
Micromammal fossils from the basal part of the Jiaozigou Formation in Yagou area,Linxia Basin,Gansu Province
2023-12-08 02:42WANGBanYueQIUZhanXiang
古脊椎动物学报(中英文) 2023年4期
WANG Ban-Yue QIU Zhan-Xiang
(Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences Beijing 100044 wangbanyue@ivpp.ac.cn)
Abstract Renewed fieldwork in 2003 produced a rich micromammal assemblage from the basal part of the Jiaozigou Formation in the Yagou area of Linxia Basin.17 genera distributed among 13 families of three orders were recovered.The micromammal fauna is a typical Oligocene assemblage for Central and Eastern Asia.14 genera (~82% of the fauna) are common in the Oligocene of Asia.Of them four genera and four species of Eucricetodon are restricted to the Oligocene,one genus (Bagacricetodon) is restricted to Late Oligocene and Glis and Eomyodon made their first appearances in the Late Oligocene.Based on this micromammal composition,the basal part of the Jiaozigou Formation in the Yagou area could be mainly of Late Oligocene in age,which is in accordance with the conclusion based on large mammal fossils.In comparison with the other Late Oligocene micromammal faunas in Central and East Asia,the Yagou Fauna is slightly older than the Ulan III biozone of Nei Mongol and biozone C of Mongolia,because it has two Eocene genera and lacks more advanced genera.This is roughly in accordance with the recent palaeomagnetic interpretation for the Maogou section,where the lower boundary of the Jiaozigou Formation was correlated with Chron C10r (~29 Ma).The presence of large number of xerophilous zapodines,ctenodactylids,cricetids and lagomorphs combined with fossorial Tsaganomys and the lithology of the fossil-bearing deposits,composed of gypsiferous reddish brown mudstone,tend to show a semiarid woodland-shrubland habitat during the Late Oligocene in Yagou area.In Late Oligocene more frequent faunal interchange might have occurred between Asia and Europe (4 genera commonly shared) rather than between Asia and America (only 1 genus shared),partly because of the disappearance of the Turgai Strait.
Key words Yagou,Linxia,Gansu;Late Oligocene;Jiaozigou Formation;micromammal fossils
1 Introduction
The Jiaozigou Formation was named by Qiu et al.in 1990.At first considered as Early Miocene in age (Qiu et al.,1990),it later was transferred to Late Oligocene (Deng et al.,2004,2013;Qiu et al.,2004a,b),based on revised study of the large mammal fossils.However,the transfer of the fossiliferous layers of the Jiaozigou Formation from Early Miocene to Late Oligocene based largely on the removal of proboscidean fossils remained unsatisfactorily convincing and should be further confirmed by other means,especially from the field of the micromammals.Fortunately,during a field trip in 2003,Prof.Zhang Zhaoqun found one tooth ofDesmatolagusfrom the lower part of the Jiaozigou Formation in Yagou area.Zhang’s site was then chosen as a place to apply wet-sieving to collect micromammal fossils.Ultimately,three micromammal localities were registered: two in Nanlong Town,Linxia City (LX 200301 and LX 200303) and one in Dongyuan Town,Dongxiang Autonomous County (LX 200302)(see Fig.1;Table 1).Please note: the large mammal locality LX 200213 (near LX 200302) had been incorrectly labeled LX 200214 in Qiu et al.(2004b),but later corrected as LX 200213 by Deng et al.(2013).All small and large mammal fossils come from the same brown and yellow sandstone layers with purple mudstone in the basal part of the Jiaozigou Formation in the Yagou area (Fig.2A).

Table 1 Localities yielding mammal fossils in lower part of Jiaozigou Formation in Yagou,Linxia Basin
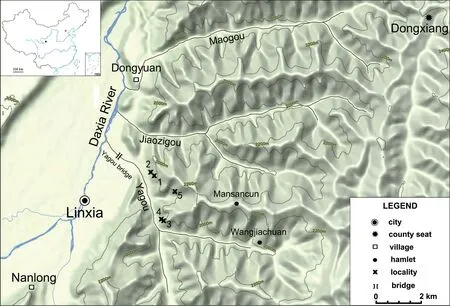
Fig.1 Map showing the localities yielding mammal fossils from the Jiaozigou Formation near Yagou in the Linxia Basin

Fig.2 Cenozoic strata in Yagou
All 198 micromammal specimens from the basal part of the Jiaozigou Formation are isolated teeth,representing 17 genera and 13 families (Orders Eulipotyphla,Lagomorpha and Rodentia).These fossils show that the basal part of the Jiaozigou Formation in the Yagou area is earliest Late Oligocene or possibly latest Early Oligocene in age.
As to the terminology of cheek tooth elements and measurements,we follow Qiu and Tong (2015) for Erinaceidae,Li (1965) and Qiu (1996) for Lagomorpha,Wang and Qiu (2019)for Aplodontidae,Lu et al.(2016) for Mylagaulidae,Daams (1999) and Freudenthal (2004) for Gliridae,Wu (2019b) for Eomyidae,Qiu and Wang (2019) for Zapodinae,Wang (1997b) for Ctenodactylidae,and Gomes Rodrigues et al.(2012) and Wang et al.(2020) for Cricetidae.
The measurements (in mm): length (L) × width (W).
2 Systematic paleontology
Order Eulipotyphla Waddell Okada &Hasegawa,1999
Suborder Erinaceomorpha Gregory,1910
Family Erinaceidae Fischer von Waldheim,1814
Subfamily Erinaceinae Fischer von Waldheim,1814
Amphechinus Aymard,1850
Amphechinus sp.
(Fig.3A-C)
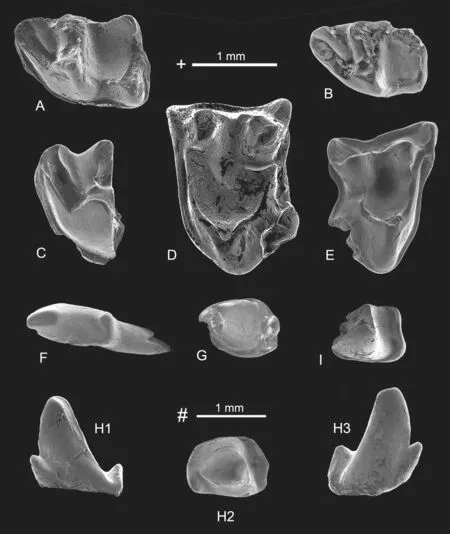
Fig.3 The lipotyphla from Yagou area
SpecimensLX 200301: left m1,trigonid of left m2,and talonid of left m1/2 (IVPP V26968.1-3);LX 200303: left m1 (V26969).
Measurementsm1 (V26969): 1.50 × 1.00,trigonid: 0.95 × 1.00,talonid: 0.60 × 1.00;m1 (V26968.1): 1.35 × 0.90,trigonid: 0.85+× -,talonid: 0.50 × -;m2 (V26968.2): trigonid:0.80 × 1.00;m1/2 (V26968.3): talonid: 0.75+× 1.00.
Description and comparisonOn the m1 (V26969 and V26968.1) the trigonid is low and narrow,longer than talonid;the paralophid extends anterolingually and has a distinct paraconid;the entoconid is higher than the hypoconid with a developed entoconid crest extending to the posterior side of the metaconid;the oblique crista is low and buccally located,extending to the posterior side of protoconid;the sinusid is shallow;the hypoconulid is located at the middle part of the hypolophid;the sloping ridge is distinct (V26969) or indistinct(V26968.1).These features suggest a small species ofAmphechinus.
The trigonid of m2 (V26968.2) is shorter than the trigonid of m1 and the paralophid is a low and circular as in m2 of otherAmphechinus.The talonid of V26968.3 is similar to that of V26969 in its longer entoconid crest extending to the posterior side of the metaconid and sloping ridge extending from the middle part of the hypolophid.The trigonid of m2 (V26968.2)and talonid of m1/2 (V26968.3) are close to those of V26969 and V26968.1 in size,so they may represent the same species.
Although smaller than knownAmphechinusin size,these teeth are too fragmentary to show distinctive features.They are identified asAmphechinussp.temporarily.The geological age ofAmphechinusis Oligocene and Miocene (Qiu and Tong,2015).
Family Changlelestidae Tong &Wang,1993
Subfamily Tupaiodontinae Butler,1988
Anatolechinos Wang,2008
cf.Anatolechinos sp.
(Fig.3D)
SpecimenOne left M2 (IVPP V26970) from LX 200301.
Description and comparisonThe M2 (V26970) is similar to that ofAnatolechinosin being sub-quadrate (L × W: 1.30 × 2.00),with low crown and obtuse main cusps,small and low parastyle,short and low metastyle,narrow stylar shelf,incomplete centrocrista,short precrista of protoconule and postcrista of metaconule not extending to the parastyle or metastyle,and the small hypocone joining with posterior cingulum,without lingual cingulum.However,V26970 is different fromAnatolechinosin its postcrista of metaconule extending to the posterior cingulum.It seems that V26970 represents a slightly more progressive form than knownAnatolechinos.
The geological age ofAnatolechinosis Middle-Late Eocene (Qiu and Tong,2015).
Zaraalestes Storch &Dashzeveg,1997
Zaraalestes sp.
(Fig.3E)
SpecimenOne right M2 (IVPP V26971) from LX 200303.
Description and comparisonThe M2 is sub-rectangular in outline,wider than long [L× W: 1.35 × 1.85],with low crown and obtuse main cusps,small and low parastyle,narrow stylar shelf,subequal paracone and metacone,distinct centrocrista,reduced protoconule,and short postcrista of metaconule,which does not meet the posterior cingulum.All these features are as inZaraalestes,but because only one M2 is known,it is difficult to identify the species.
The geological age ofZaraalestesis Middle Eocene to Early Miocene (Storch and Dashzeveg,1997;Ziegler et al.,2007)
Tupaiodontinae gen.et sp.indet.
(Fig.3F-I)
SpecimensFive isolated lower antemolars.LX 200303,four teeth (IVPP V26972.1-4):one right i2/3/c1?,one left p2,one left p3 and one left p4;no field locality number in Yagou:one right p3 (V26973).
Description and comparisonThe right i2/i3/c1? (V26972.1: 1.40 × 0.70) is long oblate-conical in form and procumbent,with an oblate main cusp on the anterior end and a curved posterior part.There is a small cusp on the posterior end.Single root extends posterodownward.
The left p2 (V26972.2: 1.30+× 0.92) is larger than V26972.1 in width and slightly procumbent.The trigonid is triangular-conical with a high main cusp.The low crested talonid has a distinct cuspid.The cingulid is developed along the whole tooth and joins the low talonid.The single root extends posterodownward.
There are two p3 (V26972.3: -× 0.80+and V26973: 1.30 × 0.95).The crown of right p3 (V26973) is well-preserved.It is similar toAnatolechinosin its triangular pyramid-shaped trigonid,low and small paraconid,and the talonid formed by a low transverse crest.The left p3(V26972.3) preserves only part of the crown and one posterior root,showing that the crown is similar to V26973 in morphology.It is clear that the left p3 has two roots.
The left p4 (V26972.4: 1.10+× 0.80+) preserves part of trigonid with protoconid and part of metaconid.The metaconid is located on the lingual side of the high protoconid.The talonid is formed by a low transverse crest.The cingulid extends on the buccal side and continues with the transverse crest on the talonid.
The lower antemolars are similar to those of Tupaiodontinae in basic features.They differ fromAnatolechinosandZaraalestesin p3 and p4 being larger in size,and p3 having higher and more obtuse protoconid;fromZaraalestesin p2 with more developed cingulid;fromIctopidium,AnatolechinosandZaraalestesin p3 having lower talonid.They are designated Tupaiodontinae gen.et sp.indet.for the time being.
The temporal range of Tupaiodontinae is Middle Eocene-Oligocene (Storch and Dashizeveg,1997;Qiu and Tong,2015).
Order Lagomorpha Brandt,1855
Family Leporidae Gray,1821
Ordolagus De Muizon,1977
Ordolagus sp.
(Fig.4A)

Fig.4 Lagomorpha and Rodentia from Yagou area
One broken left upper cheek tooth (IVPP V27881) is collected from Loc.LX 200303.It is compressed longitudinally and wide transversely.It has a curved and high crown without root.On the occlusal surface no hypostria is seen.It may represent an upper cheek tooth ofOrdolagus.It is 1.40 mm in length,close to the P3 ofO.teilhardi.The single tooth cannot be identified at the species level.
The geological age ofOrdolagusis Oligocene (Erbajeva and Daxner-Höck,2014).
Family Ochotonidae Thomas,1897
Desmatolagus Matthew &Granger,1923
Desmatolagus sp.
(Fig.4B-I;Table 2)

Table 2 Measurements of cheek teeth of Desmatolagus sp.from the lower part of the Jiaozigou Formation in Yagou area,Linxia Basin (mm)
SpecimensAltogether 25 cheek teeth.LX 200301,18 cheek teeth (IVPP V26974.1-18):one right P2,four P3,two right P4,one right M1,two M2,four p4/m1/2 and four m3;LX 200303,five cheek teeth (V26975.1-5): two right P4,one right p3,one left p4/m1/2 and one right m3;no field locality number in Yagou: two right p4/m1/2 (V26976.1-2).
DescriptionThe P2 (V26974.1) is oval in occlusal view,wider than long and worn heavily.There are two anterior reentrants (valleys) on the anterior side.The crescentic valley is larger than the buccal valley.The paracone is larger than the protocone.
The other upper cheek teeth are unilaterally hypsodont with a strong lingual main root and distinct buccal roots.P3 has buccally curved crown.The four P3s (V26974.2-5) described here are all worn,trapezoidal in occlusal view.The anteroloph is short and crosses about 1/3 of the anterior border of P3.On the anterior side of the P3 there is a shallow anterior reentrant between the protocone and anteroloph.The large crescentic valley is shorter anteriorly than posteriorly and opens anteriorly.The hypostria may be very shallow or indistinct.The four right P4 described here (V26974.6-7 and V26975.1-2) are all worn on the buccal and lingual sides.However,the well-preserved occlusal surface shows distinct crescentic valley and hypostria.The crescentic valley is closed and has nearly equal long anterior and posterior parts.The hypostria extends to about 2/5 of width of the crown and is opened lingually or closed.The heavily worn M1 and M2 (V26974.8-10) are wide and short,with their crescentic valleys very shallow and hypostriae only represented by enamel circles on occlusal surface.
The lower cheek teeth have high crowns with distinct roots.The p3 (V26975.3) is subtriangular in occlusal view,with a narrow anterior end and a wide and deep hypostriid.The middle lower cheek teeth (p4/m1/2) are similar to each other in morphology.The trigonid is higher and wider than the talonid.The sharp posterior fold of the thicker posterior enamel of the trigonid is located on its buccal 1/3.The m3 is small,with a wide trigonid and a reduced talonid.
All features are similar toDesmatolagus,specially toD.gobiensis.However,the measurements (Table 2) show that the cheek teeth described above are smaller than those ofD.gobiensis,and more important is that the m3 is much more reduced than that ofD.gobiensis.ThisDesmatolagussp.may be more progressive than those ofD.gobiensis.
The geological age ofDesmatolagusis Late Eocene to Miocene (Erbajeva and Daxner-Höck,2014;Li and Zhang,2019).
Sinolagomys Bohlin,1937
cf.Sinolagomys sp.
(Fig.4J)
One large right middle upper cheek tooth (right P4/M1/2: IVPP V27882) is collected from LX 200303.It has a high crown and lacks buccal root.On the occlusal surface no crescentic valley is seen but there is a distinct hypostria extending along the lingual side.All these features are as inSinolagomys.It is similar toS.majorin size (L × W: 2.00 × 3.50),but differs from the known species ofSinolagomysin having a transversely shorter hypostria,which is only about 1/6 of the tooth in width;and the protocone and hypocone are subequal in size.V27882 may represent a primitive species ofSinolagomysor even a more primitive genus thanSinolagomys,but only one tooth is known,so it is considered cf.Sinolagomyssp.
The geological age ofSinolagomysis late Early Oligocene-Early Miocene (Erbajeva and Daxner-Höck,2014,Li and Zhang,2019).
Order Rodentia Bowdich,1821
Family Aplodontidae Brandt,1855
Prosciurus Matthew,1903
Prosciurus sp.
(Fig.4K)
Specimen and measurementOne left M1/2 (IVPP V26977) without field locality number;L × W: 2.00 × 2.20+.
DescriptionThe brachyodont M1/2 (V26977) is incomplete and missing the posterior and lingual borders.The central basin opens buccally,and no ectoloph is seen.The protoloph extends to protocone and meets the posterior border of protoconule.The metaloph meets the anterior border of the metaconule.The metaconule extends to the posteroloph.Both the anteroloph and posteroloph are lower than the protoloph and metaloph.The protocone has a free anterior arm.All these features are similar to those ofProsciurus.Within the genusProsciurusit resemblesP.ordosiusin lacking mesostyle,but differs from it in having more developed protoconule.
The known geological range ofProsciurusis Late Eocene to Early Oligocene in North America and Late Eocene to Late Oligocene in Asia (Matthew and Granger,1923;Kowalski,1974;Wang,1987a,2008a;Emry et al.,1998;Lopatin,2000;Wang and Dashzeveg,2005;Daxner-Höck et al.,2010;Wang and Qiu,2019).
Aplodontidae gen.et sp.indet.
(Fig.4L)
One right upper molar (IVPP V26978) from LX 200303 is broken lingually.It is small(L: 0.75) and has a low crown.The upper molar is similar to that of Aplodontidae in having four transverse lophs,protocone having anterior and posterior arms,and developed metaconule extending to posteroloph.Although it is similar toProsciurusin lacking ectoloph,it differs from all known Aplodontidae in lacking mesostyle and protoconule,but having a conical paracone.It may represent a special genus of Aplodontidae,here designated as Aplodontidae gen.et sp.indet.
Family Mylagaulidae Cope,1881
Subfamily Promylagaulinae Rensberger,1980
Irtyshogaulus Lu et al.,2016
cf.Irtyshogaulus sp.
(Fig.4M)
Specimen and measurementOne right M3 (IVPP V26979) from LX 200303: 1.70 × 1.70.
DescriptionThe M3 is prismatic and mesodont.The occlusal surface is subquadrate in outline,with developed anterior and posterior cingula.There are four large deep fossettes.The protocone is elongated longitudinally to meet the anterior and posterior cingula.The conical paracone is smaller than the protocone.The protoloph extends from the paracone to the protocone.The protoconule extends from the protoloph to the anterior cingulum.The metaconule extends from the lingual part of the protoloph to the posterior cingulum.The metacone is reduced.No metaloph is seen.All features resemble the M3 ofIrtyshogaulus,speciallyI.minor.However,the M3 (V26979) has a low crown.But the lingual crown is higher than the buccal one,with a large lingual main root and two small buccal ones.Probably the M3 (V26979) represents a primitive form of the genus.
The Mylagaulidae are known to range from Late Oligocene to Early Pliocene in North America and Asia (Wu,2019a),and knownIrtyshogaulusis Early Miocene in Asia (Lu et al.,2016;Wu,2019a).
Family Gliridae Muirhead,1819
Glis Brisson,1762
Glis sp.
(Fig.4N)
Specimen and measurementOne left m2 (IVPP V26980) from LX 200303: 1.05 × 1.15.
DescriptionThe small m2 is subquadrate in occlusal view,wider slightly than long,and low crowned.The occlusal surface is slightly concave.The m2 has four main ridges(anterolophid,metalophid,mesolophid and posterolophid) and three extra ridges (anterior extra ridge,centrolophid and posterior extra ridge) (see Daams and de Bruijn,1995:fig.1).The main ridges are well developed and almost transverse.The extra ridges are weaker than the main ridges.Features of V26980 invokeGlis(Daams and de Bruijn,1995;Daams,1999).
Among the known species ofGlis,the m2 (V26980) is similar to that ofG.guerbueziandG.aff.G.guerbuezifrom Turkey in occlusal features (Ünay,1989),but different in being slightly larger in size,the metalophid failing to meet the metaconid,the mesolophid being thinner and separated from the mesoconid,and the mesoconid being isolated.Furthermore,it differs fromG.guerbueziin the main cusps (metaconid,entoconid and hypoconid) being more distinct,the metaconid being larger than the entoconid and the three extra ridges being lower and weaker.It seems that V26980 may represent a form more primitive thanG.guerbuezi.
The genusGlisranges from Late Oligocene to Recent (Daams and de Bruijn,1995;Daams,1999).
Family Eomyidae Winge,1887
Subfamily Eomyinae Winge,1887
Eomyodon Engesser,1987
cf.Eomyodon sp.
(Fig.4O)
Specimen and measurementOne left p4 (IVPP V26981) from LX 200301: 0.75 × 0.75.
Description and comparisonIt is a small eomyid.The p4 is trapezoid in occlusal view,with a slightly concave occlusal surface.It is brachyodont and lophodont.The anterolophid meets the protoconid.The metalophid is slightly curved anteriorly and meets the protoconid.The long mesolophid meets the posterior arm of the protoconid and is as high as the other four main transverse lophids.The hypolophid is transverse and meets the posterior arm of the hypoconid.The ectolophid is interrupted.These features are similar to those ofEomyodon,especiallyE.mayoi(Engesser,1987,1990),but the p4 (V26981) is smaller and relatively shorter but wider than those ofE.mayoi.Thus the p4 (V26981) is identified as cf.Eomyodonsp.
The geological age ofEomyodonis Late Oligocene-Early Miocene in Europe (Engesser,1987).
Family Dipodidae Fischer von Waldheim,1817
Subfamily Zapodinae Coues,1875
Genus Allosminthus Wang,1985
Allosminthus sp.
(Fig.5A-C)

Fig.5 Allosminthus and Parasminthus from Yagou area
SpecimensThree molars from LX 200301: two right M2 (IVPP V27007.1-2) and one left m1 (V27006);and one right M1 (V27002) from LX 200303.
MeasurementsM1: 1.05 × 0.95;M2 (V27007.1): 1.12 × 1.15;m1: 1.28 × 0.84.
Description and comparisonThe molars are low crowned and have obtuse main cusps.The M1/2 are subquadrate in outline and have three roots.On M1/2 the protoloph I is complete and meets the anterior arm of the protocone,the protoloph II is incomplete or absent,and the metaloph meets the hypocone or its anterior arm.On M1 the mesoloph is absent and on the M2 the mesoloph is forked.On the m1 the trigonid is shorter and narrower than the talonid,the anteroconid is small,the posterior arm of the protoconid meets the posterior arm of metaconid to form a complete metalophid II,the mesoconid is present,the mesolophid and ectomesolophid are short,the hypolophid is transverse and the posterolophid reaches the base of the entoconid.
All molar features are similar to those ofAllosminthus,especially to those ofA.ernos,and the sizes are within the range ofA.ernos(Wang,1985,2008a;Li et al.,2017).However,the mesoloph is absent on the M1 and the m1 has a developed anterolophid joining the protoconid and metaconid,which are different from those ofA.ernos.
The geological age of the genusAllosminthushas been extended from Middle Eocene to Late Oligocene (Daxner-Höck et al.,2014;Qiu and Wang,2019)
Genus Parasminthus Bohlin,1946
Parasminthus asiaecentralis Bohlin,1946
(Fig.5J)
SpecimenOne left m3 (IVPP V28426) from LX 200301.
Description and comparisonThe arm of the metaconid of V28426 meets with the anteroconid to form metalophid I.The posterior arm of the protoconid is long and extends to the lingual border of the tooth to form a pseudomesolophid.The hypolophid extends from the entoconid to the ectolophid.The ectolophid is oblique and extends from protoconid to the anterior arm of the hypoconid.The lingual part of the posterolophid extends anteriorly to meet with the entoconid and the metaconid to form a longitudinal crest on the lingual side of m3 and close three sinusids (anteromesosinusid,posteromesosinusid and posterosinusid).All these features plus size of the m3 (1.50 × 1.20) are within the range of variation ofParasminthus asiaecentralis(see Bohlin,1946;Huang,1992).
RemarksWhile describingParasminthus,includingP.asiaecentralis,P.tangingoliandP.parvulus,from Yindirte (=Taben Buluk),Gansu,Bohlin (1946) considered the geological age as Late Oligocene.This viewpoint has been affirmed by Wang (2003).Later,more fossils ofParasminthuswere described from the Ulantatal area of Nei Mongol,Gansu,Xinjiang and Qinghai.The geological age of the genusParasminthushas been extended from Early Oligocene to Early Miocene (Qiu and Wang,2019) but the age ofP.asiaecentralis,P.tangingoliandP.parvulusremains Oligocene.
Parasminthus tangingoli Bohlin,1946
(Fig.5D-I;Table 3)

Table 3 Measurements of cheek teeth of Parasminthus tangingoli from the lower part of the Jiaozigou Formation in Yagou,Linxia Basin (mm)
ParasminthusdebruijniLopatin,1999,p.432
SpecimensAltogether 46 cheek teeth.LX 200301,26 cheek teeth (IVPP V26982.1-26):two left P4,four M1,five M2,five m1,seven m2 and three m3;LX 200302 (V26983.1-2): left m1 and right m2;LX 2000303,18 cheek teeth (V26984.1-18): two left P4,one left M1,three M2,two left m1,seven m2 and three right m3.
Description and comparisonThe P4 is small and conical in form and with a main cusp and a posterior cingulum.The molars have distinct main cusps.On M1/2 the mesoloph is developed.On M1 the protoloph joins the posterior arm of protocone and the metaloph joins the hypocone or its posterior arm.On M2 the protoloph may be double or single and the metaloph joins with the hypocone or its anterior arm.The posterior concavity between the hypocone and posteroloph is developed on M1,and may be distinct or indistinct on M2.The m1 usually has developed anteroconid and mesolophid,and subequal protoconid and metaconid.On m2/3 the metalophid I is complete and the metalophid II and mesolophid may be developed or not.All of the features mentioned above occur inParasminthustangingoli(Bohlin,1946;Huang,1992;Wang and Qiu,2000a).In addition,the size of the cheek teeth is within the range of the variation ofP.tangingoli(see Table 3;Huang,1992).
MeasurementSee Table 3.
DiscussionLopatin (1999) described a new species ofParasminthus,P.debruijni,from North Aral Region.He mentioned features ofP.debruijnias differing fromP.asiaecentralisandP.tangingoli: “the smaller size,the absence of contact between metaloph II and the posteroloph on M1 and M2,reduced anteroconid on m1 and reduced mesolophid on m2 and invariably a four-rooted M1 and M2”.Having compared the specimens ofP.debruijniwith those ofP.tangingoli,we found that all of the “different features” ofP.debruijnimentioned by Lopatin (1999) are similar to those ofP.tangingoli: 1)P.debruijniis within the range ofP.tangingoliin size (see Lopatin,1999:table 2;Huang,1992:tables 8,10);2) the features of M1 and M2 ofP.debruijniare similar to those of holotype ofP.tangingoli(Bohlin,1946:fig.2: 7,8);3) the other features of molars ofP.debruijnifall within the range exhibited byP.tangingoli(see Bohlin,1946:23-30;Huang,1992:255-260).It seems to us thatP.debruijniis a junior synonym ofP.tangingoli.
According to Lopatin (1999),the fossils of “Parasminthusdebruijni” were collected from the Early Miocene Aral Formation of North Aral Region.However,Lucas et al.(1998)considered the age of the Aral Formation as Late Oligocene based on the Aral local fauna,soParasminthustangingoli(=Lopatin’s “Parasminthusdebruijni”) from the Aral Formation likely is also Late Oligocene in age.Daxner-Höck et al.(2014:170) also thought that Lopatin’s“Parasminthusdebruijni” is Late Oligocene in age.Thus,in Asia the geological age ofParasminthustangingoliis Oligocene only.
Parasminthus parvulus Bohlin,1946
(Fig.5K-O;Table 4)
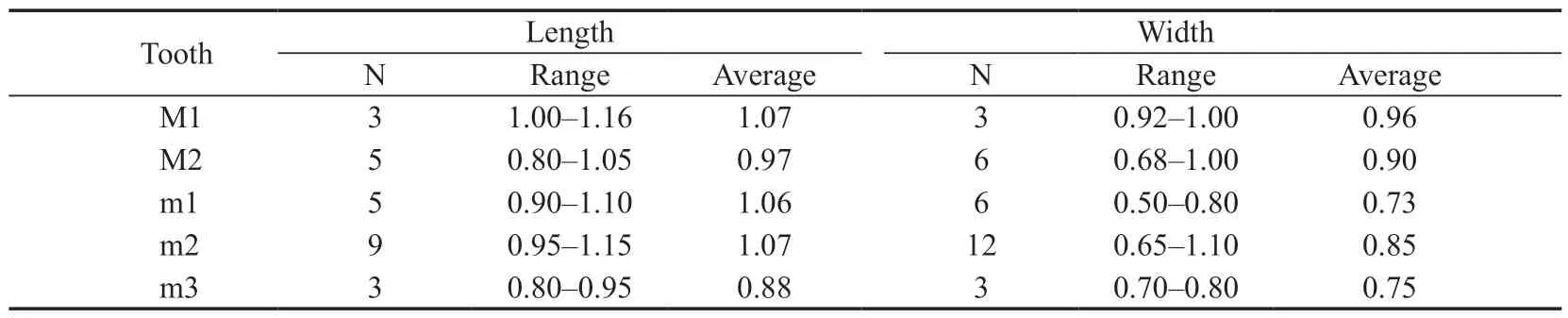
Table 4 Measurements of cheek teeth of Parasminthus parvulus from the lower part of the Jiaozigou Formation in Yagou,Linxia Basin (mm)
SpecimensAltogether 31 cheek teeth.LX 200301,22 cheek teeth (V26985.1-22): three M1,three M2,five m1,nine m2 and two m3;LX 200303,five lower molars (V26986.1-5):one right m1,three m2 and one right m3;no field locality number in Yagou,four molars(V26987.1-4): three M2 and one right m1.
Description and comparisonThe cheek teeth described here represent a small species ofParasminthus.As inP.tangingoli,the mesoloph of M1/2 is developed,but without posterior concavity between the hypocone and posteroloph on M1/2.The metaloph of M1 extends transversely and joins with the hypocone.On M2 only proroloph I is present and the metaloph joins with the anterior arm of the hypocone.On m1 the anteroconid may be present or absent.On the m2/3 the metalophid I is complete,metalophid II and mesolophid are variable.All of the features described above are similar to those ofP.parvulus,rather thanP.asiaecentralisandP.tangingoli.In addition,the size of cheek teeth described here is within the range of variation ofP.parvulus(Huang,1992).
MeasurementSee Table 4.
DiscussionLopatin (1999) referredParasminthusparvulusto his new genusBohlinosminthus.However,López-Antoñanzas and Sen (2006) thought thatBohlinosminthusis a junior synonym ofParasminthus.We follow their opinion as to the synonymy ofBohlinosminthuswithParasminthus.
Family Ctenodactylidae Gervais,1853
Tataromys Matthew &Granger,1923
Tataromys plicidens Matthew &Granger,1923
(Fig.6A-E)
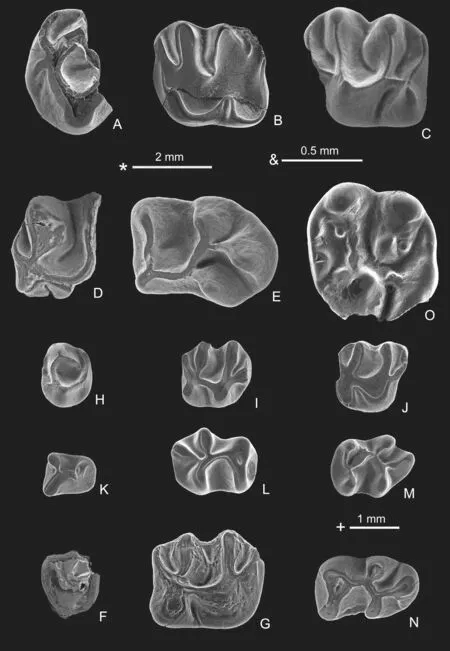
Fig.6 Tataromys and Gobiomys from Yagou area
SpecimensTwelve teeth.LX 200301,four molars (IVPP V26989.1-4): one left M1/2 and three right m1/2;LX 200302,three upper molars (V26990.1-3): two M1/2 and one left M3;LX 200303,five cheek teeth (V26991.1-5): one right P4,three M1/2 and one right M3.
MeasurementsP4: -× 3.00;M1/2: 2.90 × 2.70,2.70 × 2.50,2.90 × -,3.20 × 2.80,-×2.85;M3: -× 2.40,-× 2.65;m1/2: 3.40 × 2.60,-× 2.10,3.70 × 2.70.
Description and comparisonThe cheek teeth are moderately high-crowned.The right P4 (V26991.1) is broken anteriorly.The protocone is higher than the paracone and compressed anteroposteriorly.The metaloph is curved anteriorly.The developed posterior cingulum joins the metaloph at its middle and is divided into buccal and lingual parts.The M1/2 is roughly quadrate in occlusal view.The paracone and metacone are compressed anteroposteriorly.The protocone is V-shaped and larger than the hypocone.There are four transverse lophs,which are nearly parallel to each other.The lingual part of the metaloph curves slightly anteriorly to meet the posterior arm of protocone and the mesosinus is wide U-shaped.The anterosinus and posterosinus extend transversely.Of the two broken M3 the posteroloph is short and the metaloph extends slightly posteriorly to join the hypocone.The lower molars are nearly rectangular with a convex posterior border in occlusal view.The trigonid is very short,with or without small trigonid basin.The ectolophid is straight and lingually situated.The arms of entoconid,hypoconid and hypoconulid are curved.All of the features are similar to those ofTataromys plicidensand the cheek teeth are within the range ofT.plicidensin size (Wang,1997b:16) and fit exactly the diagnosis ofT.plicidens.
TataromysandT.plicidensare Early-Late Oligocene in Asia (Wang,1997b,2019a).
Tataromys sigmodon Matthew &Granger,1923
(Fig.6F,G)
SpecimensOne right P4 (IVPP V26992) from LX 200301;one right M1/2 (V26993)from LX 200303.
DescriptionThe right P4 (V26992) is broken.The protoloph joins the paracone.The posterior arm of the protocone meets the posterior arm of the paracone to form the metaloph.The posterior cingulum meets the metaloph to form a closed buccal basin after wear.No lingual part of the posterior cingulum is seen.It is similar to P4 ofTataromys sigmodonin morphology.The right M1/2 (V26993) is also roughly quadrate in occlusal surface.The paracone and metacone are compressed anteroposteriorly.The protocone is L-shaped with the posterior arm extending nearly longitudinally.There are also four transverse lophs.However,the metaloph curves posteriorly to join the posteroloph,so the mesosinus is L-shaped and the posterosinus is short.All features are as inT.sigmodonrather thanT.plicidens.In addition,the P4 (V26992: 1.40 × 1.60+) and M1/2 (V26993: 2.65 × 2.30) are smaller than those ofT.plicidens,but within the variation range ofT.sigmodon.
Geological range ofT.sigmodonis Oligocene of Asia (Wang,1997b,2019a).
Tataromys minor (Huang,1985)
(Fig.6H-N;Table 5)
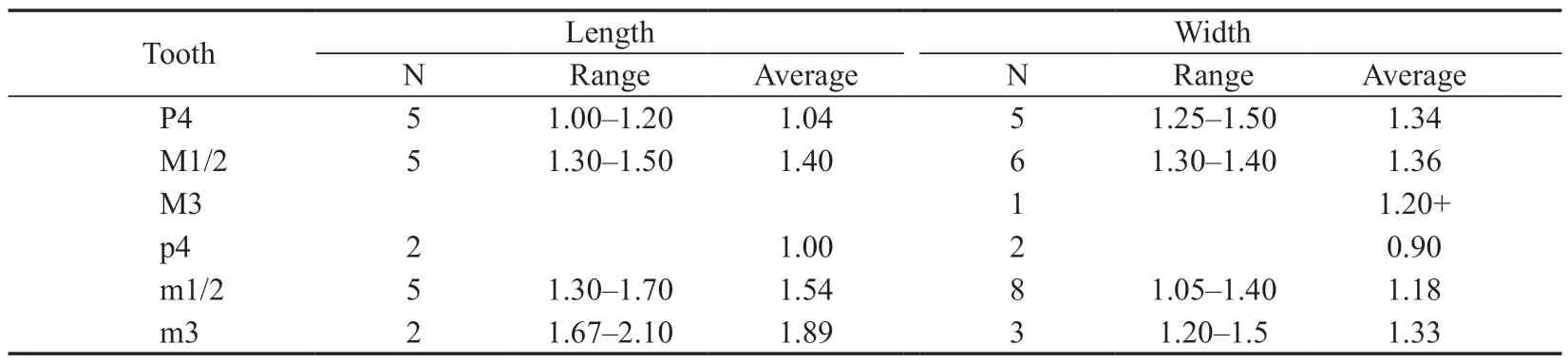
Table 5 Measurements of cheek teeth of Tataromys minor from the lower part of the Jiaozigou Formation in Yagou area,Linxia Basin (mm)
SpecimensAltogether 29 cheek teeth.LX 200301,17 cheek teeth (IVPP V26995.1-16 and V26994): three P4,three right M1/2,one right M3,two p4,six m1/2 and two left m3;LX 200302: one left M1/2 (V26996);LX 200303,11 cheek teeth (V26997.1-11): two P4,six M1/2,two m1/2 and one left m3.
DescriptionThe cheek teeth described here have compressed main cusps and lophs.On P4 the protocone is V-shaped.The protoloph is complete.The metaloph extends posterolingually and usually joins with the posterior arm of protocone to close the trigon basin.The anterior and posterior cingula usually are present.On the upper molars the protocone is wide V-shaped,with the posterior arm extending rather longitudinally and meeting the hypocone.The protoloph is developed and extends to the protocone.The metaloph slightly bends posteriorly and joins the hypocone or posteroloph.Thus the mesosinus is wide L-shaped.The anterosinus and posterosinus are short transversely and the posterosinus is separated from the sinus.The anterocone may be distinct or indistinct.On the p4 the trigonid is higher and wider than the talonid,protoconid larger than the metaconid.The entoconid is as distinct as the metaconid.The hypoconid is absent or weakly developed as a lower transverse crest.On the lower molars the trigonid is short,with protoconid and metaconid compressed anteroposteriorly.The straight metalophid II shifts anteriorly to fuse with the metaconid.Thus,the trigonid basin is small and closed.The ectolophid is short and situated lingually.The arm of the entoconid extends anterobuccally to meet the ectolophid.The arm of the larger hypoconulid extends anteriorly to meet the long and curved arm of the hypoconid.
All of the features of the cheek teeth described above are similar to those ofTataromys minorand they are close in size (see Table 5).
MeasurementsSee Table 5.
RemarksWang (1997b) erectedTataromys parvus.Based on more specimens from Ulantatal area,Nei Mongol,Vianey-Liaud et al.(2006:132) thought thatTataromysparvusis a junior synonym ofTataromysminor.We agree and follow their idea here.
T.minoris known from Early to Late Oligocene (Wang,1997b,2019a;Vianey-Liaud et al.,2006).
Family Gobiomyidae Wang,2001
Gobiomys Wang,2001
cf.Gobiomys sp.
(Fig.6O)
Specimen and measurementOne left M1/2 (IVPP V26988) from LX 200301: 0.80 ×0.85.
Description and comparisonThe M1/2 (V26988) is similar toGobiomysin being wider than long,having brachydont crown,distinct main cusps,thin protoloph without protoconule,developed metaconule,incomplete metaloph extending anterolingually,weak entoloph,developed anterior and posterior cingula.However,it differs fromGobiomysin having accessory cusps mesial to protoloph and distal to metaconule.Besides,it is smaller than all the known species ofGobiomys,includingG.exiguus.The M1/2 (V26988) may represent a new species.
The geological age ofGobiomysis Middle-Late Eocene (Wang,2001b,2019b).
Family Tsaganomyidae Matthew &Granger,1923
Tsaganomys Matthew &Granger,1923
Tsaganomys altaicus Matthew &Granger,1923
(Fig.7)

Fig.7 Tsaganomys altaicus from Yagou area
Specimens and measurementOne left upper cheek tooth and one right upper incisor(IVPP V26998.1-2) from LX 200301;and one left upper cheek tooth (V26999) from LX 200302.
Cheek teeth: 2.80 × 4.70,2.60 × 3.50;I2: -× 6.50 (mm).
Description and comparisonBoth V26998.1 and V26999 are isolated upper cheek teeth.They are oblate cylindrical in form and strongly unilaterally hypsodont.The occlusal surfaces are slightly concave with wearing and featureless.The enamel covers the anterior and lingual wall on upper cheek teeth.Each cheek tooth has a well-developed dentine part composed of primary dentine and a short conical pulp cavity with pointed apex.The right upper incisor preserves only part of labial enamel,which is slightly convex longitudinally and nearly flat transversely.On the labial surface there are distinct longitudinal ridges.All these features suggestTsaganomysaltaicusfrom Oligocene of Asia (Wang,2001a,2019c).
Family Cricetidae Fischer von Waldheim,1817
Subfamily Eucricetodontinae Mein &Freudenthal,1971
Eucricetodon Thaler,1966
In the Yagou area the genusEucricetodonincludes four species:E.asiaticus,E.bagus,E.caducusandE.jilantaiensis.They share such generic features ofEucricetodonas: brachyodont molars with bunodont cusps and simple lophs;M1 with simple prelobe,variable anterocone and complete protoloph II,M1/2 with short mesoloph;m1 with simple anteroconid,metaconid located more anteriorly than the protoconid,complete metalophid II,distinct mesoconid,short mesolophid and ectomesolophid,and entolophid extending to the anterior arm of the hypoconid.
The temporal range ofEucricetodonis Late Eocene to Early Miocene,but most species are Oligocene in Asia (Gomes Rodrigues et al.,2012;Wang et al.,2020).
Eucricetodon asiaticus (Matthew &Granger,1923)
(Fig.8A-E)
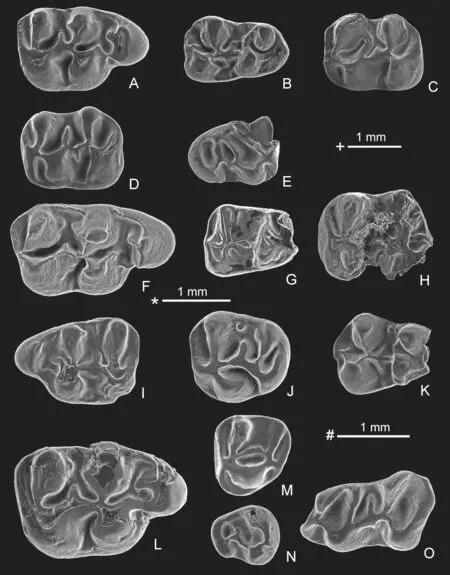
Fig.8 Eucricetodon,Bagacricetodon and Cricetidae gen.et sp.indet.from Yagou area
SpecimensAltogether 12 cheek teeth.LX 200301,seven molars (IVPP V27000.1-7):two right M1,three m1,one left m2 and one left m3;LX 200303,five molars (V27001.1-5):one right m1,two left m2 and two m3.
Description and comparisonThe prelobe of M1 is simple and has one anterocone and a short anterocone spur.On M1 the protocone has a free anterior arm.The metaloph meets the hypocone.The m1 has a developed anteroconid.The m1/2 has a small but distinct hypoconulid.The metalophid II of the m2/3 may be long or short.All these features of the molars are those ofEucricetodonasiaticusand they are within the range of variation of the molars ofE.asiatcusin size (Lindsay,1978;Gomes Rodrigues et al.,2012).
MeasurementsM1: 2.35 × 1.50,2.45 × 1.60;m1: 2.10 × 1.30,2.00 × 1.35,1.95 × 1.20,2.05 × 1.35;m2: 1.80 × 1.35,1.75 × 1.50,1.90 × 1.50;m3: 1.75 × -,1.75+× 1.26.
Eucricetodon bagus Gomes Rodrigues et al.,2012
(Fig.8F-H)
SpecimensFive molars (IVPP V27003.1-5): three M1,one right m1 and one left m2 from LX 200303.
MeasurementsM1: 2.25 × 1.30,2.05+× -,2.25 × 1.40;m1: -× 1.05;m2: 1.75+× 1.30.
Description and comparisonOn M1 the anterocone has a tendency to split into two.There is also a tendency to form double connection between the anterocone and protocone by the anterolophule and anterior arm of the protocone.The metaloph connects with the posterior arm of the hypocone.On the m1 the anteroconid is broken away but the other part is well preserved: the metaconid is located more anteriorly than the protoconid and the metalophid II is present;the mesoconid is distinct and mesolophid and ectomesolophid are short;the entolophid extends to the anterior arm of the hypoconid.The m2 is poor preserved,but it shows that the metalophid I is complete,the posterior arm of the protoconid is short and no mesolphid is seen.The molars described here are similar to those ofEucricetodonbagusin features and are within or close to those ofE.bagusin size (Gomes Rodrigues et al.,2012).
The geological range ofE.bagusis Oligocene in Asia (Gomes Rodrigues et al.,2012,2014;Wang et al.,2020).
Eucricetodon caducus (Shevyreva,1967)
(Fig.8I-K)
SpecimensFour molars (IVPP V27004.1-4): one left M1,one left M2,and two right m1 from LX 200301.
MeasurementsM1: 1.75 × 1.20;M2: 1.40 × 1.20;m1: 1.50+× 1.05,1.50+× 1.15).
Description and comparisonThe molars are small in size.The prelobe of M1 has a wide arched anterocone.M1 has double protoloph I and II,which are low and short,and nearly enclose a small proto-fossetus.On M2 single protoloph (=protoloph I) is present and the metaloph extends to the anterior arm of the hypocone.The m1 has a low anteroconid.All these features of the molars are similar to those ofEucricetodoncaducusand size range of variation is like that of other samples (Shevyreva,1967;Wang,1987b).
The geological age ofE.caducusis early Oligocene in Asia (Shevyreva,1967;Wang,1987b;Wang et al.,2020),and that ofEucricetodonaff.E.caducusfrom Junggar basin is Late Oligocene (Maridet et al.,2009).According to Maridet et al.(2009:11),the molar morphology and size of the holotype ofE.caducusare within of variation ofEucricetodonaff.E.caducus,so the Junggar material may be referred toE.caducuswith age spanning Early to Late Oligocene.
Eucricetodon jilantaiensis Gomes Rodrigues et al.,2012
(Fig.8L-M)
SpecimensLX 200303: one right M1 (IVPP V27005);LX 200301: two left M3(V30714.1-2).
Description and comparisonThe M1 has a large prelobe,a simple antercone,and a strong anterolophule extending to the protocone.The protocone has no distinct free anterior arm.The metaloph extends slightly posterolingually to meet the posterior arm of the hypocone.On M3 the metacone and hypocone are much smaller than the paracone and protocone,the entoloph joins with protoloph and the posterior arm of protocone or metaloph to enclose a small pit and the sinus is small.All of the features of the upper molars are similar to those ofEucricetodonjilantaiensis.In addition,the M1 (V27005: 2.40 × 1.65) and two M3 (1.06 × 1.10;1.05 × 1.06) are within the range of variation ofE.jilantaiensisin size (see Gomes Rodrigues et al.,2012).
Bagacricetodon Gomes Rodrigues et al.,2012
Bagacricetodon sp.
(Fig.8N)
Specimen and measurementOne right m3 (IVPP V27008) from LX 200301: 0.95 × 0.85.
DescriptionThe m3 is brachyodont.The anterocingulid is absent.The anteroconid is located at the middle of the anterolophid.The anterior arm of the metaconid joins the anterolophid.The posterior arm of the protoconid is absent.The entoconid is indistinct.Both the ectolophid and sinusid extend posterolingually.These features are similar to those ofBagacricetodontongiexcept for smaller size.
The age ofBagacricetodonis Late Oligocene (Gomes Rodrigues et al.,2012,2014;Wang et al.,2020).
Cricetidae gen.et sp.indet.
(Fig.8O)
Specimen and measurementOne left m1 (IVPP V27009) from LX 200301: 1.70 × 1.10+.
DescriptionThe m1 (V27009) is moderately high crowned and lophodont.It is similar toPlesiodipusrather than to other genera of the Cricetidae in the anteroconid being narrow transversely,the anterior lingual main cusp (=metaconid) being compressed obliquely and located more anteriorly than the buccal one (=protoconid),having an anterobucally extending transverse lophid (=metalophid I),and lacking mesoconid.However,it differs fromPlesiodipusin its developed metalophid II and mesolophid,distinct ectomesolophid and long sinusid.At present,this molar can not be referred to any known genus of the Cricetidae.
3 Discussion
The Cenozoic fluviatile-lacustrine deposits of Linxia were named by the Regional Geological Survey Team of Gansu Province as Linxia Formation in the geological map(1:200000) of the Linxia area (RGSTGP,1965).This formation was originally considered Pliocene in age and subdivided into four unnamed members.Qiu et al.(1990) first reported some large-mammal fossils obtained from the lower part of the Linxia Formation at Jiaozigou,Dongxiang Autonomous County,includingGomphotheriumsp.,Dzungariotheriumorgosense,Rhinocerotidae gen.et sp.indet.andParaentelodonmacrognathus.Based on these fossils,they claimed the fossiliferous part of the Linxia Formation to be Early Miocene in age and renamed the lower part (including the first and second members) as Jiaozigou Formation.Later,while measuring the Maogou section where the same deposits are better exposed,geologists of the Lanzhou University subdivided this part into Late Oligocene Tala Formation and Early Miocene Zhongzhuang Formation (Fang et al.,1995,1997;Li et al.,1995).In 2004,mainly based on newly discovered large-mammal fossils,including some indricotheres(Dzungariotherium orgosenseandParaceratherium yagouense),and noting that the previously reportedGomphotheriumsp.might not have come from the Jiaozigou Formation,Deng et al.(2004) and Qiu et al.(2004a,b) reassigned Qiu et al.’s (1990) Jiaozigou Formation as Oligocene in age.They further considered the deposits exposed in Jiaozigou,Maogou and Yagou areas as mainly Late Oligocene.Recently,this opinion has been further corroborated by Deng et al.(2021).
The micromammal fossils described here were collected from the basal part of the Jiaozigou Formation in the Yagou area in 2003.All 17 genera are previously known and well dated.Among them 14 genera are known to occur in Oligocene deposits.Four of the 14 Oligocene genera (Ordolagus,Tataromys,TsaganomysandBagacricetodon) and four species ofEucricetodon(E.asiaticus,E.bagus,E.caducusandE.jilantaiensis) are known from Oligocene deposits only.Bagacricetodonis only Late Oligocene,andGlisandEomyodonmade their first appearances in Late Oligocene.All these data are in good accordance with the existing opinion that the basal part of the Jiaozigou Formation is mainly Late Oligocene in age(Fig.9).
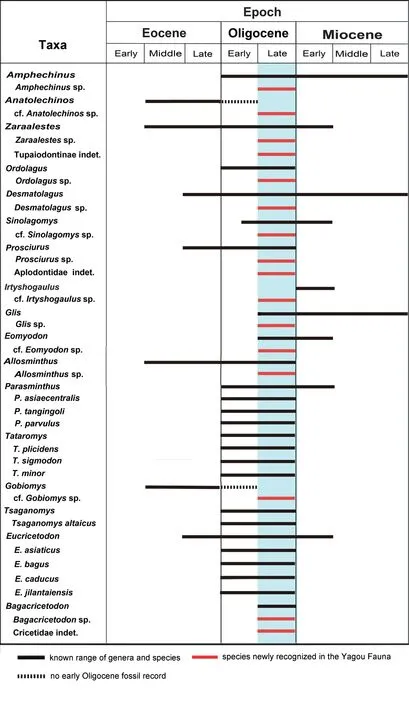
Fig.9 Chronological range of micromammals from the basal part of the Jiaozigou Formation in Yagou area,Linxia Basin
The three remaining genera constrain the age less rigorously:AnatolechinosandGobiomyshave been known to be Eocene,but our referred fossils are more progressive than the known species,so quasi-consistent with Late Oligocene age.Likewise,Irtyshogaulusis mainly Early Miocene,but our fossil species is more primitive than any knownIrtyshogaulus.Therefore,the Late Oligocene age of the basal part of the Jiaozhigou Formation in Yagou area can thus be considered substantiated.
Five Late Oligocene faunas are well known in China (Bohlin,1937,1942,1946;Wang et al.,1981;Wang,1997a;Wang and Qiu,2000b;Meng et al.,2006): Shargaltein-Tal,Tabenbuluk,Yikebulage and Xiagou faunas and Tieersihabahe assemblage.In comparison with these faunas,the Yagou Fauna seems to be slightly more primitive in evolutionary level.For example,cf.Sinolagomyssp.in the Yagou Fauna is more primitive thanSinolagomys kansuensisfound in the other five faunas.On the other hand,some relatively advanced genera,such asTachyoryctoides,Yindirtemys,HeterosminthusandLitodonomysare present in all the above mentioned faunas,but absent in the Yagou Fauna.On the contrary,the Yagou Fauna contains some genera of Eocene age,such asAnatolechinosandGobiomys,and several archaic species ofEucricetodon,which are so far not found in the other faunas.Taken as a whole,the Yagou Fauna may be slightly older than the other known Late Oligocene faunas.
The Shargaltein-Tal Fauna is closest to the Yagou Fauna in preserving the archaic generaTataromysandTsaganomys,but lacking some advanced taxa found in the others.
Recently,some other Oligocene assemblages,including Ulan I-III biozones in Nei Mongol of China,and biozones A-C1 in Valley of Lakes of Mongolia have been reported.The Ulan I and II biozones and biozones A and B are referred to Early Oligocene and the Ulan III biozone and biozones C and C1 are assigned to Late Oligocene (Vianey-Liaud et al.,2006;Daxner-Höck et al.,2010,2017;Gomes Rodrigues et al.,2012,2014;Zhang et al.,2016;Wasiljeff et al.,2020;Wasiljeff and Zhang,2022).The Yagou Fauna shares with Ulan III biozone and biozone C such genera asDesmatolagus,Tataromys,Parasminthus,Tsaganomys,EucricetodonandBagacricetodon.However,in the Yagou Fauna the genera of Eocene origin,such asAnatolechinosandGobiomys,are absent from the above mentioned biozones.At the same time,in both biozones there are some other advanced genera,such asYindirtemys,Litodonomys,PlesiodipusandTachyoryctoides,which are known extending into the Miocene,but absent from the Yagou Fauna.Obviously,the Yagou Fauna may be slightly older than either Ulan III biozone or biozone C in age.
In comparison with the two Early Oligocene faunas (biozone B in Valley of Lakes of Mongolia and UTL II biozone in Ulantatal area) the Yagou Fauna is similar to them in havingDesmatolagus,Sinolagomys,Tataromys,Tsaganomys,AllosminthusandParasminthus,but differs in lacking such older genera asKarakoromys,Bounomys,Anomoemys,HeosminthusandShamosminthus.Obviously the Yagou Fauna is younger than biozone B or ULT II biozone in age.If so,the Yagou Fauna may represent a fauna of an age ranging between the early and late Oligocene and so far unknown from China and Mongolia.It may be the oldest Late Oligocene Fauna or possibly partly latest Early Oligocene.According to recent magnetostratigraphic study (Zheng et al.,2023),the lower boundary of the Jiaozigou Formation in the Maogou section is calibrated to the base of Chron C10r,ca 29 Ma,which is a little older than the layer bearing the Yagou Fauna.
It is well known that the late Middle Eocene-Early Oligocene interval was one of the most significant episodes of Tertiary climatic changes in Asia (Wolfe,1978).Global paleoclimate gradually cooled and dried,with increasing mean annual range of temperature and steepening latitudinal temperature gradient (Wolfe,1978;Berggren and Prothero,1992;Retallack,1992),and the vegetation changed from dense forests to open forested grassland(Prothero and Heaton,1996).In Asia the perissodactyl-dominant fauna of the Eocene was replaced by rodent/lagomorph-dominant faunas of Oligocene (Meng and McKenna,1998).The faunal turnovers were effected by major climatic changes: from a warm and humid Eocene to a cool and arid Oligocene.
The Yagou micromammal fauna witnessed and materially reflected the changed cool and arid climate of the Late Oligocene.The majority of the Yagou micromammal fauna,13 (~76%)of the 17 genera and 175 (93%) of the 189 teeth,belongs to variously xerophilous terrestrial mammals.The lagomorphs,zapodines,ctenodactylids and cricetids are the best representatives.The living ctenodactylids are well known as xerophilous animals,well adapted to dry climate and desert or semidersert landscape.This family flourished and diversified in other Asian Oligocene localities as well (Wang,1997b).Tataromys,as analyzed by Wang (1997b:65): “may already have more developed visual and auditory organs,adapting to wide desert or wilderness as the living ctenodactylids do,have a well-developed masseter medialis muscle,especially the anterior part,and have strong capability of gnawing and chewing,and have more developed temporalis muscle and a lower cheek tooth crown than living Ctenodactylinae,showing thatTataromyswas probably more omnivorous”.The living lagomorphs,zapodids and the cricetids are known to flourish and diversify in dry and semiarid grassland,shrub-grassland or forestgrassland in the temperate Holarctic Region (Luo,1988;Qiu and Li,2016;Qiu and Wang,2019;Chen and Gao,2000),although the cricetids are more or less omnipresent in various environments.Besides,TsaganomysandIrtyshogaulusmay be adapted for a fossorial mode of life and may feed on insects and worms (Wu,2019a;Wang,2001a).This is in good accordance with the arid and semiarid woodland habitat of the giant rhinos paraceratheres proposed by Qiu and Wang (2007).
On the other hand,the fossil-bearing deposits,the Jiaozigou Formation in the Yagou area is mainly composed of reddish brown mudstone,implying an arid and semiarid climate.
Based on the mammal fauna and lithology,the Yagou area can thus be postulated as semiarid woodland-shrubland,an environment suitable for subsistence of the giant-rhinos and ctenodactylid-zapodines rodents during the Late Oligocene.
The Yagou micromammal fauna also shed some light on the faunal intercontinental exchanges.It is interesting to note that in the Yagou area four genera (Amphechinus,Glis,EomyodonandEucricetodon) (about 24%) are found common to Asia and Europe,but only one genus (Prosciurus) is common to Asia and North American.It seems that during the Late Oligocene more frequent faunal interchanges might have occurred between Asia and Europe than between Asia and North America.This bias may directly be affected by the disappearance of the Turgai Strait during this time period.
AcknowledgementsThe authors would like to express their gratitude to Profs.Qiu Zhuding,Zhang Zhaoqun,Wu Wenyu,Li Qian and Dr.Li Lu from Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences (IVPP) for their valuable discussion;to Prof.Deng Tao of IVPP for his valuable geological information on the Jiaozigou Formation and field photographs;to Mrs.Feng Wenqing and Liang Zhong of IVPP and Chen Shanqin of Hezheng Paleozoological Museum of Gansu for their hard work in collecting micromammals in field.Many thanks are given to Dr.Larry Flynn from Harvard University,USA,for critiquing the manuscript and correcting the English.Many thanks are also given to Miss Si Hongwei of IVPP for the microphotographs and preparing the pictures and Mr.Gao Wei of IVPP for the photographs,and Mr.Xie Shuhua of IVPP for the preparation work on the specimens.This work is supported by the Frontier Science Key Research Project of the Chinese Academy of Sciences (QYZDY-SSW-DQC022).

- 古脊椎动物学报(中英文)的其它文章
- New findings of Xiyuichthys (Xiushuiaspidae,Galeaspida)from the Silurian of Jiangxi Province and Tarim Basin
- Reappraisal of Bothriolepis sinensis Chi,1940 from the Tiaomachien Formation,Hunan,China
- A giant bamboo rat from the latest Miocene of Yunnan
- An egg clutch of the Stalicoolithidae discovered in Wuning,Jiangxi,China