
12 个抗稻瘟病基因在山东省地方水稻品种中的分布
2023-11-23 03:26房文文王海凤郭涛姜艳芳薛芳张焕霞张士永
山东农业科学 2023年10期
房文文,王海凤,郭涛,姜艳芳,薛芳,张焕霞,张士永
(山东省农业科学院湿地农业与生态研究所/山东省水稻工程技术研究中心,山东 济南 250100)
水稻是世界上的主粮作物之一,近一半的人口以稻米为主食[1-2]。 稻瘟病是水稻生产上最重要的病害,严重影响水稻产量和品质[3-4]。 山东省一般5 年左右会暴发一次,发病田块一般减产10%~30%,严重的甚至超过50%[5]。 山东属黄淮海稻区,以粳稻为主,年均种植面积达13 万公顷[6]。 稻瘟病的发病条件为气温26 ~28 ℃、相对湿度90%以上,临沂、日照等稻区水稻抽穗灌浆期的气候特点符合其发生流行的条件,是山东省稻瘟病发生较重的地区[7]。 目前,选育和合理种植抗病品种是控制稻瘟病流行最经济有效的途径[8-9]。 由于田间稻瘟病菌小种类型多样,易发生变异,一些抗病水稻品种连续种植3 ~5 年后抗性就会丧失,田间表现为感病,从而造成减产[10]。
随着水稻基因组学和测序技术的迅速发展,越来越多的抗稻瘟病基因被定位与克隆,目前已被定位的抗瘟基因有100 多个,已克隆的抗瘟基因超过30 个[11]。 抗瘟基因多为显性基因,其作用方式多为通过与核苷酸结合位点-富含亮氨酸重复序列蛋白(NBS-LRR)结合,从而使病原菌失去侵染性[12-13]。 近年来,抗瘟基因分子标记的开发,主要为SSR、Indel、SNP 标记,为高效检测抗瘟基因奠定了基础[14-16]。 张亚玲等利用35 个抗瘟基因标记检测了黑龙江省50 个水稻主栽品种,发现抗瘟基因Pish、Pi36、Pi33和Pi-CO39在供试品种中的出现频率为100%,Pi63、Ptr、Pi37、Pi64、pi21、Pi9、Pi54、Pia、Pikp、Pi35、Pikm和Pik的出现频率为50%~100%,Pita、Pib、Pii、Pi5、Piz-t、Pi50和Pi2的出现频率为10%~50%,Pid2仅在2个品种中被检测到,Pigm仅在1 个品种中被检测到,而Pit、Pid3、Bsr-d1、Pi25、Pid3-A4、Pi56、Pi1、Pike和Pb1在供试品种中未检测到[17];杨林浩等利用一套稻瘟病菌标准菌株和特异性分子标记相结合,检测了吉林省68 个主栽水稻品种,发现Pita、Pia、Pish、Pita2、Pib和Pi9为吉林省主栽品种中出现频率较高的抗瘟基因,推测所有品种可能不含有Pi2、Pi54、Pikm、Pi11和Pi12基因[18]。
山东省作为重要的粳稻产区,有着丰富的地方品种资源,但有关这些品种资源的抗瘟基因分布未见报道。 本研究利用Pita、Pita2、Pia、Pi9、Pigm、Pikm、Pi54、Pi5、Pii、Pib、Pish、Pi1共12 个抗瘟基因的分子标记对78 个山东省地方品种进行抗瘟基因检测,以初步明确这些品种的抗瘟基因组成,为今后进一步挖掘和利用抗瘟基因、培育持久广谱抗瘟品种提供基因信息和数据支持。
1 材料与方法
1.1 试验材料
供试材料为78 个山东省地方水稻品种,见表1,均由山东省农业科学院湿地农业与生态研究所种质资源团队收集和保存,均以常规方法栽培。

表1 供试山东省地方水稻品种
试剂及仪器: 十六烷基三甲基溴化铵(CTAB),购于生工生物工程(上海)股份有限公司;PCR 试剂,北京全式金生物技术(TransGen Biotech) 有限公司产品;EvaGreenqPCR 核酸染料,美国Biotium 公司产品。 SPX 智能型生化培养箱、RXZ-500C-LED 人工气候培养箱,宁波江南仪器厂产品;BCD-290W 型立式冰箱,青岛海尔股份有限公司产品;罗氏LightCycler96 实时荧光定量PCR 仪,Roche 公司产品。
1.2 水稻DNA 提取
剪取3 段供试水稻品种的叶片(大小为2 mm×2 mm)于2 mL 离心管中,采用CTAB 法提取DNA。具体步骤如下:
(1)向装有供试水稻叶片的离心管中加600 μL 65 ℃预热的2×CTAB,然后65 ℃温浴30 min;
(2)冷却后加600 μL 氯仿-异戊醇溶液(氯仿和异戊醇体积比为1∶1)混匀,12 000 r/min 离心10 min,吸200 μL 上清液,加入等体积的异丙醇,轻轻颠倒混匀后于-20 ℃条件下沉淀30 min;
(3)12 000 r/min、4 ℃条件下离心15 min,弃上清液,加入600 μL 体积百分含量为70%的乙醇洗涤沉淀1~2 次,放入通风橱内吹干;
(4)加入100 μL TE(10 mmol/L Tris-HCl,0.1 mmol/L EDTA,pH 8.0)溶解DNA,使用核酸定量仪测量DNA 浓度,并将浓度调整为约10 ng/μL,-20℃保存待测DNA。
1.3 抗瘟基因分子标记
本研究选用抗瘟基因Pita、Pi9、Pikm、Pigm、Pia、Pi54、Pish、Pii、Pi1、Pib、Pita2、Pi5的分子标记进行试验,其PCR 特异性引物信息见表2。 所有引物均由华大基因有限公司合成。

表2 抗瘟基因的特异性引物
1.4 PCR 扩增
荧光PCR 反应体系:10×Taqbuffer(含20 mmol/L Mg2+)1 μL,250 μmol/L dNTP 0.2 μL,0.5 UTaqDNA Polymerase 0. 1 μL,20 × Evagreen 0.125 μL,10 ng/μL DNA 模板3 μL,0.1 mmol/L正、反向引物各0.2 μL,用ddH2O 定容至10 μL。荧光PCR 扩增程序:94 ℃预变性2 min;94 ℃变性10 s,60 ℃退火30 s,40 个循环。
普通PCR 反应体系:2× ESTaqMaster Mix(Dye)7.5 μL,水稻DNA 3.0 μL,正、反向引物各0.5 μL,3.5 μL ddH2O。 PCR 反应程序:95 ℃预变性5 min;95 ℃变性30 s,55 ~60 ℃退火30 s(根据梯度PCR 的结果选择最佳退火温度),72 ℃延伸0.5~1.5 min(由扩增产物的长度决定),32 次循环;72 ℃延伸10 min。
1.5 琼脂糖凝胶电泳检测
抗瘟基因Pish、Pii、Pib、Pita2、Pi5的扩增产物用1%琼脂糖凝胶电泳检测,150 V 稳压电泳30 min,然后在凝胶成像系统中拍照。
1.6 HRM 检测
抗瘟基因Pita、Pi9、Pikm、Pigm、Pia、Pi54、Pi1的扩增产物经熔解曲线(HRM)检测。 分析程序:95 ℃变性15 s;60 ℃退火15 s;以0.07 ℃/s的速率升温,10 reading/℃采集荧光信号,至90℃保持1 s,采用Light Cycler 96 software 软件进行熔解曲线分析。
1.7 数据处理与统计分析
采用Microsoft Excel 对数据进行整理和作图。 采用NTSYS 2.10 软件计算78 个品种间的遗传相似系数,然后利用非加权类平均法(UPGMA)对遗传相似系数进行聚类分析,得到聚类树形图。
2 结果与分析
2.1 稻瘟病抗性基因的分子标记检测及在品种中的分布
本研究利用Pita、Pita2、Pia、Pi9、Pigm、Pikm、Pi54、Pi5、Pii、Pib、Pish和Pi1的功能标记对78个山东省地方水稻品种进行分子检测,结果(表3)发现,有53 个品种含有Pi54,有39 个品种含有Pia,有23 个品种含有Pi5,有17 个品种含有Pikm,有16 个品种含有Pib和Pii,有13 个品种含有Pi1,有1 个品种含有Pi9、Pish和Pita2,所有品种中均未检测到Pita和Pigm。 部分基因分子标记的检测结果如图1、图2 所示。

图1 高分辨率熔解曲线检测地方水稻品种的抗瘟基因

图2 琼脂糖凝胶电泳检测地方水稻品种的抗瘟基因Pi54、Pib、Pii

表3 山东省地方水稻品种抗瘟基因检测结果
Pi54和Pia在供试品种中的分布频率最高,分别高达67.95%和50.0%;其次是Pi5、Pikm、Pib、Pii和Pi1,分布频率分别为29.49%、21.79%、20.51%、20.51%和16.67%;分布频率较低的是Pi9、Pish和Pita2,均为1.28%;检测品种中都不含有抗性基因Pita和Pigm(表3、图3)。
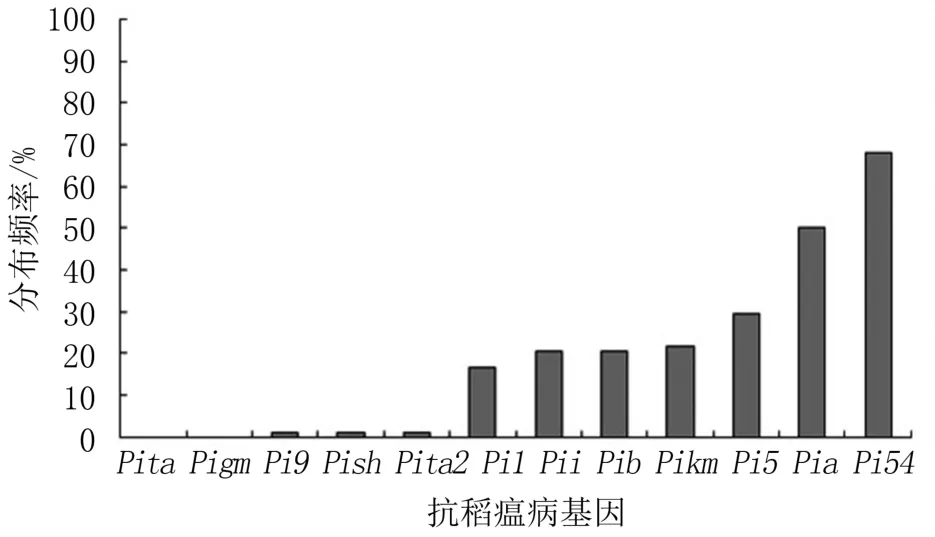
图3 12 个抗稻瘟病基因在山东省地方水稻品种中的分布频率
2.2 不同地方水稻品种含抗瘟基因的数量分析
由表3 和图4 可知,竹秆青-1、竹秆青-2、塘稻、明水米旱稻-1、明水香稻、紫秆旱稻等27 个品种检测出2 个抗瘟基因,占检测品种总数的34.62%,其中,有23 个品种检出Pi54,有15 个品种检出了Pia。 小黄稻、大青秸-3、小红芒、旱稻子、秋白水稻、蒋庄1 号等19 个品种检测出1 个抗瘟基因,占检测品种总数的24.36%,其中,有8个品种检测出Pi54。 粘旱稻-1、秋旱稻、高秸水旱稻-2、蓬莱稻-3、香稻、郯城野生稻等13 个品种检测出3 个抗瘟基因,占检测品种总数的16.67%。 另外,紫皮旱稻、水牛皮旱稻-1、青旱稻、蓬莱稻-1、葫芦香稻等7 个品种检测出4 个抗瘟基因,铁耙子、八月芒、乐陵旱稻、短顶芒白米4 个品种检测出5 个抗瘟基因,仅秃头红米稻检测出6 个抗瘟基因,红皮高丽旱稻-1、粘稻2 个品种检测出7 个抗瘟基因。 而大青秸-1、汪稻、水牛皮旱稻-1、老旱稻-2、旱粘稻-2 共5 个品种未检测到12 个抗瘟基因中的任何一个。

图4 山东省地方水稻品种含抗瘟基因数统计
2.3 山东省地方水稻品种的聚类分析
基于遗传相似系数,利用UPGMA 法对78 个山东省地方水稻品种进行聚类分析,结果(图5)显示,在遗传相似系数0.80 处,可将78 个供试材料分为四类。 第一类包括大青秸-1、汪稻及水牛皮旱稻-1 共3 个品种;第二类仅有高秸水旱稻-1;第三类包括大青秸-3、紫颖稻、粘旱稻-2 及白芒子共4 个品种;其余70 个品种均聚在第四类中。

图5 78 个山东省地方水稻品种的聚类分析结果
3 讨论与结论
水稻与稻瘟病菌存在协同进化关系,一个抗病新品种在生产上使用几年后,由于稻瘟菌变异而突破水稻的防御机制,使抗病品种的抗病能力下降甚至丧失,变为感病品种。 因此,在改良水稻品种的稻瘟病抗性时,要首先弄清品种中的抗瘟基因(组合),从而更好地利用不同的抗瘟基因组合对品种进行改良。 有关我国现有水稻品种中抗瘟基因的分布情况,前人已进行过一些研究,如岂长燕等研究了抗瘟基因Pib、Pita、Pi5、Pi25和Pi54在我国124 份水稻微核心种质中的分布[19];范方军等研究了Pib、Pita、Pi54和Pikm在江苏省迟熟中粳稻预试64 份品系中的分布[20]。 本研究利用12 个抗瘟基因(Pita、Pita2、Pia、Pi9、Pigm、Pikm、Pi54、Pi5、Pii、Pib、Pish、Pi1)的分子功能标记检测了78 个山东地方水稻品种的抗瘟基因分布情况,结果表明,除Pita、Pigm未检测到外,其余10 个基因均能在山东省地方水稻品种中检测到。 其中,Pi54和Pia在供试品种中的分布频率较高,达67.95%和50.0%;其次是Pi5、Pikm、Pib、Pii和Pi1,分布频率分别为29.49%、21.79%、20.51%、20.51%和16.67%;Pi9、Pish、Pita2的分布频率较低,仅为1.28%。 各品种含有抗瘟基因的数量为1~7 个不等,抗瘟基因组合种类丰富。
目前,培育持久广谱抗病品种和合理布局抗病品种仍是控制稻瘟病最经济有效的途径,而挖掘种质资源中的抗瘟基因是重要基础。 本研究结合山东省稻瘟病菌群体生理小种的变异动态,挑选出可进一步利用的抗瘟基因Pi54、Pia、Pi5、Pikm、Pib和Pii,下一步可作为目的基因利用到主栽品种中,这为育种家培育抗稻瘟病新品种提供了数据支持,助力了培育持久和广谱抗稻瘟病水稻新品种。 但需注意有些抗瘟基因与不良性状的“连锁累赘”效应,如Pigm与小粒性状连锁,以避免在提高抗病性的同时负向影响产量或品质。
本研究仅检测了12 个抗瘟基因在78 个山东省地方水稻品种中的分布情况,但其他抗瘟基因的分布情况还有待进一步研究;另外,地方品种中抗瘟基因数量与水稻品种稻瘟病抗性频率间的关系也有待进一步研究。
猜你喜欢
山东交通科技(2022年3期)2022-08-05
中国集体经济(2022年9期)2022-04-12
作物学报(2022年6期)2022-04-08
河南农业·综合版(2021年11期)2021-11-30
河南农业(2021年11期)2021-11-28
——山东省济宁市老年大学之歌
老年教育(老年大学)(2021年10期)2021-11-12
山东农业工程学院学报(2020年12期)2020-03-19
今日农业(2019年13期)2019-08-12
中国学校体育(2016年11期)2016-12-01
河南农业(2016年6期)2016-11-26
