
濒危植物闽楠幼苗更新及环境解释
2023-12-05 03:40姜冬冬林建勇罗应华
西北植物学报 2023年11期
姜冬冬,林建勇,李 娟,罗应华
(1 广西壮族自治区林业科学研究院,国家林业局中南速生材繁育实验室,南宁 530002;2 广西大学 林学院,南宁 530004)
植物种群更新是森林自我繁殖和演替的重要途径,其幼苗期是植物生活史中生长最脆弱、对外界环境反映最敏感的阶段[1],当幼苗成功转成幼树,表明它成功逃离了整个生活史最危险的阶段[2]。种子从母树上产生、扩散、萌发、定植,进而成为幼苗进入种群,期间会受到3种更新限制[3-6]:一是种源限制,种子数量少,使得有效传播率低;二是扩散限制,种子不能扩散到适合萌发地点,导致种子萌发率低;三是微环境限制,种子成功扩散到微生境后,由于群落中剩余生态位有限,且受微环境的生物和非生物因素影响而不能萌发形成幼苗或幼苗不能生长成幼树,仅有少量种子能建成幼树完成更新。种源限制决定了种子库的数量,扩散限制决定了种群的分布,微环境限制决定了种子萌发和幼苗建成的成功率[5-7]。幼树的成功建成是决定种群命运的重要因素,是植物种群维持和更新的重要阶段[8]。影响种群更新的微环境因子主要有土壤因子(土壤温度、湿度、盐分)、生物因子(林分郁闭度、灌草层覆盖、凋落物覆盖、物种竞争等)、光因子等。植物种群更新证实了幼树更新成功率远低于母树繁殖投入成本。从小尺度而言,微生境限制是幼苗建成和生长的重要影响因子。
闽楠(Phoebebournei)是中国特有的国家二级重点保护植物,是中国暖温带和亚热带地区珍贵用材“金丝楠木”中商品材量最大、分布范围最广、生长最快的一种[9-10]。闽楠主要分布于广西北部和西北部、贵州东南部、湖南、江西、福建西北部,浙江南部、广东北部、湖北南部有少量分布。闽楠材质优良、木纹美观、价值高,但长期的人为破坏导致野生资源日益减少,已处于濒危状态。富川县是广西闽楠野生资源分布面积最大和数量最多的区域,分别占广西闽楠总量的38.33%和66.73%[9],其中蚌贝村种群个体密度高达80 249株/hm2[10],是全国个体密度最大的种群。在前期研究中笔者发现,富川县白面寨闽楠天然林在以采集种实为主的非破坏性干扰下,闽楠更新个体向高龄级个体补充的成功率更高,种群的生存率也有所提高,而在相距仅1.1 km无干扰的蚌贝村闽楠天然林中,幼苗密度很大,但幼苗死亡率高,种群出现衰退趋势[10]。
研究表明,闽楠幼苗更新与凋落物厚度、林下植被盖度、坡度呈负相关,而与土壤养分呈正相关[11-12],遮荫会降低闽楠幼苗光合效率和生物量积累[13]。植物天然更新状况是多种影响因子共同作用的结果,各种微环境因子是如何影响闽楠天然更新的?环境因子之间的关系如何?决定种子萌发和幼苗存活的微环境因子对闽楠天然更新的影响有多大?这些问题仍不够清楚。基于此,本研究以未受人为干扰的富川县蚌贝村闽楠种群为对象,通过分析闽楠幼苗种群结构与环境因子的关系,掌握闽楠幼苗定居的影响因子,探讨各种环境因子对闽楠种群幼苗更新过程的相对影响,并找出影响种群更新的关键限制因子,为促进闽楠天然更新和野生资源保护提供科学依据。
1 材料和方法
1.1 研究地概况
研究区域位于广西东北部的富川县(111°5′—111°29′E,24°37′—25°9′N),样地附近地貌属溶蚀平原,气候属亚热带季风气候,年平均气温19 ℃,年均降雨量1 700 mm,降雨集中于夏季,年相对湿度75%。富川县朝东镇蚌贝村拥有广西面积最大、个体集中分布的闽楠天然林。群落优势种除闽楠外,还有苦槠(Castanopsissclerophylla)、木荷(Schimasuperba)、枫香树(Liquidambarformosana)。
1.2 野外调查
1.2.1 样地设置与调查
蚌贝村闽楠天然林为风水林,面积2.1 hm2,四周农田环绕,无人为破坏。海拔290 m,地势平坦无坡向,土壤类型为红壤。避开林缘部分,于群落中设置80 m×80 m固定监测样地。样地划分为10 m×10 m的乔木层调查样方,每个样方东北角再划分出1个5 m×5 m的更新层调查小样方。小样方内按划分的高度级统计胸径5.0 cm以下个体的数量,共调查80个5 m×5 m样方。
1.2.2 闽楠更新个体年龄结构的划分与调查
在树种年龄结构研究中,常采用以大小级代替年龄级的“空间代替时间”方法,本研究根据闽楠生长特性,将幼苗幼树根据高度(H)分为4级:其中Ⅰ级(小幼苗),H<10 cm;Ⅱ级(中幼苗),10 cm≤H<50 cm;Ⅲ级(大幼苗),50 cm≤H<100 cm;Ⅳ级(幼树),H≥100 cm,DBH<5.0 cm。记录更新层调查小样方内闽楠各级幼苗幼树的数量。
1.2.3 林内微环境因子的调查
在各小样方内用TES-1332A照度计测定林内光照,LAI-2200冠层分析仪记录每个样方的叶面积指数(LAI),WET土壤三参数速测仪测定土壤水分、温度、电导率,以上数据每个样方测量5次,分别在每个样方的中心和四角位置。为减少误差,以上指标测定于12:00—13:00的1 h内完成。此外,调查样方内的草本层覆盖度、灌木层覆盖度、凋落物覆盖度、草本层高度、灌木层高度、凋落物厚度及样方中心与最近母树的距离等指标。
1.3 数据处理
采用Excel进行数据初步整理,采用R语言中的corrplot包对群落微环境因子之间、幼苗密度与微环境因子之间的关系进行Pearson相关系数分析,用vegan包对幼苗密度与微环境因子之间的关系进行冗余分析(RDA)并用蒙特卡罗置换检验对RDA结果进行显著性分析,用ggplot2包绘图。
2 结果与分析
2.1 闽楠更新幼苗高度级和龄级结构
闽楠更新层个体共调查到12 834株,个体密度80 212.5株/hm2。由图1可以看出,闽楠更新层种群高度级和龄级结构均呈典型的“J”形。小高度级个体占有较大比重,尤其是H<0.1 m的小幼苗,占总数比例高达81.43%,H≥0.3 m之后的幼苗数量变化较稳定,大幼苗和幼树比例分别仅占2.07%和3.36%。幼苗死亡率最高时出现在H<0.1 m高度级至0.1~0.2 m高度级的转化阶段,死亡率达到89.63%。

图1 闽楠更新层种群高度级、龄级结构Fig.1 The age structure of P. bournei population in update layer
说明,闽楠种群能在较长时间内保持稳定,小幼苗库的建成不存在种源限制,但幼苗死亡率高,低龄级个体向高龄级的转化成功率低,尤其是在小幼苗阶段更为明显,闽楠种群存在严重的幼苗更新限制。
2.2 微环境因子之间的相关性
对林下微环境各因子之间的相关性进行分析,结果见图2。光照强度与土壤盐分含量、乔木层叶面积指数、灌木层覆盖度和高度、凋落物覆盖度和高度等环境因子均存在显著或极显著的负相关。土壤水分与土壤温度呈正相关,与土壤盐分含量和叶面积指数呈负相关。土壤温度与土壤盐分含量、乔木层叶面积指数、草本层覆盖度显著负相关。土壤盐分含量与乔木层叶面积指数显著正相关,与光照、土壤水分、土壤温度显著负相关。群落微环境因子之间关系复杂,并且有着不一样的关联性,说明林下微环境因子对林下幼苗更新并非由单一因子决定而是多因子综合作用的结果。
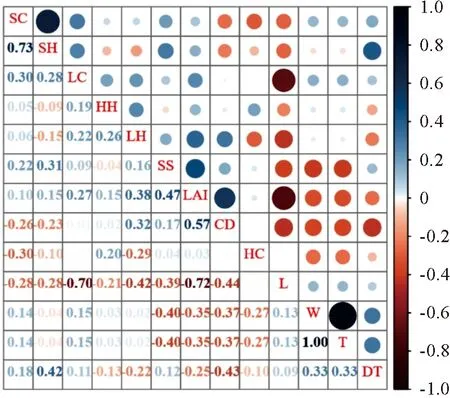
L. 光照; W. 土壤水分; T. 土壤温度; SS. 土壤盐分含量;LAI. 叶面积指数; CD. 林冠层郁闭度; DT. 与最近母树距离;HC. 草本层覆盖度; SC. 灌木层覆盖度; LC. 凋落物覆盖度;HH. 草本层高度; SH. 灌木层高度; LH. 凋落物厚度。0~0.1无相关,0.1~0.24弱相关,0.25~0.30显著相关,0.31~1.0极显著相关。下同。图2 环境因子之间的相关性分析L. Light; W. Soil water; T. Soil temperature; SS. Soil salt content;LAI. Leaf area index; CD. Canopy closure; DT. Distance from the nearest parent tree; HC. Herb coverage; SC. Shrub coverage;LC. Litter coverage; HH. Herb height; SH. Shrub height;LH. Litter thickness. 0-0.1 no correlation, 0.1-0.24 weak correlation, 0.25-0.30 significant correlation, 0.31-1.0 extremely significant correlation. The same below.Fig.2 Correlation between environmental factors
2.3 更新个体与环境因子的相关性
将各样方中的闽楠更新个体密度与微环境因子进行相关分析,结果如图3。小幼苗密度与光照、草本层覆盖度呈负相关,与乔木层叶面积指数、凋落物厚度呈正相关;中幼苗与草本层覆盖度呈负相关,与灌木层覆盖度、灌木层高度存在正相关;大幼苗和幼树与微环境因子的关系相似,均与光照呈负相关,而与土壤盐分含量、叶面积指数、与母树距离、灌木层盖度、灌木层高度呈正相关。闽楠不同龄级间的个体密度呈正相关关系,大幼苗的更新个体数对幼树的更新最为显著。综合分析表明,闽楠幼苗更新的限制因子有光照和草本层盖度;乔木层叶面积指数、土壤养分含量、灌木层盖度、灌木层高度、凋落物厚度则是闽楠幼苗更新的促进因子;土壤水分、土壤温度、林冠层郁闭度、凋落物覆盖度、草本层高度等因子与幼苗密度不相关,不是闽楠幼苗更新的影响因子。大部分微环境因子对不同龄级更新个体的影响相同,或在部分龄级无影响,未在不同龄级间出现明显相反作用。

xym. 小幼苗; zym. 中幼苗; dym. 大幼苗; ys. 幼树; zt. 更新层整体。下同。0~0.1无相关,0.1~0.24弱相关,0.25~0.30显著相关,0.31~1.0极显著相关。图3 更新苗密度与微环境因子相关性xym. Small seedlings; zym. Medium seedlings; dym. Large seedlings; ys. Sapling; zt. Whole regeneration. The same below. 0-0.1 no correlation, 0.1-0.24 weak correlation, 0.25-0.30 significant correlation, 0.31-1.0 extremely significant correlation.Fig.3 Correlation between density of regenerated seedlings and microenvironmental factors
2.4 冗余分析
闽楠幼苗更新是多种微环境因子共同作用的结果,直观表现闽楠更新过程中所受微环境的影响,对不同龄级更新个体与微环境因子进行RDA排序,环境因子对闽楠幼苗更新的解释量为85.91%;从图4、表1可以看出,RDA前两轴均反映了多个环境因子的综合影响,RDA第一轴与土壤盐分含量、灌木层覆盖度、灌木层高度、凋落物覆盖度、叶面积指数显著正相关,与草本层覆盖度、草本层高度、光照负相关;RDA第二轴与林冠层郁闭度、凋落物厚度正相关,与土壤水分、土壤温度负相关。
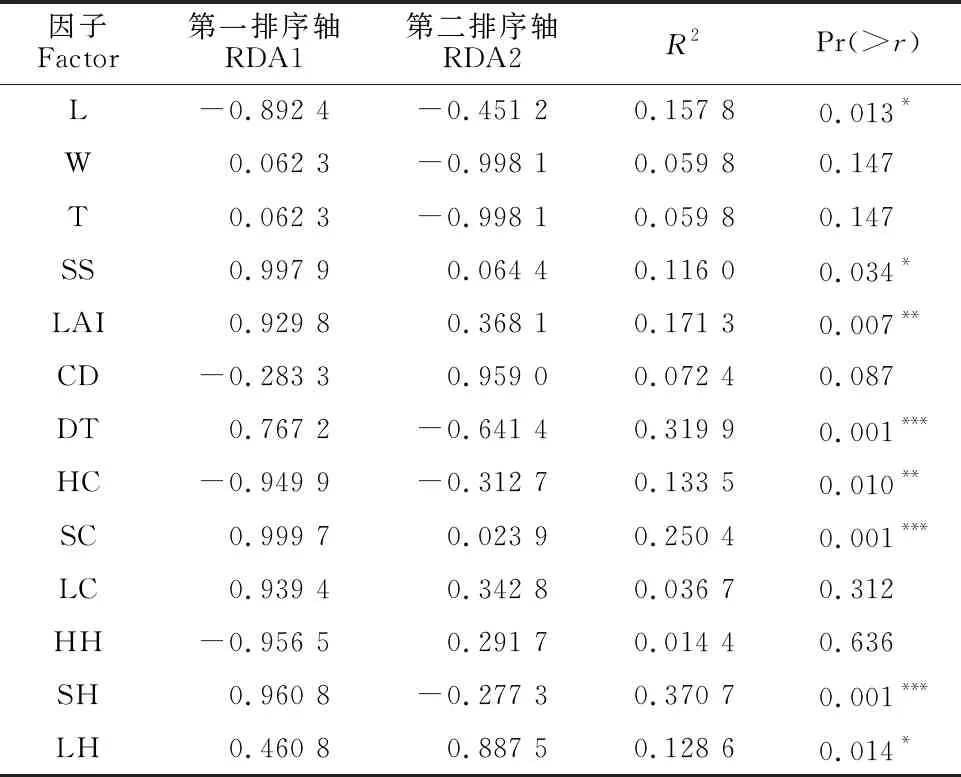
表1 更新苗密度与微环境因子的蒙特卡罗检验

图4 更新苗密度与微环境因子冗余分析Fig.4 Redundancy analysis of regeneration seedling density and microenvironment factors
结合RDA排序图与微环境因子的蒙特卡罗检验分析可知,灌木层高度对闽楠幼苗更新影响最大,其次为与母树的距离、灌木层覆盖度,再者是叶面积指数、光照、草本层覆盖度、凋落物厚度、土壤盐分,这些都是决定闽楠幼苗更新的主导因子,其中限制因子是光照和草本层覆盖度;而较高水平的土壤盐分、凋落物覆盖度、叶面积指数、灌木层覆盖度、灌木层高度、与母树距离对幼苗更新有促进作用。
3 讨 论
母树对幼苗更新的影响一直是种群生态学研究的热点,土壤种子库是植物种群天然更新的基础[14],种源限制是种群更新需要克服的第一步。本研究中闽楠更新层种群高度级和龄级均呈典型的“J”形,小幼苗的密度很高,不存在种源限制,种群稳定,但小幼苗死亡率高。从更新个体整体上看,母树并未产生明显影响,但在不同龄级中的影响差异较大,小幼苗与母树距离之间存在弱负相关,且相关性已接近显著水平,随着龄级变大,更新个体密度与母树距离的相关性转变为正相关,且相关性逐渐变大。说明小幼苗密集分布于母树下,但母树下环境对小幼苗生长不利,远离母树的环境中更适宜大幼苗和幼树的生长,这可能是受密度制约效应[15-16]和种内竞争[17]的影响。
影响植物天然更新的环境因子可以归纳为光、水、土壤、林下植被、凋落物、地形等,环境因子之间关系复杂,有些因子并非直接影响植物天然更新,如坡向、坡位、坡度、海拔等地形因子,其作用是因为改变了环境中的光、水、土壤等环境因子[18-19],乔木层郁闭度、叶面积指数则是通过改变透光率而影响林下光照强度。众多研究[11,18,20]表明,环境因子对闽楠幼苗更新的影响是多因子综合作用的结果,本研究所选环境因子对闽楠幼苗更新的解释率为85.91%,限制因子主要有光照和草本层覆盖度。对闽楠天然更新有促进作用的环境因子有土壤养分、灌木层覆盖度和高度等,小幼苗的影响因子还有凋落物。
林下植被对幼苗更新有较大的影响,这多与密度制约效应和种内种间竞争有关。刘晓静等[21]在宝天曼落叶阔叶林中的研究表明,小径级个体的生存在较小范围内受密度制约的影响比中大径级个体更为明显,何中声等[22]认为竞争强度与幼苗的高度呈显著负相关,这与本研究中小幼苗密度大但死亡率高的结果相同。随着高度增加,幼苗受到的种内竞争强度越来越小,但种间竞争强度却逐渐增大[22]。徐来仙等[23]发现,林下环境对种群天然更新有显著影响,林下灌木、草本、凋落物均有阻碍作用。李雪云等[11]认为,闽楠天然更新与林下植被盖度负相关,与林下植被高度不相关,本研究中,草本层盖度呈负相关,但灌木层盖度为正相关,灌木层高度对中、大幼苗和幼树也有极显著相关,产生研究结果不同的原因可能是李雪云等[11]的研究中未把灌木层和草本层区别分析,这也说明了林下植被对幼苗更新的复杂性。为促进闽楠天然更新,应适当去除林下草本植物,以缓解草本植物对闽楠幼苗的限制作用,但要保留林下木本植物。在营造闽楠人工林的幼林阶段,也应控制草本植物生长。
光照环境对闽楠幼苗更新有较大影响,在森林的不同位置中,闽楠幼苗生长差异显著[24-25]。闽楠更新个体耐阴性强,较强的光照条件对闽楠幼苗更新有明显的抑制作用。在闽楠野生资源保育实践中,为促进其天然更新,应保证所在群落有较高的郁闭度,结合李雪云等[11]的研究,建议把林分郁闭度控制在0.5以上。
猜你喜欢
科学技术创新(2022年30期)2022-10-21
农业与技术(2021年23期)2021-12-14
山东林业科技(2021年4期)2021-08-31
原子与分子物理学报(2020年5期)2020-03-17
福建林业科技(2018年1期)2018-04-09
乡村科技(2018年8期)2018-02-13
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
浙江林业科技(2011年2期)2011-05-09
水土保持研究(2010年6期)2010-09-19
