
New findings of Xiyuichthys (Xiushuiaspidae,Galeaspida)from the Silurian of Jiangxi Province and Tarim Basin
2023-12-08 02:42SHANXianRenLINXiangHongZHANGYuMengLIXuTongGAIZhiKun
古脊椎动物学报(中英文) 2023年4期
SHAN Xian-Ren LIN Xiang-Hong ZHANG Yu-Meng LI Xu-Tong GAI Zhi-Kun*
(1 Key Laboratory of Vertebrate Evolution and Human Origins of Chinese Academy of Sciences, Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences Beijing 100044)
(2 University of Chinese Academy of Sciences Beijing 100049)
(3 Yunnan Key Laboratory for Palaeobiology,Institute of Palaeontology,Yunnan University Kunming 650500)
(4 College of Life Sciences,Jiangxi Normal University Nanchang 330022)
Abstract New findings of the early Silurian Xiyuichthys (Xiushuiaspidae,Galeaspida),Xiyuichthys lixiensis sp.nov.and X.zhangi are described from the Qingshui Formation in Jiangxi Province and the Tataertag Formation in Tarim Basin respectively.X.lixiensis sp.nov.is characterized by the partially serrated lateral margin of the headshield and the ornamentation composed of extremely coarse granular tubercles (one tubercle per square millimetre).The complete early Silurian biostratigraphic sequence in northwestern Jiangxi warrants the erection of a standard section for the correlation of Silurian shallow marine red beds in South China and Tarim blocks.Thus,the finding of X.lixiensis from the Qingshui Formation (Silurian Lower Red Beds) in Jiujiang of Jiangxi bears important biostratigraphic significance.It can directly compare to X.zhangi from the Tataertag Formation in Tarim Basin on the specific level,which corroborates the correlations between the Tataertag Formation in Tarim Block and the Silurian Lower Red Beds in South China.Fossil records suggest that Xiushuiaspidae have a relatively broader stratigraphic range,but exhibit distinct composition at different stratigraphic horizons,with Xiyuichthys and Changxingaspis arising in the Silurian Lower Red Beds (Qingshui,Tangchiawu,and Tataertag formations) and Xiushuiaspis occurring in the Silurian Upper Red Beds (Xikeng Formation).
Key words northwestern Jiangxi,Tarim Basin;early Silurian;Qingshui Formation,Tataertag Formation;Galeaspids;biostratigraphy
1 Introduction
Galeaspida is a diversified jawless clade known exclusively in the Silurian-Devonian strata of Tarim,North China,and South China blocks (Janvier,1996;Zhao,2005;Zhu and Gai,2006;Janvier et al.,2009;Zhao et al.,2009;Gai et al.,2018).Fossil records suggest that galeaspids underwent the first diversification in Tarim and South China blocks during the early Telychian,with the occurrence of three basal clades Dayongaspidae,Hanyangaspidae,and Xiushuiaspidae,the most basal clade of Eugaleaspiforme (Shuyuidae),and the most basal clade of Polybranchiaspiforme (Gumuaspidae) (Shan et al.,2022b).The Xiushuiaspidae is erected based on the type genusXiushuiaspisincluding two species,X.jiangxiensisandX.ganbeiensis,from the Xikeng Formation in Xiushui County,Jiangxi Province (Pan and Wang,1983).Wang (1991) described the second member of Xiushuiaspidae,Changxingaspisgui,from the Tangchiawu Formation (assumed as the Maoshan Formation in the original paper)in Changxing County,Zhejiang Province.Lu et al.(2007) described ‘Nanjiangaspiszhangi’based on an incomplete headshield from the Tataertag Formation in Kalpin of Xinjiang,and assigned the species into Hanyangaspidae.However,Liu et al.(2015) considered that ‘N.zhangi’ is more suggestive ofKalpinolepisof Hanyangaspidae thanNanjiangaspisbecause its headshield ornamentation is composed of small granular tubercles,a condition much like that ofKalpinolepis,but obviously different from the snow-like tubercles of the type species ofNanjiangaspis(Wang et al.,2002).The new materials of ‘N.zhangi’ revealed that its headshield is oval as that of Xiushuiaspidae rather than trapezoidal as that of Hanyangaspidae(Liu et al.,2019).Therefore,‘N.zhangi’ was removed from Hanyangaspidae to Xiushuiaspidae,and represented the third genus of Xiushuiaspidae,Xiyuaspis(useXiyuichthysforXiyuaspisin this paper) (Liu et al.,2019).
A diversified galeaspid fauna including several new species of Dayongaspidae,Xiushuiaspidae,Gumuaspidae,and Eugaleaspiforme from the Qingshui Formation in Jiujiang of Jiangxi Province has been reported (Shan et al.,2022a,b).However,apart from three new species of Eugaleaspiformes such asQingshuiaspisjunqingi,Jiangxialepis jiujiangensis,andAnjiaspisericius(Shan et al.,2022a,2023a),the materials of Xiushuiaspidae are still awaiting further investigation.Here,we describe a new species of Xiushuiaspidae,Xiyuichthyslixiensissp.nov.,from the Qingshui Formation in Jiujiang of Jiangxi Province,and some new materials ofX.zhangifrom the Tataertag Formation in Kalpin of Xinjiang.The new findings not only enrich the morphological and taxonomic diversity of Xiushuiaspidae during the early Telychian but also provide direct fossil evidence for the correlations of the Tataertag Formation in Tarim Block and the Silurian Lower Red Beds (LRBs) in South China Block.More significantly,they extend the palaeogeographical distribution ofXiyuichthysfrom the Tarim Block to the South China Block for the first time.
2 Geological setting
Xiyuichthyslixiensissp.nov.was collected from the upper part of the Qingshui Formation near Lixi Town,Wuning County,Jiujiang City,Jiangxi Province (Fig.1A).Silurian strata outcropping in the northwestern Jiangxi form a marine depositional sequence mainly comprised of sandstones,mudstones,and shales.This sequence has been subdivided into six lithostratigraphic units in ascending chronological order: Lishuwo,Dianbei,Qingshui,Xiajiaqiao,Xikeng,and Xiaoxi formations (Liu,1997;Wang et al.,2018;Rong et al.,2019).The Qingshui Formation at Lixi Town mainly consists of purple and yellow-green siltstones,argillaceous siltstones,and mudstones intercalated with sandstones,silty mudstones,and shales(Shan et al.,2023a),from the upper part of which abundant fish fossils have been reported including galeaspidsQingshuiaspis junqingi,Jiangxialepis jiujiangensis,Anjiaspis ericius,some undescribed species of Dayongaspidae,Xiushuiaspidae,and Gumuaspidae,as well as chondrichthyansNeosinacanthus(Shan et al.,2022a,b,2023a).These fish fossils are grouped into the Wentang Assemblage which is widely distributed in the LRBs of South China,such as the Rongxi Formation in northwestern Hunan,the Tangchiawu Formation in northwestern Zhejiang,the Qingshui Formation in southeastern Hubei,and the Houjiatang Formation in eastern Anhui (Zhao and Zhu,2010,2014;Shan et al.,2022b,2023b).

Fig.1 Lithological columns of the Silurian fish-bearing strata in Wuning and Kalpin
New materials ofXiyuichthyszhangiwere collected from the Tataertag Formation near the Kermaiti of Kalpin County,Aksu City,Xinjiang (Fig.1B).Silurian strata distributed in the Kalpin area are composed of a continuous depositional sequence with about 2500 m in thickness.The strata are dominated by marine red beds and subdivided into the Kalpintag,Tataertag,Ymogantau,and Kezirtag formations in ascending order (Wang et al.,1996;Zhang et al.,1996;Zhao,2005;Zhao et al.,2009).The Tataertag Formation is composed of gray,grayish-white siltstone,sandstone,and mudstone intercalated with light-purple siltstone,with ripple marks in the siltstone,reflecting shallow-water littoral facies (Li et al.,1997).Fish fossils known from the Tataertag Formation include the galeaspidsNanjiangaspiskalpinensis,Kalpinolepistarimensis,Xiyuichthyszhangi,Microphymaspis pani,and chondrichthyansNeosinacanthusplanispinatus,Neosinacanthussp.,Sinacanthus wuchangensis,Tarimacanthus bachuensis,Tielikewatielepis sinensis,andXiaohaizilepis liui(Wang P et al.,1988;Wang J Q et al.,1996,2002;Zhu,1998;Lu et al.,2007;Gai et al.,2018;Liu et al.,2019;Andreev et al.,2020).These fish remains are grouped into the Tataertag Assemblage which is considered to be coeval with the early Telychian Wentang Assemblage from South China (Zhao et al.,2009;Zhao and Zhu,2010,2014;Shan et al.,2022b).
3 Materials and methods
The new materials include four incomplete headshields (IVPP V26119.1-V26119.4) ofXiyuichthyslixiensissp.nov.from the Qingshui Formation in Jiujiang,Jiangxi Province,and one incomplete headshield (IVPP V25445.4a,b) ofX.zhangifrom the Tataertag Formation in Tarim Basin.All specimens are permanently housed in the collections of the Institute of Vertebrate Paleontology and Paleoanthropology,Chinese Academy of Sciences (IVPP),Beijing,and accessible for examination.All specimens were prepared mechanically using a vibro tool with a tungsten-carbide bit or a needle,measured with a digital vernier calliper,and studied under an Olympus SZ61 zoom stereo microscope.All specimens were photographed with a Canon EOS 5D Mark III camera with a Canon EF 100 mm 1:2.8 L macro photo lens for the general morphology as well as a Canon MP-E 65 mm 1:2.8 1-5× macro photo lens and an Olympus SZ61 zoom stereo microscope for close-up images of the ornamentation.
4 Systematic paleontology
Subclass Galeaspida Tarlo,1967
Family Xiushuiaspidae Pan &Wang,1983
Genus Xiyuichthys (Liu et al.,2019)
NomenclatureXiyuichthysforXiyuaspis.
Recently Dr.Zhong Rui-Wen from the China University of Geosciences (Wuhan) gave us an important reminder about the synonym between the generic namesXiyuaspisin Galeaspida and in Trilobita.It is true thatXiyuaspisLiu et al.,2019,a Silurian galeaspid fish,is a junior synonym ofXiyuaspisZhang,1985,an Ordovician trilobite.Therefore,we propose the generic nameXiyuichthysto replaceXiyuaspisLiu et al.,2019.
Type speciesXiyuichthyszhangi(Lu et al.,2007).
Diagnosis(emended) Large-sized xiushuiaspid fishes;headshield subelliptic;95-125 mm in maximum width;maximum length about equal to maximum width;widest part positioned at about anterior 3/5 of headshield;median dorsal opening close to rostral margin of headshield,transversely oval in outline,width/length ratio 2-4;orbits small,dorsally positioned,distance between two orbits about equal to the distance from each orbit to lateral margin of headshield;lateral dorsal sensory canals developed,with anterior end almost reaching rostral margin of headshield;more than seven pairs of lateral transverse canals,and two pairs of median transverse canals;median dorsal canal absent;ornaments consisting of small or coarser round tubercles;ventral rim of headshield broadening posteriorly,approximately 19.0 mm in maximum width on each side;ventral rim of either side showing six successive notches for branchial openings,embracing a large round-shaped oralobranchial fenestra.
RemarksThe emendedXiyuichthysincludes two species,X.zhangifrom the Tataertag Formation in Tarim Basin andX.lixiensissp.nov.from the Qingshui Formation in South China.Xiyuichthysdiffers from the other two genera of Xiushuiaspidae,Xiushuiaspis(Pan and Wang,1983) andChangxingaspis(Wang,1991),in the aspects of individual size and the length/width ratio of the headshield.Xiyuichthyshas a large headshield with more than 100 mm in both length and width,which is much larger than the headshield ofXiushuiaspiswith a maximum length of 26.0 mm and a maximum width of 20.0 mm,as well asChangxingaspiswith a maximum length of 35.0 mm and a maximum width of 32.0 mm.In addition,the length/width ratio of the headshield is equal to or less than 1 inXiyuichthys,by contrast,that ratio is larger than 1 both inXiushuiaspisandChangxingaspis.Xiyuichthysresembles moreChangxingaspisthanXiushuiaspisby its comparatively broader and longer inner cornual processes.
Xiyuichthys lixiensis sp.nov.
(Figs.2-4)
Etymologylixi,from Lixi Town,Wuning County,in reference to the fossil site.
DiagnosisA large-sized xiushuiaspid fish,oval headshield with the maximum width of 123.4 mm,maximum width equal to or slightly larger than maximum length;transversely oval median dorsal opening with width/length ratio about 4;serrated lateral margin in the middle portion of the headshield;ornamentation consisting of coarser granular tubercles with only 1 tubercle per square millimetre.
RemarksXiyuichthyslixiensisis similar toX.zhangiin oval-shaped headshield,transversely oval median dorsal opening,cornual process absent,broad leaf-shaped inner cornual process,and the distribution pattern of the sensory canal system.However,X.lixiensisis distinguished fromX.zhangiby the ornamentation of the headshield,the long axis/short axis ratio of median dorsal opening,and the partially serrated lateral margin.The headshield ornamentations both inX.lixiensisandX.zhangiare composed of granular tubercles.However,the tubercles are extremely coarse with only 1 tubercle per square millimetre inX.lixiensis,whereas they are relatively small with 5-6 tubercles per square millimetre inX.zhangi.In addition,the long axis to short axis ratio of the median dorsal opening is about 4 inX.lixiensis,but the ratio is only 2 inX.zhangi.Therefore,a new species ofXiyuichthyscan be erected.
HolotypeAn incomplete headshield and its external mould,IVPP V26119.1a,b.
ParatypeAn incomplete headshield and its external mould,V26119.2a,b.
Referred specimensAn incomplete headshield,V26119.3,and an incomplete ventral rim,V26119.4.
Locality and horizonLixi Town,Wuning County,Jiujiang City,Jiangxi Province,China;Qingshui Formation,lower Telychian,Llandovery,Silurian.
MeasurementsSee Table 1.
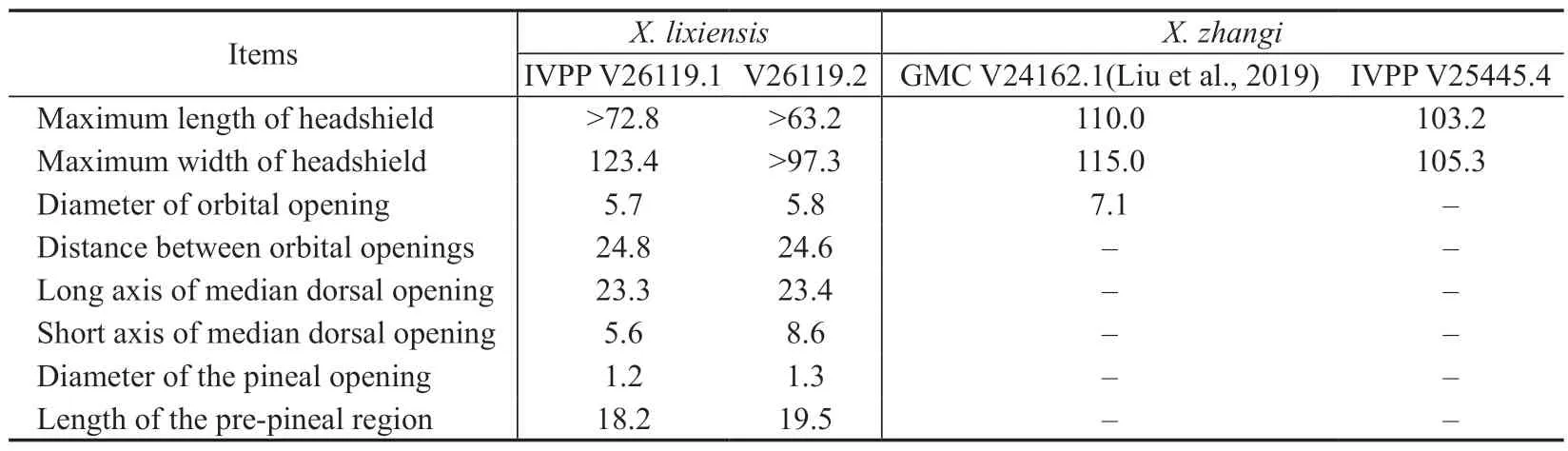
Table 1 Measurements and comparisons of Xiyuichthys lixiensis sp.nov.and X.zhangi (mm)
DescriptionBoth the holotype and paratype ofX.lixiensis(IVPP V26119.1,2) are incomplete headshields with their posterior parts missing (Fig.2A-D).The general outline of the headshield can be confirmed to be oval from specimen V26119.3 (Fig.3A) which shows clear lateral margin of the headshield.The preserved headshield of the holotype is 123.4 mm wide and 72.3 mm long,whereas that of the paratype is 97.3 mm wide and 63.2 mm long (Table 1).Reconstruction of the headshield indicates that the maximum length is likely 120.0 mm (94.6 mm long in specimen V26119.3).The rostral margin of the headshield is arciform in outline without a rostral process.The posterior margin of the headshield protrudes caudally into a pair of inner cornual processes (ic,Figs.3A;4A,B) which can be seen in specimen V26119.3 (Fig.3A).The inner cornual processes project posteriorly and are broad leaf-shaped with a length at 15.4 mm measured from their bases to tips,occupying 13% of the estimated maximum headshield length.

Fig.3 Photographs of Xiyuichthys lixiensis sp.nov.in ventral view
The median dorsal opening (md.o,Fig.2A-D) is dorsally situated,close to the rostral margin of the headshield,and transversely oval in outline with its short axis aligned with the rostro-caudal axis of the headshield.In the holotype,the long axis of the median dorsal opening measures 23.3 mm in length which is 4.2 times the length (5.6 mm) of the short axis.However,the long axis is only 2.7 times the short axis in length in paratype (V26119.2) whose long axis is 23.4 mm and the short axis is 8.6 mm.It is noteworthy that the median dorsal opening in the paratype shows an irregular outline probably deformed during the preservation process,therefore,we consider the holotype displays the natural outline of the median dorsal opening ofX.lixiensis.
The orbital openings (orb,Fig.2A-D) are dorsally positioned,posterior to the median dorsal opening,and round in outline with a diameter of about 5.7 mm (Table 1).The distance from the anterior margin of orbital openings to the posterior margin of median dorsal opening is about 4.0 mm.The distance between two orbital openings is about 24.6 mm which nearly equals to the long axis length of median dorsal opening (Table 1).
The pineal opening (pi,Fig.2A-E) is situated in the midline of the headshield and level with the posterior margin of orbital openings.It is tiny and round in outline with a diameter of 1.2 mm.The pre-pineal region of the headshield is short with a length of about 19.0 mm,constituting approximately 16% of the estimated maximum length of the headshield.
The sensory canal system is well-preserved in the paratype (V26119.2,Fig.2C,D) and partly preserved in the holotype (V26119.1,Fig.2A,B).The identified sensory canals consist of infraorbital canals (ifc),lateral dorsal canals (ldc),lateral transverse canals (ltc),and median transverse canals (mtc).The infraorbital canals (ifc,Figs.2C;4A) are lateral to the orbital opening with anterior end almost reaching the rostral margin of the headshield.Posteriorly,the infraorbital canals connect with the lateral dorsal canals (ldc,Figs.2C,D;4A) which curve inward at the middle of the headshield and then extend towards the posterior margin of the headshield.There are at least seven pairs of lateral transverse canals (ltc,Figs.2BD;4A) extending posterolaterally.In addition to five pairs of lateral transverse canals (ltc1-5)running from the lateral dorsal canals,two pairs of lateral transverse canals (ltca,b,Figs.2BD;4A) are observed issuing from the infraorbital canals.The left and right lateral dorsal canals are connected by two median transverse canals (mtc1-2,Figs.2C,D;4A),which are in level with the second and third lateral transverse canals (ltc2-3),respectively.The anterior median transverse canal is equivalent to the dorsal commissure of other galeaspids.
The lateral margin of the middle portion of the headshield is serrated,but the anterior and posterior portions of the headshield is smooth (Figs.2A,B;3A;4A).The dorsal surface of the headshield is ornamented with closely-set and extremely coarse granular tubercles with only one tubercle per square millimetre (Fig.2A,B).
The ventral part of the headshield is known from a poorly preserved ventral rim (V26119.4,Fig.3B).The ventral rim is narrow anteriorly,broadening gradually towards the inner cornual processes of the headshield.Several semicircular notches along the mesial margin of the ventral rim may display the positions of branchial openings (br.o,Figs.3B,4B).The number of branchial openings is six in basal galeaspids such asDayongaspis(Pan and Zeng,1985),Hanyangaspis(P’an et al.,1975),Xiushuiaspis(Pan and Wang,1983),andChangxingaspis(Wang,1991).Therefore,we reconstructed six branchial openings forX.lixiensis(Fig.4B).
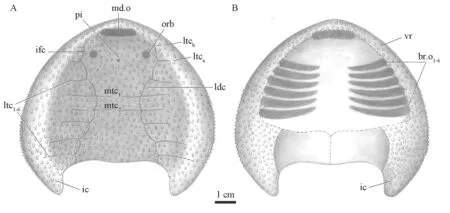
Fig.4 Restoration of Xiyuichthys lixiensis sp.nov.in dorsal (A) and ventral (B) views For abbreviations see Figs.2,3
Xiyuichthys zhangi (Lu et al.,2007)
(Figs.5,6)
NanjiangaspiszhangiLu et al.,2007
KalpinolepiszhangiLiu et al.,2015
XiyuaspiszhangiLiu et al.,2019
Diagnosis (emended)A large-sized xiushuiaspid fish with the headshield about 110 mm in maximum length,maximum width roughly equal to length;transversely oval median dorsal opening with width/length ratio of about 2;lateral margin of the headshield serrated;ornaments consisting of small granular tubercles with six tubercles per square millimetre;oralobranchial fenestra round-shaped.
HolotypeAn incomplete headshield and its external mould,GMC V2191a,b.
ParatypeAn incomplete headshield,GMC V2192.
Referred specimenAn incomplete headshield,IVPP V25445.4a,and its external mould V25445.4b.
Locality and horizonKalmaiti,Kalpin County,Xinjiang Uygur Autonomous Region,China;Tataertag Formation,Telychian,Llandovery,Silurian.
MeasurementsSee Table 1.
DescriptionThe new specimen ofXiyuichthyszhangiis an incomplete headshield(IVPP V25445.4a,Fig.5A) with a nearly complete ventral rim (vr,Fig.5C,D),and its external mould (V25445.4b,Fig.5B).The headshield is oval in outline with a maximum length of 103.2 mm and maximum width of 105.3 mm.As in the holotype ofX.zhangi,the rostral margin of the headshield in the new specimen is round and blunt,and the lateral margin of the headshield is deeply arciform,reaching the widest point at the anterior 3/5 of the headshield.It is noteworthy that the lateral margin of the headshield in the new specimen is serrated (Fig.5A,B),therefore,the smooth lateral margin in the holotype ofX.zhangimay have been caused by the preservation.The median dorsal opening and orbital openings are hard to distinguish in the new specimen.

Fig.5 Photographs and interpretive drawing of Xiyuichthys zhangi
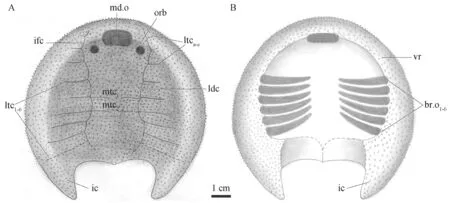
Fig.6 Restoration of Xiyuichthys zhangi in dorsal (A) and ventral (B) views For abbreviations see Figs.2,3
The sensory canal system is poorly preserved in the new specimen with only lateral dorsal canals (ldc),infraorbital canals (ifc),seven pairs of lateral transverse canals (ltca-cand ltc2-5),and two median transverse canals (mtc1-2) identified (Fig.5B).The distribution pattern of sensory canals in the new specimen is also the same as that in the holotype with a pair of lateral dorsal canals (ldc) and infraorbital canals (ifc) constituting the main longitudinal canals and eight-nine pairs of lateral transverse canals (ltc) and two medial transverse canals (mtc)issuing from them.
Like the holotype ofX.zhangi,the new specimen is ornamented with small dense and evenly distributed granular tubercles on the headshield with about six tubercles per square millimetre.After removing the dorsal headshield of V25445.4a,a nearly complete ventral rim is exposed (Fig.5C),which supplements the missing ventral information of the headshield in the holotype.The ventral rim broadens gradually posteriorly and encloses a nearly roundshaped oralobranchial fenestra in the center,showing the position of the oralobranchial chamber (obr.c,Fig.5C,D).The ventral rim expands posterolaterally to form the bases of the inner cornual processes (Fig.5C,D) where the ventral rim attains the largest width of 18.6 mm.Several semicircular pores along the mesial margins of the ventral rims may display the position of the branchial openings (br.o,Figs.5C;6B).
5 Discussion and conclusion
The discovery ofXiyuichthyslixiensissp.nov.from the Qingshui Formation at Wuning,Jiangxi Province extends the paleogeographical distribution ofXiyuichthysfrom the Tarim Block to the South China Block.The Silurian fish-bearing strata in northwestern Jiangxi include the Qingshui Formation and the Xikeng Formation (Fig.7),which are known as the Silurian Lower Red Beds (LRBs) and Upper Reds Beds (URBs),respectively (Rong et al.,2019).Fish fossils in the Qingshui Formation (LRBs) are characterized by the co-occurrence of two plesiomorphic clades (Dayongaspidae and Xiushuiaspidae),the most basal clade of Eugaleaspiforme (Shuyuidae),the most basal clade of Polybranchiaspiforme (Gumuaspidae),and diversified sinacanths (Shan et al.,2022a,b,2023a).These fish remains have been grouped into the Wentang Assemblage which represents the oldest vertebrate assemblage in the Silurian of China (Zhao and Zhu,2010,2014;Shan et al.,2022b).Fish remains in the Xikeng Formation (URBs) have been grouped into the Maoshan Assemblage which is characterized by the co-occurrence of Xiushuiaspidae and Sinogaleaspidae (Pan and Wang,1980,1983;Gai et al.,2020;Shan et al.,2020),the latter of which is a more derived clade of Eugaleaspiformes than Shuyuidae (Shan et al.,2020).

Fig.7 Biostratigraphic sequence of Telychian (Llandovery,Silurian) brachiopods,trilobites,bivalves,gastropods,and fishes in northwestern Jiangxi
The age of the Qingshui Formation and the Xikeng Formation can be well constrained to be the early Telychian and middle-late Telychian respectively,because they clip the Xiajiaqiao Formation in the middle (Zhao and Zhu,2014;Wang et al.,2018),which yields the middle Telychian Xiushan Fauna comprising trilobitesCoronocephalus,Kailia,Rongxiella,Senticuculus,andXiushuiproetus;brachiopods Striispirifer;bivalvesOrthonata;and gastropodsHormotoma(Fig.7) (Zhang,1982;Chen and Rong,1996;Wang et al.,2018).Therefore,the lower Silurian in northwestern Jiangxi can provide a standard section (Fig.7)for the correlations of Silurian shallow marine red beds in South China and Tarim blocks.Especially,some new species of Eugaleaspiformes from northwestern Jiangxi have made more precise correlations by refining the taxonomy from a generic level to a specific level.For example,the finding ofAnjiaspis ericiussuggests that the Tangchiawu Formation in northwestern Zhejiang should be correlated to the LRBs (the Qingshui Formation) rather than URBs (the Xikeng Formation) because it is comparable toA.reticularisdirectly on the specific level.In addition,the finding ofJiangxialepis jiujiangensisfrom the Qingshui Formation in northwestern Jiangxi indicates that the fish-bearing strata in Wuhan of Hubei are most likely to be the early Telychian rather than the middle Telychian as previously assumed because it is comparable toJ.retrospinadirectly on the specific level as well (Shan et al.,2023a,b).
Vertebrate fossils in the Tataertag Formation in the Tarim Basin are referred to as the Tataertag Assemblage (Zhao and Zhu,2014).The Tataertag Assemblage can compare to the Wentang Assemblage in South China in general,e.g.Microphymaspiscan compare toDayongaspisfrom the Rongxi Formation in Zhangjiajie of Hunan (Pan and Zeng,1985);KalpinolepisandNanjiangaspiscan compare toHanyangaspisfrom the Houchiatang Formation in Wuhan of Hubei andLatirostraspisfrom the Houchiatang Formation in Chaohu of Anhui (P’an et al.,1975;Wang et al.,1980);Platylomaspisis comparable to the new materials of Gumuaspidae collected from the Houjiatang Formation in Chaohu of Anhui and the Qingshui Formation in Jiujiang of Jiangxi (Shan et al.,2022b).However,all these comparisons still rest on evidence on the generic level (belonging to the same family).The findings ofXiyuichthyslixiensissp.nov.from the Qingshui Formation andX.zhangifrom the Tataertag Formation provide direct evidence on the specific level for the correlation between these two formations (Fig.8).
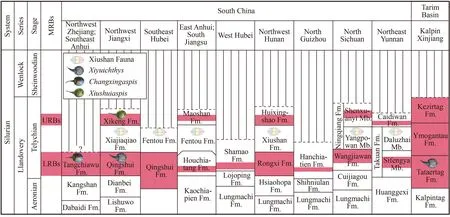
Fig.8 Stratigraphical positions and correlations of the Silurian marine red beds in South China and Tarim Basin
The fossil records indicate that Xiushuiaspidae has a broader stratigraphic range than other families of the Wentang Assemblage,which extends from the LRBs Qingshui Formation upward into the URBs Xikeng Formation in northwestern Jiangxi.However,Xiushuiaspidae shows a distinct composition in the LRBs and URBs on the specific level.Specifically,two species ofXiushuiaspisco-occurred in the Xikeng Formation (URBs) in northwestern Jiangxi (Pan and Wang,1983),whereas two species ofChangxingaspisare known from the Tangchiawu Formation (LRBs) in northwestern Zhejiang (Wang,1991) and the Tataertag Formation in Tarim Basin (to be described in another paper),and two species ofXiyuichthysare known from the Qingshui Formation in northwestern Jiangxi and the Tataertag Formation in Tarim Basin,all of which belong to the LRBs (Figs.7,8).
AcknowledgementsWe are grateful to Prof.Rong Jiayu for improving Fig.7,Zhao Ridong,Meng Xinyuan,Shen Yiming,Liu Yuhang,and Liu Wenyu for their great help in the fieldwork,Yang Dinghua and Gui Fang for the illustrations and ecological restoration.

- 古脊椎动物学报(中英文)的其它文章
- Reappraisal of Bothriolepis sinensis Chi,1940 from the Tiaomachien Formation,Hunan,China
- A giant bamboo rat from the latest Miocene of Yunnan
- Micromammal fossils from the basal part of the Jiaozigou Formation in Yagou area,Linxia Basin,Gansu Province
- An egg clutch of the Stalicoolithidae discovered in Wuning,Jiangxi,China