-16
少儿科学周刊·少年版(2017年7期)2017-09-29
少儿科学周刊·儿童版(2017年7期)2017-09-29
西南农业学报(2016年5期)2016-05-17
西藏科技(2015年5期)2015-09-26
