
外源褪黑素对养分胁迫下苹果砧木幼苗氮代谢相关酶活性及基因表达的影响
2023-11-28 09:48刘怀特杜培华周莎莎李中勇张学英徐继忠梁博文
植物营养与肥料学报 2023年10期
刘怀特,曹 扬,杜培华,周莎莎,李中勇,张学英,徐继忠,梁博文
(河北农业大学园艺学院,河北保定 071001)
苹果是多年生经济作物,我国的苹果栽培面积和产量位居世界第一[1]。研究表明,植物需要从土壤中获取至少14 种必需的营养元素,以维持正常的生长发育,当土壤中的营养元素缺失时,植物会产生一系列的应答反应[2-3],比如土壤中氮素亏缺会导致植株生长缓慢,叶色变黄,甚至作物产量降低等现象[4]。为缓解营养缺乏而造成的缺素症状,农业上常采取施肥措施,但施肥不科学容易导致资源浪费、环境退化和生态环境破坏等问题[4]。因此,如何提升养分胁迫下苹果养分吸收利用效率,对减少人工合成肥料的施用与生态环境的保护具有重要意义。
氮是植物生长必需的大量元素,对植物器官分化、物质代谢及果实品质形成等过程具有不可替代性,参与苹果生长发育的各个阶段,在细胞发育和生命活动中发挥着至关重要的作用[5-6]。研究表明,铵态氮(NH4+)和硝态氮(NO3-)是植物体内主要的氮源,NO3-可通过植物根表皮和表皮细胞从土壤中吸收利用[7-8],对植物生长的促进作用显著[9],而NH4+可以更快地填充土壤溶液中的无机氮溶解池。AMT1和AMT2是植物中两类主要的NH4+转运蛋白,负责植物体内NH4+的转运与吸收[10],NRT1与NRT2家族参与植物器官对NO3-的吸收与转运[11]。
褪黑素是一种强抗氧化剂,能够抑制活性氧的积累,调控逆境响应基因的表达,对盐碱、重金属和干旱等环境胁迫具有很强的缓解作用[12]。研究表明,在养分胁迫和干旱胁迫下,施加褪黑素会促进氮、磷和钾的吸收与分配[13-14];低氮胁迫下,外源褪黑素可以促进小麦幼苗的生长,提高幼苗茎部和根部NO3-含量[15];低NO3-条件下,褪黑素可以促进叶片15NO3-积累,增加15NO3-含量和15NO3-吸收通量[16]。上述研究表明,褪黑素在单一元素亏缺或其他环境胁迫下能够促进植株生长及养分吸收。
虽然对单一元素亏缺或其他环境胁迫下苹果植株氮素吸收利用的研究很多,但是关于外源褪黑素对养分全缺情况下苹果植株氮代谢酶活性及基因表达的影响研究尚缺乏。因此,本试验以平邑甜茶实生苗为材料,在水培条件下模拟养分胁迫,从养分胁迫下平邑甜茶幼苗的氮代谢关键酶活性、氮代谢及氮转运相关基因的表达等方面,探究外源褪黑素对养分胁迫下苹果幼苗氮代谢和转运的影响,为外源褪黑素提升苹果砧木氮素吸收利用效率提供理论依据。
1 材料与方法
1.1 试验设计
本试验以平邑甜茶幼苗为试验材料,于河北农业大学创新实验园(38°23′ N,115°28′ E)中进行,将经低温层积处理30 天的平邑甜茶种子种于育苗钵中,待幼苗长至7~8 片叶时,将其转移至盛有1/2 Hoagland 营养液的水培盆中,通过气泵通气以保持植物根部正常生长,使用H3PO4调节营养液的pH为6.1±0.1,每5 天更换1 次营养液。
首先进行预培养,第12 天时,将材料分成两组,一组为对照组,另一组施加0.1 μmol/L 褪黑素,第15 天时,开始进行正式处理。将幼苗均匀分成4 个处理组,分别为1/2 Hoagland 营养液(CK)、1/2 Hoagland 营养液+0.1 μmol/L 褪黑素(MCK)、1/20 Hoagland 营养液(ST)、1/20 Hoagland 营养液+0.1 μmol/L 褪黑素(MST),每个处理3 次重复,每个重复40 株幼苗,共120 株幼苗,每5 天处理1 次,共处理20 天。
在每次试验处理之前(第0、5、10、15 天上午9:00) 和第20 天时,采集顶端(生长点) 以下第5~6 片健康、无破损的成熟叶片,同时在第20 天采集幼苗根系,液氮冷冻后置于-80℃冰箱保存,用于后续酶活性、基因定量表达和褪黑素含量的测定分析,每个指标进行3 次生物学重复。
1.2 氮代谢关键酶活性的测定
硝酸还原酶(NR)、亚硝酸还原酶(NiR)、谷氨酰胺合成酶(GS)、Fd-谷氨酸合成酶(Fd-GOGAT) 和NADH-谷氨酸合成酶(NADH-GOGAT)活性采用苏州科铭生物技术有限公司的试剂盒进行测定。
1.3 荧光定量PCR 检测
按照多糖多酚植物样本RNA 提取试剂盒(北京聚合美生物科技有限公司)的说明书从叶和根中提取总RNA,使用UEIriRT-PCR System for First-Strand cDNA Synthesis system (苏州宇恒生物科技有限公司)进行反转录得到cDNA,使用Roche LightCycler 96 real-time PCR system (Roche,Basel,Switzerland)分析基因的相对表达量,以β-Actin为内参基因,每个样品进行3 次重复,引物序列见表1。
1.4 褪黑素含量的测定
褪黑素含量的测定参照Cao 等[17]的方法。取0.5 g 样品于液氮中研磨成粉末,加入5 mL 甲醇,超声35 min,4℃,10000×g离心15 min,上清液过0.22 μm 有机滤膜后,使用高效液相色谱(HPLC)检测叶片中褪黑素含量。
1.5 数据处理与分析
使用Excel 2016 和IBM SPSS Statistics 20 进行数据处理与分析,采用SigmaPlot 10.0 绘制图表,使用单因素方差分析(ANOVA)比较统计学差异,Tukey’s多区间检验(P<0.05)评价处理间差异。
2 结果与分析
2.1 外源褪黑素对褪黑素含量及其相关基因表达的影响
如图1 所示,处理20 天中,CK 处理的褪黑素合成相关基因的表达相对稳定,MCK 处理的MdTDC、MdT5H和MdAANAT的表达量呈先上升后下降的趋势,在处理第10 天达到峰值,MdASMT的表达呈上升趋势,在处理第20 天值最高。ST 处理的MdTDC、MdT5H、MdAANAT和MdASMT的表达量呈先上升后下降的趋势,MST 处理的MdTDC和MdAANAT的表达量呈上升趋势,在处理第20 天值最高,MdT5H的表达量呈先下降再上升后下降的趋势,在处理第15 天达到峰值,MdASMT的表达量呈先上升再下降后上升的趋势,在处理第20 天值最高。处理第20 天时,与CK 处理相比,ST 处理上调了褪黑素合成相关基因MdTDC、MdT5H、MdAANAT和MdASMT的表达。与ST 处理相比,外源褪黑素提高了苹果幼苗叶中MdTDC、MdT5H、MdAANAT和MdASMT的表达水平。如图2 所示,养分胁迫下,苹果幼苗叶片中褪黑素含量显著增加,外源施加褪黑素显著增加了苹果幼苗叶中的褪黑素含量(P<0.05)。MCK 处理较CK 处理显著增加了30.21%,MST 处理较ST 处理显著增加了3.10%。
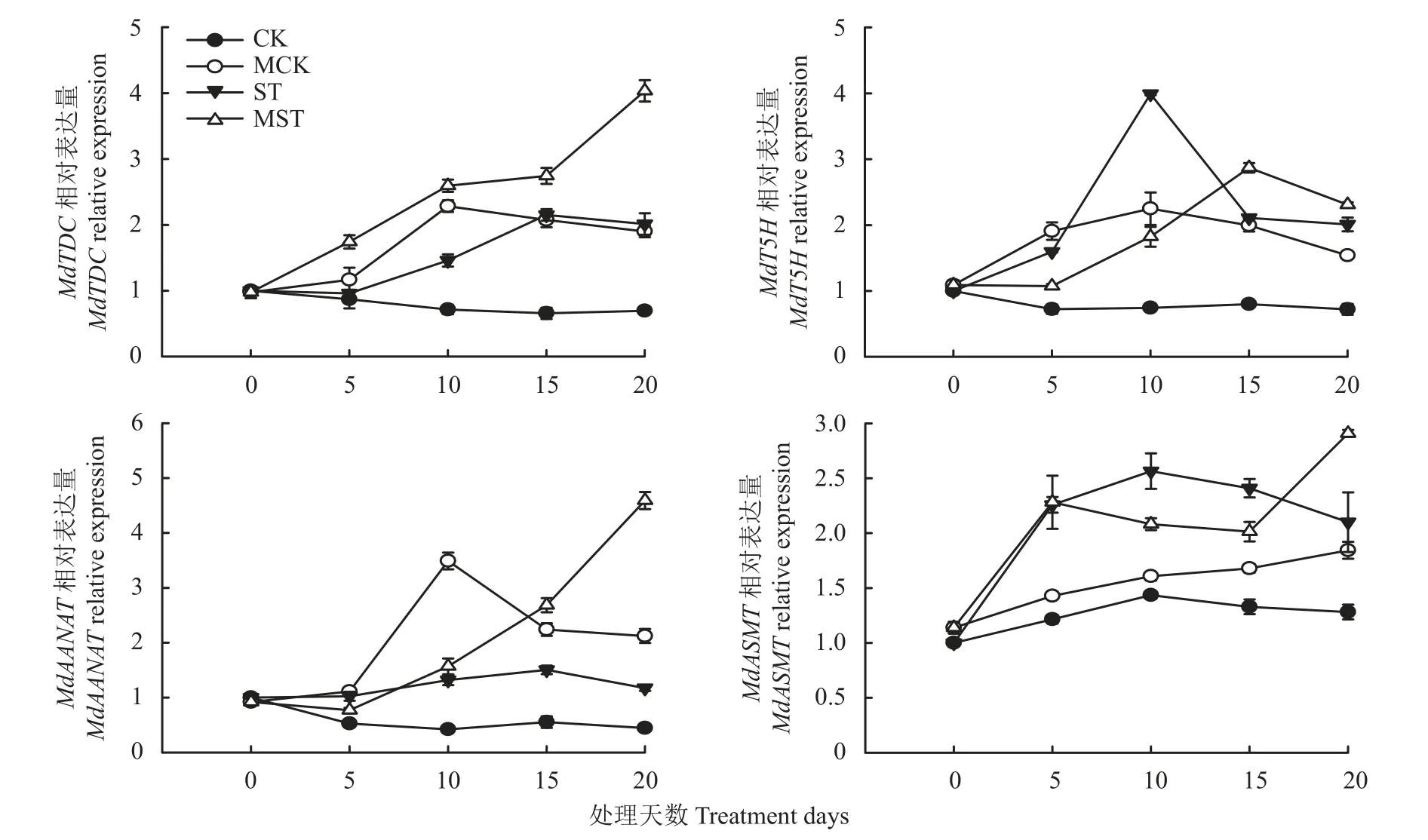
图1 外源褪黑素对养分正常和胁迫条件下苹果叶片中褪黑素合成相关基因表达量的影响Fig.1 Effects of exogenous melatonin on the relative expression of melatonin synthesis-related genes in M.hupehensis leaves under normal and low nutrient supply conditions

图2 外源褪黑素对养分正常和胁迫条件下苹果叶片中内源褪黑素含量的影响Fig.2 Effects of exogenous melatonin on endogenous melatonin content in M.hupehensis leaves under normal and low nutrient supply conditions
2.2 外源褪黑素对叶片和根中氮代谢关键酶活性的影响
如图3 所示,在养分正常和胁迫条件下,外源褪黑素均提高了苹果叶片中氮代谢酶活性。与CK相比,MCK 处理显著提高了叶片中的NR、NiR、GS、Fd-GOGAT 和NADH-GOGAT 活性(P<0.05),分别较CK 提高了28.13%、13.72%、17.36%、9.34%和26.82%;ST 处理显著降低了叶片中的NR、NiR、GS、Fd-GOGAT 和NADH-GOGAT 活性(P<0.05),分别较CK 降低了50.00%、36.75%、29.66%、14.01%和36.59%;与ST 处理相比,MST 显著提高了养分胁迫下叶片中的NR、NiR、GS 和Fd-GOGAT 活性(P<0.05),分别较ST 处理提高了65.62%、51.49%、59.63%和17.53%。

图3 外源褪黑素对养分正常和胁迫条件下苹果叶片中NR、NiR、GS、Fd-GOGAT 和NADH-GOGAT 酶活性的影响Fig.3 Effects of exogenous melatonin on the activities of NR, NiR, GS, Fd-GOGAT and NADH-GOGAT in M.hupehensis leaves under normal and low nutrient supply conditions
如图4 所示,在养分正常和胁迫条件下,外源褪黑素均提高了苹果根中氮代谢酶活性。与CK 相比,MCK 处理显著提高了叶片中的NADH-GOGAT活性(P<0.05),较CK 提高了26.88%;ST 处理显著降低了根中的NR、NiR、GS、Fd-GOGAT 和NADHGOGAT 活性(P<0.05),分别较CK 降低了25.00%、9.56%、43.25%、15.88%和44.04%;与ST 处理相比,外源褪黑素显著提高了养分胁迫下根中的NiR、GS 和Fd-GOGAT 活性(P<0.05),分别提高了19.52%、52.09%和12.72%。
2.3 外源褪黑素对叶片和根中氮代谢相关基因表达的影响
如图5 所示,处理第20 天时,在养分正常和胁迫条件下,外源褪黑素均上调了苹果叶片中氮代谢相关基因的表达水平。养分正常条件下,外源褪黑素显著上调了NR、NiR和GS基因的表达水平(P<0.05),MCK 处理分别为CK 处理的1.37、1.77 和1.40 倍。ST 处理的幼苗叶片中NR、GS和NADHGOGAT基因的表达水平显著下调(P<0.05),分别较CK 降低了51.07%、37.64%和24.51%。与ST 处理相比,外源褪黑素显著提高了养分胁迫下叶中NR、NiR、GS、Fd-GOGAT和NADH-GOGAT基因的表达水平(P<0.05),MST 处理分别为ST 处理的2.16、2.08、2.22、1.71 和1.19 倍。
如图6 所示,处理第20 天时,在养分正常和胁迫条件下,外源褪黑素均上调了苹果根中氮代谢相关基因的表达水平。养分正常条件下,外源褪黑素显著上调了NiR基因的表达水平(P<0.05),MCK 处理为CK 处理的1.88 倍。ST 处理根中的NR、NiR、GS、Fd-GOGAT和NADH-GOGAT基因的表达水平显著低于CK (P<0.05),分别较CK 降低了62.22%、60.25%、48.41%、59.14%和57.90%;MST 处理显著提高了养分胁迫下根中NiR、GS、Fd-GOGAT和NADH-GOGAT基因的表达水平(P<0.05),分别为ST 处理的1.31、1.41、1.74 和1.54 倍。

图6 外源褪黑素对养分正常和胁迫条件下苹果根部NR、NiR、GS、Fd-GOGAT和NADH-GOGAT 基因相对表达量的影响Fig.6 Effects of exogenous melatonin on the relative expression of NR, NiR, GS, Fd-GOGAT and NADH-GOGAT genes in M.hupehensis roots under normal and low nutrient supply conditions
2.4 外源褪黑素对叶片中氮转运相关基因表达的影响
如图7 所示,处理20 天内,MCK 处理叶片中NO3-转运基因MdNRT1.1、MdNRT2.5、MdNRT2.7的表达量呈现先升高后降低的趋势,NH4+转运基因MdAMT1.2、MdAMT1.5和MdAMT2.1的相对表达量呈上升趋势,ST 和MST 处理叶片中NO3-转运基因MdNRT1.1、MdNRT2.5、MdNRT2.7和NH4+转运基因MdAMT1.2、MdAMT1.5和MdAMT2.1的相对表达量呈现先升高后降低的趋势,在处理第10 天或第15 天达到峰值,这些基因的表达在CK 处理的叶片中相对稳定。处理第20 天时,与CK 处理相比,ST 处理叶中的MdNRT1.1、MdNRT2.5、MdNRT2.7、MdAMT1.2、MdAMT1.5和MdAMT2.1的表达量上调。与ST 处理相比,外源褪黑素提高了MdNRT2.4、MdNRT2.5、MdNRT2.7、MdAMT1.2和MdAMT2.1的表达水平。MST 处理的叶片中MdNRT2.4的表达水平呈现上升趋势,处理第20 天值最高,为ST 处理的9.23 倍。
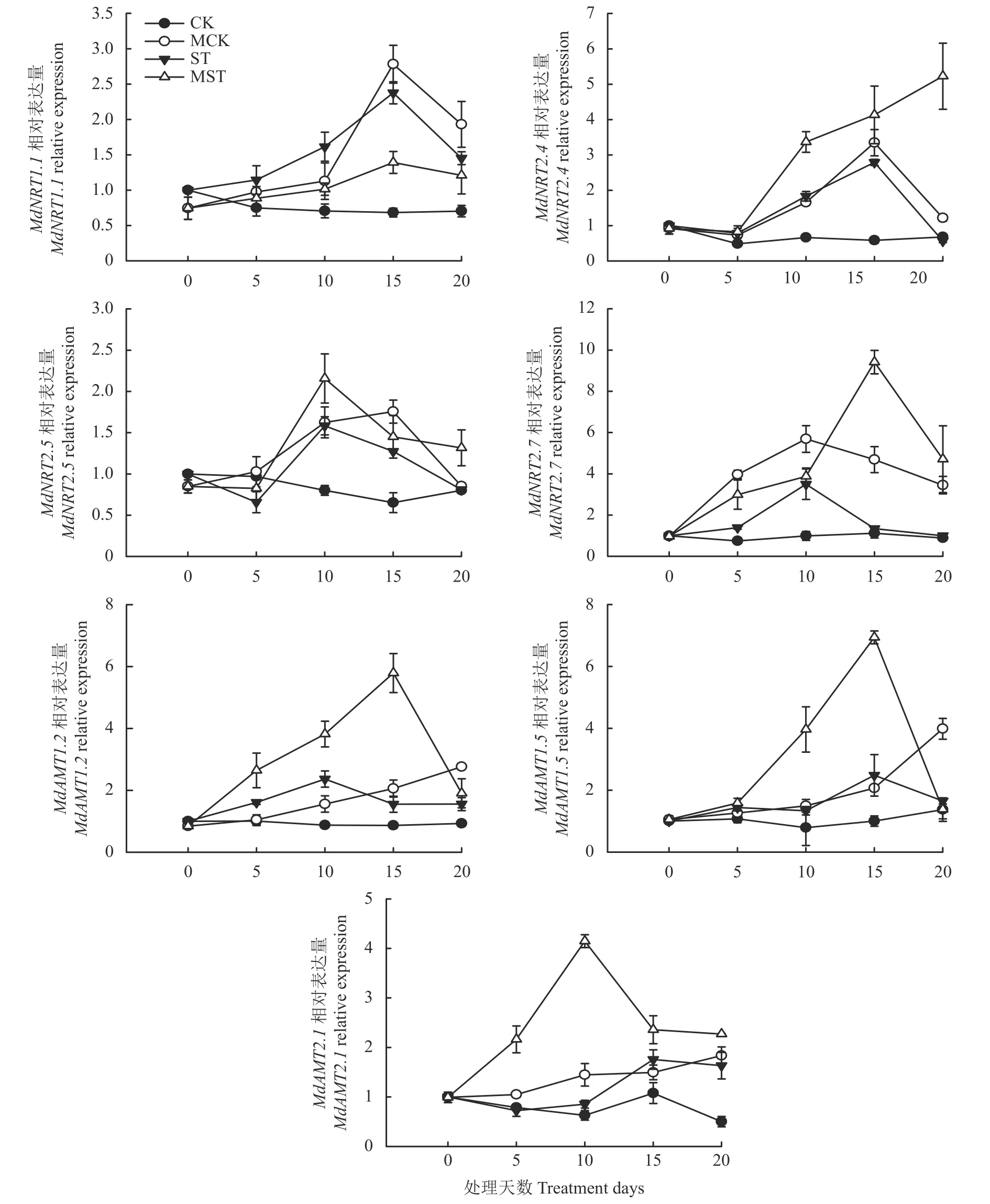
图7 外源褪黑素对养分正常和胁迫条件下平邑甜茶幼苗叶片中氮转运相关基因表达的影响Fig.7 Effects of exogenous melatonin on the expression of nitrogen transport-related genes in leaves of M.hupehensis under normal and low nutrient supply conditions
3 讨论
当土壤中的营养元素缺乏时,易导致植物产生缺素症状[2],褪黑素作为抗氧化剂,可以通过调节关键基因表达来应对环境胁迫[18]。本试验通过水培来模拟养分胁迫,通过测定平邑甜茶幼苗的褪黑素合成基因表达量、褪黑素含量、氮代谢酶活性以及氮代谢和转运相关基因表达量,探究外源褪黑素对养分胁迫下苹果幼苗氮代谢和转运的影响。结果表明,对养分胁迫下的苹果幼苗外源施加褪黑素,能上调平邑甜茶幼苗叶中褪黑素合成基因的表达水平,显著提高苹果幼苗叶中褪黑素含量,提高苹果幼苗叶和根中氮代谢酶活性,增强氮代谢和转运相关基因的表达。
在植物正常生长条件下,褪黑素水平可以保持相对稳定的状态。然而,褪黑素水平受环境胁迫的强烈影响。研究表明,植物中褪黑素的形成始于色氨酸,包括4 个酶促步骤,由色氨酸脱羧酶(TDC)、色氨酸羟化酶(TPH)、色胺-5-羟化酶(T5H)、5-羟色胺-N-乙酰基转移酶(SNAT)、5-羟色胺-N-乙酰基转移酶(ASMT)等进行催化,因此,TDC、T5H、SNAT、ASMT 等酶的相关基因参与褪黑素合成过程[12,19]。干旱胁迫下,桃苗的ASMT和TDC表达显著上调,经褪黑素处理的植株表达量进一步升高[19],上调褪黑素生物合成基因MdTDC1、MdAANAT2、MdT5H4和MdASMT1的表达可以调节干旱胁迫下平邑甜茶植株的水分平衡[20]。此外,外源施加褪黑素可以增加内源褪黑素含量[21-22],也可以自我调节褪黑素合成基因的表达[18]。本试验结果表明,养分胁迫下,苹果幼苗叶片中褪黑素合成相关基因的表达上调,褪黑素含量升高,同时,外源施加褪黑素会提高养分胁迫下苹果幼苗叶片中褪黑素合成相关基因MdTDC、MdT5H、MdAANAT和MdASMT的表达水平,增加内源褪黑素的含量。这表明养分胁迫一定程度上会刺激褪黑素合成基因的表达,致使褪黑素含量增加以响应养分胁迫,外源褪黑素可以进一步促进褪黑素合成基因的表达,提高内源褪黑素的含量。
NR、NiR、GS、Fd-GOGAT 和NADH-GOGAT是氮代谢过程的关键酶,硝酸盐在NR 的催化下变成亚硝酸盐,NiR 将亚硝酸盐还原成铵,GS 参与植物体内的氨同化过程,将植株体内的NH4+催化成谷氨酰胺,Fd-GOGAT 和NADH-GOGAT 将其催化成谷氨酸,参与植物体内氨基酸循环,GOGAT 能够与GS 结合,完成氨同化[4]。研究表明,氮磷亏缺会严重影响苹果叶片氮代谢酶活性,低氮情况下,苹果砧木的NR 活性低于正常供氮[7],适氮低磷与低氮适磷均会导致嘎啦植株中NR 和NiR 活性降低[23]。低氮胁迫降低了平邑甜茶幼苗叶片的NR、NiR、GS与GOGAT 活性,外源褪黑素会缓解氮代谢酶活性的降低[4]。本试验结果显示,与CK 处理相比,ST 处理的叶和根中NR、NiR、GS、Fd-GOGAT 和NADHGOGAT 活性降低,外源褪黑素可以减小下降幅度。褪黑素可以调控氮代谢相关基因的表达。研究表明,低氮条件下植物NR、NiR、GS的表达显著下调,外源褪黑素能增强低氮条件下植物NR、NiR、GS、Fd-GOGAT和NADH-GOGAT基因的表达[4,24],这与本研究的结果一致。表明低氮胁迫会抑制氮代谢关键酶活性及氮代谢基因表达,外源褪黑素可以通过提高氮代谢关键酶活和氮代谢基因的表达水平,从而提高苹果幼苗氮代谢水平。
褪黑素能够缓解养分胁迫导致的植物氮转运受阻现象。研究表明,低氮胁迫会促进植株NRT和AMT家族部分基因的表达,如低氮胁迫下,苹果植株的NRT1.1基因表达量上调[13],油菜的BnAMT1.1、BnAMT1.2、BnAMT1.5、BnAMT1.4和BnAMT2.1以及BnNRT2.4a、BnNRT2.4b、BnNRT2.5a和Bn NRT2.5b的表达上调[25],本试验中,ST 处理叶中的NRT1.1、AMT2.1、AMT1.2和AMT1.5的表达量上调,表明养分胁迫可能一定程度上提高了平邑甜茶幼苗对NO3-和NH4+的吸收与转运能力。低NH4+胁迫下,外源褪黑素不同程度上调了AMT1.2、AMT1.5、AMT2.1、NRT1.1的表达水平,促进了平邑甜茶叶片或根部对NH4+的吸收和转运[4]。本研究中,养分胁迫下,外源褪黑素提高了NRT2.4、NRT2.5、NRT2.7、AMT1.2和AMT2.1的表达水平,表明外源褪黑素可能通过增强平邑甜茶幼苗的NH4+和NO3-转运基因表达,来促进苹果植株对氮的吸收与转运。
4 结论
养分胁迫下,平邑甜茶幼苗叶片中褪黑素合成基因的表达上调,内源褪黑素含量显著提高,苹果幼苗叶片及根中氮代谢酶活性显著降低,氮代谢相关基因表达水平显著下调,氮转运相关基因的表达上调,表明苹果幼苗在养分胁迫条件下通过提高内源褪黑素合成基因的表达,提高对氮的利用。添加外源褪黑素强化了内源褪黑素的效果,尤其在养分胁迫条件下,总之,外源褪黑素可以通过促进苹果植株对氮的代谢和吸收转运来缓解养分胁迫。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
家庭影院技术(2021年7期)2021-08-14
西北农林科技大学学报(自然科学版)(2021年2期)2021-03-09
落叶果树(2021年6期)2021-02-12
文苑(2020年12期)2020-04-13
西藏农业科技(2019年3期)2019-11-04
海峡姐妹(2019年1期)2019-03-23
中国果业信息(2019年10期)2019-01-05
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
