
大别山特有种都支杜鹃的遗传多样性和遗传结构1)
2023-11-24 05:30赵凯章伟裴亚丽翟伟
东北林业大学学报 2023年12期
赵凯 章伟 裴亚丽 翟伟
(安庆师范大学,安庆,246011) (安徽大学) (安庆师范大学)
杜鹃花属(Rhododendron)植物因其花繁叶茂、绮丽多姿成为世界著名的木本花卉[1-2],自从19世纪被大量发现并引种到欧洲以来,就在园艺学上占据了相当重要的地位[3]。都支杜鹃(RhododendronshanniFang)隶属于杜鹃花科杜鹃花属常绿亚属植物,具有极佳的观赏特性[4-6]。该种现仅分布于我国大别山区岳西县、霍山县及英山县的三县交界处,生于海拔1 400 m以上的山坡或山脊上,分布区面积不足110 km2,种群总占地面积不到1.2 km2[7-8],是亟需保护且具有较高开发价值的狭域片段化分布的濒危物种[7-9]。
目前,对都支杜鹃的研究主要集中在形态解剖学和种群生态学[7,10-11],而基于分子数据对都支杜鹃的研究还相对缺乏,仅见赵凯等[11]基于ISSR分子标记分析了都支杜鹃的遗传多样性,Pan et al.[9]筛选了都支杜鹃24对多态性EST-SSR引物,但这些研究并没有清楚地揭示都支杜鹃的遗传结构特征。EST-SSR是基于表达序列标签开发微卫星的一种新型分子标记,与基因组SSR相比,EST-SSR具有在植物物种之间可转移性的优点,目前已被广泛应用于植物基因组学研究,如遗传多样性评价、种质鉴定、系统发育与进化研究等方面[12]。鉴于此,本研究利用24对多态性EST-SSR引物,进一步对濒危物种都支杜鹃自然种群的遗传多样性和遗传结构进行研究,旨在揭示都支杜鹃的遗传多样性和遗传背景,为濒危植物都支杜鹃的保护和合理开发利用奠定基础。
1 材料与方法
1.1 样品采集
2016年5月在都支杜鹃的分布区域内选取了5个具代表性的自然种群进行采样,共采集158个个体,其中白马尖(BMJ)种群33个个体、天河尖(THJ)种群30个个体、都支尖(DZJ)种群33个个体、石壁沟(SBG)种群26个个体、坨尖(TJ)种群36个个体。采样时每株采集5~10片嫩叶于硅胶中干燥后带回实验室,植株间的间隔大于50 m。采样点之间最近直线距离4.01 km,最远直线距离为11.29 km(图1)。
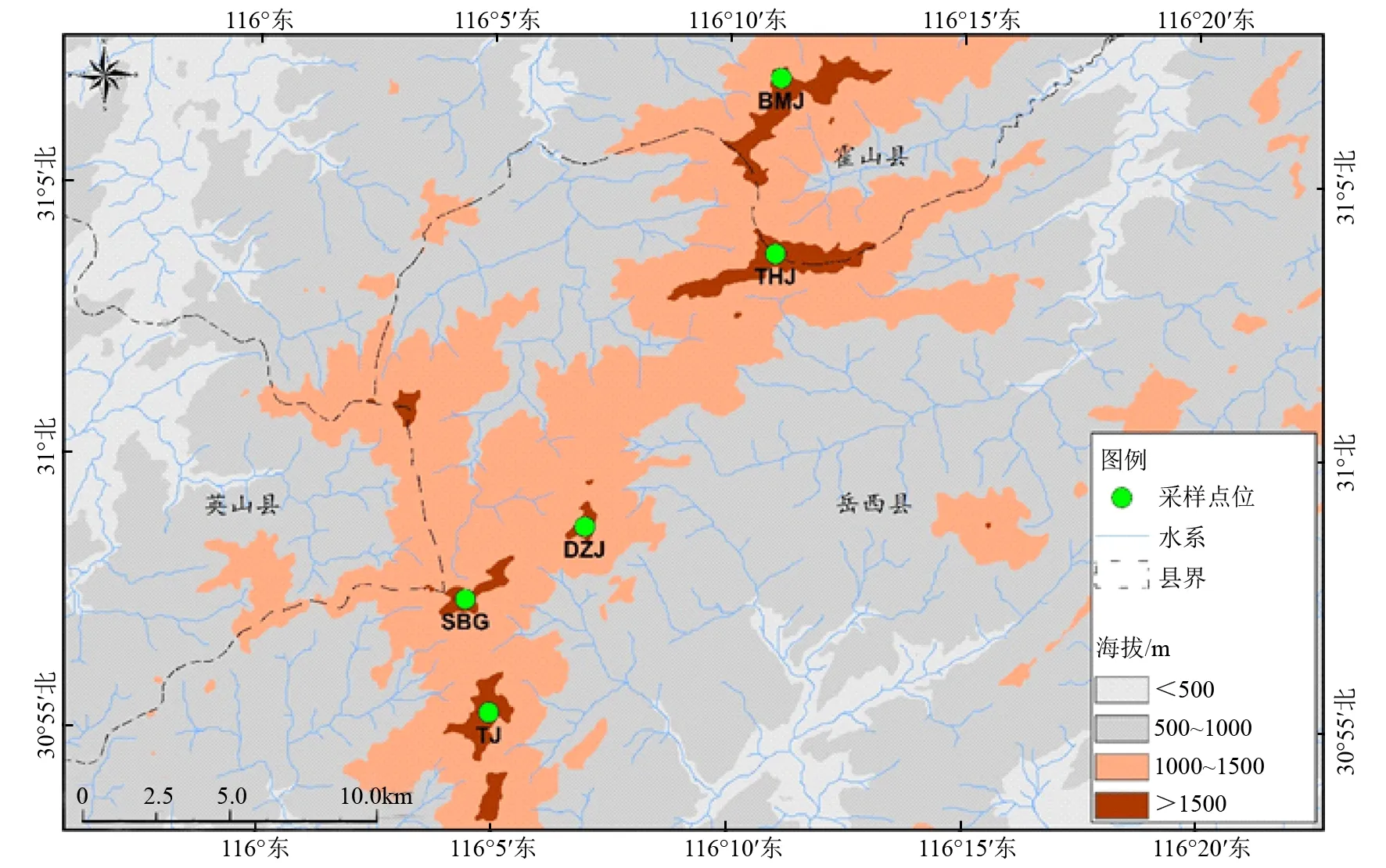
图1 都支杜鹃采样点位分布图
1.2 DNA提取与扩增结果检测
采用改良CTAB法进行样品DNA提取。利用Pan et al.[9]开发的24对都支杜鹃EST-SSR引物对每个样品进行PCR扩增,扩增后的产物使用2%的琼脂糖凝胶电泳进行检测,检测合格的样品送至生物科技公司进行测序分析。使用GeneMarker 1.85对分型的结果进行分析读数,获取微卫星基因型数据。
1.3 数据处理
在遗传多样性上,使用Popgene 3.2软件进行期望杂合度和观测杂合度的计算。使用FASTA 2.9.3软件计算各居群的平均等位基因数和种群近交系数。使用GenALEx 6.5软件进行遗传分化系数、遗传距离和地理距离的计算,同时利用公式Nm=0.25(1-Fst)/Fst计算种群间的基因流(式中,Nm代表基因流,Fst代表遗传分化系数)。利用Ntsys软件进行Mantel检验分析种群间遗传距离与地理距离之间的相关性。在遗传结构上,使用Powermaker软件计算居群间的Nei’s遗传距离并使用MEGA 7.0软件绘制居群间的UPGMA聚类树[13]。使用Structure 2.3软件对所有个体单独聚类,K值设置为2~8,重复15次;burn-in设置为100 000,run-length设置为1 000 000,运行完成后用Structure Harvester分析STRUCTURE运行结果得到最佳分组的K值[14-16]。使用GenALEx 6.5软件进行主坐标(PCoA)分析并制图[17]。最后,使用Arlequi 3.5软件进行AMOVAs分析[18]。
2 结果与分析
2.1 都支杜鹃种群的遗传多样性
采用24对EST-SSR引物对都支杜鹃5个天然种群158个个体的遗传多样性分析结果显示该物种的遗传多样性水平较高(表1),平均等位基因的均值为7.602,期望杂合度(HE)的均值0.665,观测杂合度(HO)的均值为0.712。其中SBG种群的遗传多样性最低(平均等位基因=6.38,HE=0.589,HO=0.649),BMJ种群的HE最高为0.700,DZJ种群的HO最高为0.759,TJ种群的平均等位基因最高为8.333。此外,所有种群的观测杂合度均大于期望杂合度。近交系数分析结果表明,都支杜鹃5个种群的Fis值均小于0,说明大别山地区的都支杜鹃种群主要为远交种群。
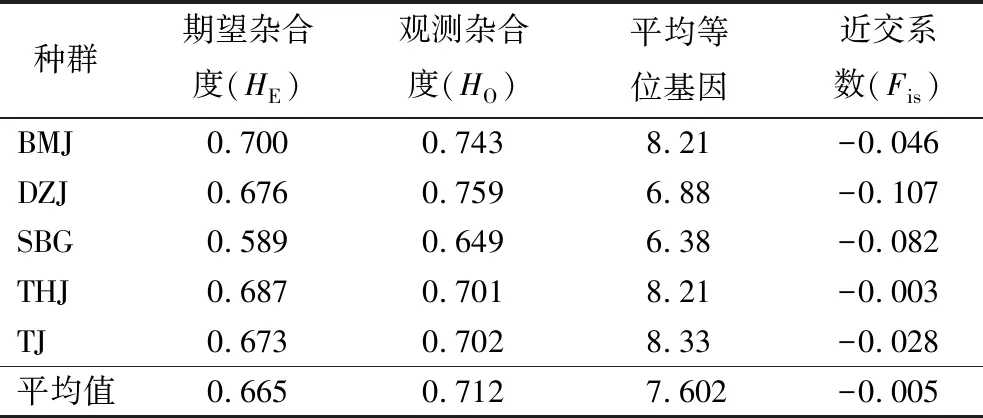
表1 都支杜鹃的遗传多样性和近交系数
2.2 都支杜鹃种群的遗传分化和遗传结构
遗传分化系数(Fst)计算结果显示都支杜鹃5个种群间的遗传分化系数均较低,均值为0.041,变化范围为0.024~0.059(表2),表明都支杜鹃在种群水平上的分化程度较低。基因流(Nm)计算结果显示,都支杜鹃5个种群间基因流大小在3.976~10.327波动,均值为6.40,表明都支杜鹃各种群间存在比较频繁的基因交流,这与都支杜鹃种群间遗传分化小的结果相一致。Mantel检验结果表明:都支杜鹃各种群间的遗传距离与地理距离之间的相关性系数为R2=0.157(P=0.666),表明都支杜鹃各种群间的遗传距离与地理距离无相关性。
UPGMA聚类分析结果显示:在Nei’s遗传距离0.105处,都支杜鹃的5个天然种群可以分为两个集群,其中,DZJ种群为一个集群,剩下的4个种群个体(BMJ、THJ、SBG、TJ)聚为一个集群(图2)。基于Structure软件的遗传聚类分析结果表明,ΔK的峰值给出的最佳结果为K=2。当K=2时,5个都支杜鹃种群可以被分为两个谱系,其中DZJ种群的大部分个体聚为红色谱系,剩下的4个种群个体(BMJ、THJ、SBG、TJ)聚为绿色谱系(图3)。主坐标分析(PCoA)结果显示,都支杜鹃5个种群的158个个体可以明显聚为两类,其中DZJ种群的大部分个体聚为一类,剩下的4个种群个体(BMJ、THJ、SBG、TJ)聚为一类(图4),这一结果与主坐标分析Structure软件结果基本一致。进一步的分子方差(AMOVA)分析结果表明,两个谱系间的遗传变异占总体变异的2.43%,谱系内种群间变异占总体变异的4.60%,而种群内部变异占总体变异的92.97%(P<0.001),说明都支杜鹃种群的遗传变异主要存在于种群内(表3)。

表3 都支杜鹃5个种群的分子方差分析
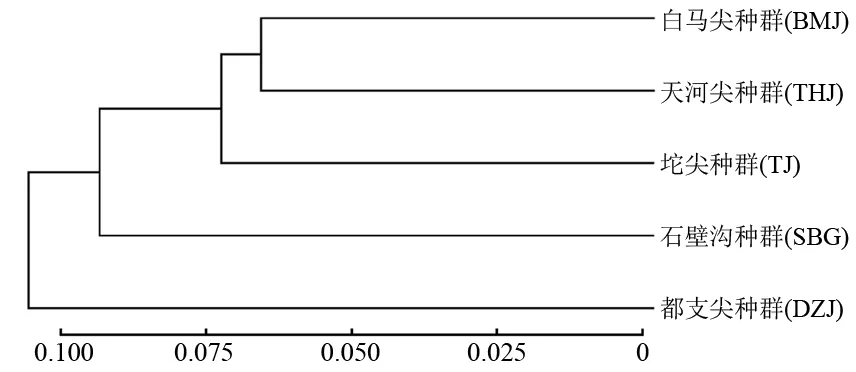
图2 基于Nei’s遗传距离构建的5个都支杜鹃居群间的UPGMA聚类树
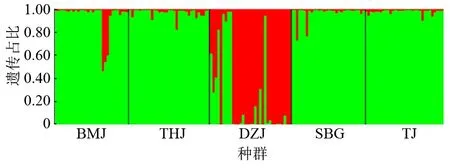
BMJ为白马尖种群;DZJ为都支尖种群;SBG为石壁沟种群;THJ为天河尖种群;TJ为坨尖种群。
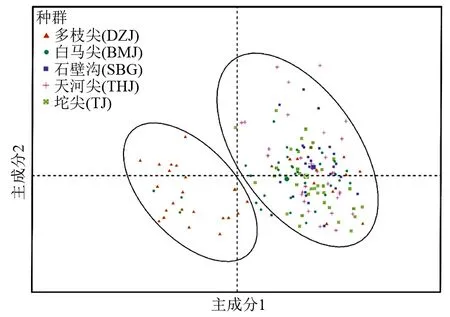
图4 都支杜鹃5个种群的主坐标分析(PcoA)结果
3 结论与讨论
本研究基于24个多态性EST-SSR位点对都支杜鹃5个野生种群的158个个体进行遗传多样性和遗传结构分析,结果显示各种群均具有较高的遗传多样性(5个种群的等位基因数、观测杂合度和期望杂合度均值分别为7.602、0.712和0.665),这与赵凯等[11]2013年基于ISSR数据分析都支杜鹃的遗传多样性结果相反。赵凯等[11]2013的研究中ZTJ、DYJ、BMJ 3个种群地理距离较近(BMJ种群距离ZTJ和DYJ分别为0.88和0.069 km),且ZTJ和DYJ种群均只包含几十株个体。本研究将这3个种群并为BMJ一个种群(未采集ZTJ和DYJ的样品),新增了THJ和TJ两个种群,这两个种群数量分别为1 000、2 000株,占了都支杜鹃野生个体总数的60%。考虑到本次采样范围更全面、样点设置更具代表性,加上新型分子标记EST-SSR的应用,本研究揭示的都支杜鹃遗传多样性应该更能反应真实情况。
多数研究表明,对于狭域分布的濒危物种通常具有较低的遗传多样性[19-20],也有部分物种保持了较高的遗传多样性,如毛果木莲(Manglietiaventii,H=0.654)[21]、银缕梅(Parrotiasubaequalis,Ho=0.520,He=0.558)[22]、长果安息香(Changiostyraxdolichocarpa,Ho=0.594,He=0.643)等[22]。一般认为,遗传多样性高的濒危种在历史上可能有广泛的分布,后期经历的某种地质气候事件导致其种群规模的快速减小,使其丰富的遗传多样性仍保留在有限的居群和个体内[23-24],这可能也是都支杜鹃拥有较高遗传多样性的原因。此外,都支杜鹃各居群的自交系数均为负数(均值为-0.005),表明其为典型的远交物种,这也可能是其维持较高遗传多样性的原因之一。
对于种群规模近期快速减小且呈岛屿状分布的物种,它们种群间的遗传分化一般较小,遗传变异主要存在种群内[23-24]。本研究对都支杜鹃5个种群间的遗传分化系数分析表明,其种群间的遗传分化系数范围为0.024~0.059,均较小。此外,分子方差分析结果表明种群内的遗传变异占总体变异的92.97%。这些结果均支持都支杜鹃的种群规模是近期快速减小的假设,即现在的种群规模和种群分布式样可能是近期形成的,各种群内部都保留了其祖先种群高的遗传多样性,且在短期内没有发生明显的遗传变异。UPGMA聚类分析、Structure软件聚类分析和主坐标分析(PCoA)结果基本一致,均显示研究的5个种群可以分为两个谱系,即DZJ种群的大部分个体聚为一个谱系,剩下的所有个体聚为另一个谱系。而分子方差(AMOVA)分析却显示两个谱系间的遗传变异很小,且不存在差异显著性。从地理分布来看,DZJ种群处于相对中间的地理位置,与其他种群间较大的基因流(Nm均值5.20)可能是导致谱系间遗传变异占比小的原因,DZJ种群独特的遗传结构可能与遗传漂变有关。在现存的都支杜鹃种群中,DZJ种群占地面积最小,数量也最少(仅存不到100株),这加剧了遗传漂变。同时也有可能与其生境有关,现存都支杜鹃基本都生存在海拔1 500 m以上的陡峭悬崖之上,DZJ种群周围距离最近的类似生境最近直线距离在3 km以上,这加剧了DZJ种群的孤岛效应。
尽管都支杜鹃的遗传多样性较高,但是其现存成年个体仅有5 400株左右,且主要呈岛屿状分布在大别山区江淮分水岭的高海拔地区、生存环境恶劣[7],受当代气候波动,特别是全球气候变化的影响仍然很大。所以对都支杜鹃的保护应该以就地保护为主,同时对一些小种群可以采用人工辅育的方式进行种群恢复。此外,位于岳西县鹞落坪多枝尖的DZJ种群与其它种群在遗传上有一定程度的分化,且该种群是该种模式产地之一,仅存100株左右,建议应特别注意对该种群的保护。后续还应加强对都支杜鹃全面采样,基于基因组水平的研究为其提供更准确、合理的保护意见。
猜你喜欢
艺术品鉴(2022年16期)2022-07-09
河池学院学报(2021年1期)2021-07-10
种子(2021年3期)2021-04-12
景德镇陶瓷(2021年1期)2021-03-24
海南医学(2020年1期)2020-01-18
电子制作(2019年24期)2019-02-23
东方考古(2017年0期)2017-07-11
外语教学理论与实践(2016年1期)2016-06-11
广西林业科学(2016年3期)2016-03-16
中国康复理论与实践(2015年10期)2015-12-24
