
中华绒螯蟹螯足长节产地差异的几何形态测量学研究
2023-11-22 08:30:28薛竣仁刘洪波陈修报
安徽农业大学学报 2023年5期
唐 静,薛竣仁,刘洪波,姜 涛,陈修报,杨 健,*
中华绒螯蟹螯足长节产地差异的几何形态测量学研究
唐 静1,薛竣仁1,刘洪波2,姜 涛2,陈修报2,杨 健1,2*
(1. 南京农业大学无锡渔业学院,无锡 214081;2. 中国水产科学研究院淡水渔业研究中心渔业微化学实验室,中国水产科学研究院淡水渔业研究中心,无锡 214081)
为掌握中华绒螯蟹螯外形精细结构的产地差异性,首次应用几何形态测量学方法,对长江中下游湖南省、安徽省、江苏省等7个产地蟹的螯足长节形态进行了比较研究。依据地标点法选取原则建立了螯足长节的14个特征点数据系统,并通过将其标准化,获得此7个产地蟹的平均型, 再利用薄板样条分析和网格变形,对形态变异矢量进行了可视化分析。结果表明,与平均形比较,除无为野生蟹无明显的变化外,安徽省其他3个产地蟹长节形态有同趋势变化(即长节左下端向内缩小,长节内部凹陷点向左下端靠近);湖南、江苏3个产地蟹长节形态亦有同趋势变化(即长节左下端向外扩展,长节内部凹陷点向右上方向偏移)。此外,对上述标准化数据开展判别分析的结果更显示,7个产地雌、雄蟹个体间的判别准确率均可达100%。上述发现进一步证实了几何形态测量学法不仅在大至背甲,小至螯足长节的形态方面能有效把握中华绒螯蟹产地差异性,而且在非致死、致伤产地溯源技术研发上也极具潜力。
中华绒螯蟹;几何形态测量学;地标点;螯足长节;产地溯源
中华绒螯蟹(俗称大闸蟹、河蟹,)是一种降河洄游性蟹类,肉鲜味美,人体所需的蛋白质、碳水化合物和维生素等营养物质丰富[1],是我国高经济价值水产增养殖品种的代表[2]。目前我国中华绒螯蟹的人工养殖区已从黄海、渤海、东海沿岸和通海江河及其附属湖泊扩大到新疆、内蒙古等地区。大闸蟹养殖已成为我国独具特色的淡水渔业支柱产业之一。由于不同产地蟹的养殖环境及模式等存在差异,一些优质产地所产出的蟹业已形成了知名品牌[3-4],并被赋予了更高的产品品质和商业价值。但随着优质产地、知名品牌大闸蟹的影响力不断扩大,营收价值不断提高,其产业也越来越受到假冒蟹(如“过水蟹”、“洗澡蟹”等)[5]及劣质蟹(如污染蟹等)[6-7]的侵害。因此,研究不同产地大闸蟹外部形态和内部成分的产地差异性特征并有效开发产地鉴别技术,对蟹产业的有序发展意义重大。
在外部形态研究方面几何形态测量学(geometric morphometrics)正起到越来越重要的作用。其被认为是主要基于二维或三维形态结构上符合特定定义的同源地标点及其精密的数据系统,来破解形态差异性及其动态和可视化的学科[8]。该法所选取的地标点是具有明显特征,容易辨识的点;且在后续可视化处理时,还可消除在选取时可能发生位置、取向和比例上非形态变化的干扰[9]。郑朝臣 等[10]首次尝试引入该几何形态测量学分析法,研究了中华绒螯蟹背甲及腹甲结构形态的产地差异性,并发现基于雌、雄蟹背甲形态的产地判别准确率高达100%,可比肩以往通过地理元素“指纹”方式才能达到的判别准确率。近期,一些利用该法研究不同国家和我国不同水系间中华绒螯蟹的背甲形态特征差异的工作也取得了较为满意的判别效果[11-12]。
需要注意的是,上述前人相关研究多聚焦于中华绒螯蟹大型结构(特别是背甲)形态,对蟹外形其他精细小型结构或部位(如步足,含特化的螯足及其分节结构)的形态特征却缺乏关注。这些小型结构或部位应该同样具有受产地影响而发生地理适应性变化的可能。实际上,这种趋势在其他的动物类群上已有发现。王超等[13]在利用几何形态测量法探究了剑尖枪乌贼()角质颚的生长变化规律时发现,不同性别个体的角质颚大小在发育过程中存在一定差异。雌性个体的角质颚大于雄性个体,且角质颚形态随着其个体不断发育呈渐变规律,同时存在异速生长现象;可能与该种乌贼发育过程中的摄食和环境变化有关。李晓慧等[14]采用几何形态学测量法并结合Micro CT扫描技术,对两水系花斑裸鲤()的形态进行了定量分析,特别对两种群背鳍支鳍骨插入椎骨间的相对位置这一具体部位形态特征差进行了显著性检验;结果显示,在具分类学意义的骨骼形态特征上的差异已达显著水平。张秀霞等[15]关于唐鱼群体的研究也发现,不同野生和养殖群体鱼在头部以及眼径大小等精细部位的形态均存在变异性。通过这些变异进行群体产地判别的正确率可高达97.6%,且揭示了唐鱼野生种群和养殖群体之间存在显著的形态差异。这也为研究养殖和野生中华绒螯蟹群体差异性提供了理论基础。上述研究进展显示,水生生物外部精细结构或小型部位的形态特征也应该具有进行生物分类及产地鉴别的潜力。
本研究以长江水系中安徽、江苏7个产地中华绒螯蟹背甲作为研究对象,创新性地选用螯足长节作为精细小型结构或部位的代表,对其产地间的几何形态测量学差异性特征开展了较为详细的研究,以期达到确认除背甲外,中华绒螯蟹其他精细小型结构或部位的形态是否真实存在地理差异性,以及这种差异性是否可以用于替代背甲,来开展蟹非致死、致伤产地溯源手段的开发及应用的目的。
1 材料与方法
1.1 材料
本研究所选用的长江水系中华绒螯蟹样本分别于2017年10月采自长江中游地区的湖南大通湖,2018年11月采自长江下游地区的安徽黄陂湖、女山湖、沱湖,2017年12月采自长江安徽无为江段,2017年11月采自江苏溱湖、石臼湖7个产地(表1)。除无为江段的蟹为长江的野生个体外,其余蟹均为放养或围网养殖于相应湖区的个体(苗种均为购自江苏南通繁育场的长江水系扣蟹,种质资源遗传背景相同)。本研究中所采集各湖区产地的养殖蟹均符合国标GB/T19957—2005[16]所标准定义的原产成蟹(至少从扣蟹养到成蟹这一过程保持在同一水域内)的要求。至采样时,各产地蟹均为商品规格成蟹,已完成全部蜕皮过程[17]。各产地蟹采集后均尽快运回实验室,并活体进行常规测量。
1.2 方法
地标点的选取建立:按照生物学中应用的地标点法分类和选取基本原则来进行。由于中华绒螯蟹螯足指节和掌节部分长有绒毛,难以处理,易使地标点的选取存在误差,且腕节处的特征点较少;故确定以中华绒螯蟹螯足长节[18]为实验对象,选取14个Ⅱ型地标点(即以不同组织间交点为特征的Ⅰ型地标点,以结构中凹陷或凸起点为特征的Ⅱ型地标点和以结构极值点为特征的Ⅲ型地标点)[19](图1)来开展研究。在LED灯光(6W)条件下,利用三脚架设定拍摄固定高度(24 cm)的Nikon COOLPIX P6000数码相机(日本Nikon集团公司),进行蟹背甲拍照。将拍好的照片导入Photoshop软件(13.0版,美国Adobe系统公司)后,再在利用tpsDig2软件中进行[20]地标点的标记,获取地标点的x、y坐标值,建立二维地标点的数据文件。
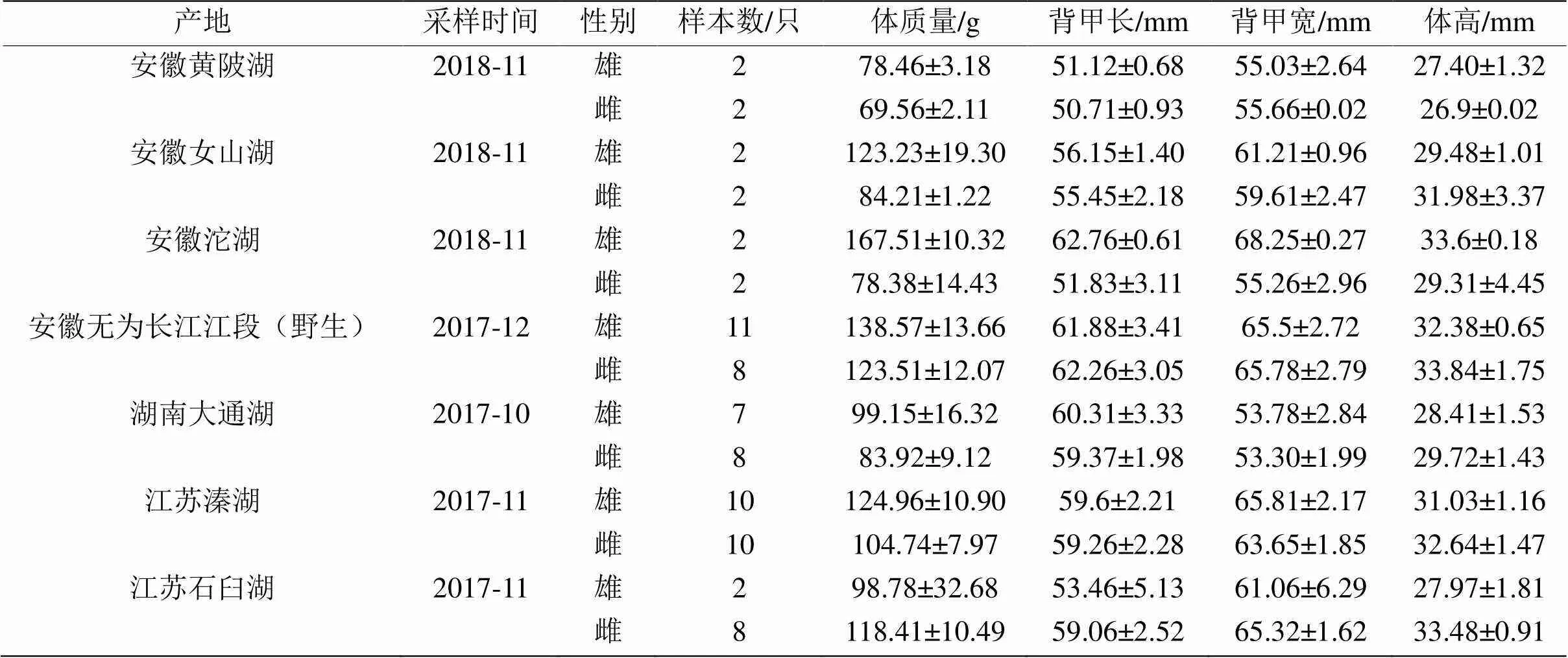
表1 长江水系7个不同产地中华绒螯蟹群体的形态测量信息
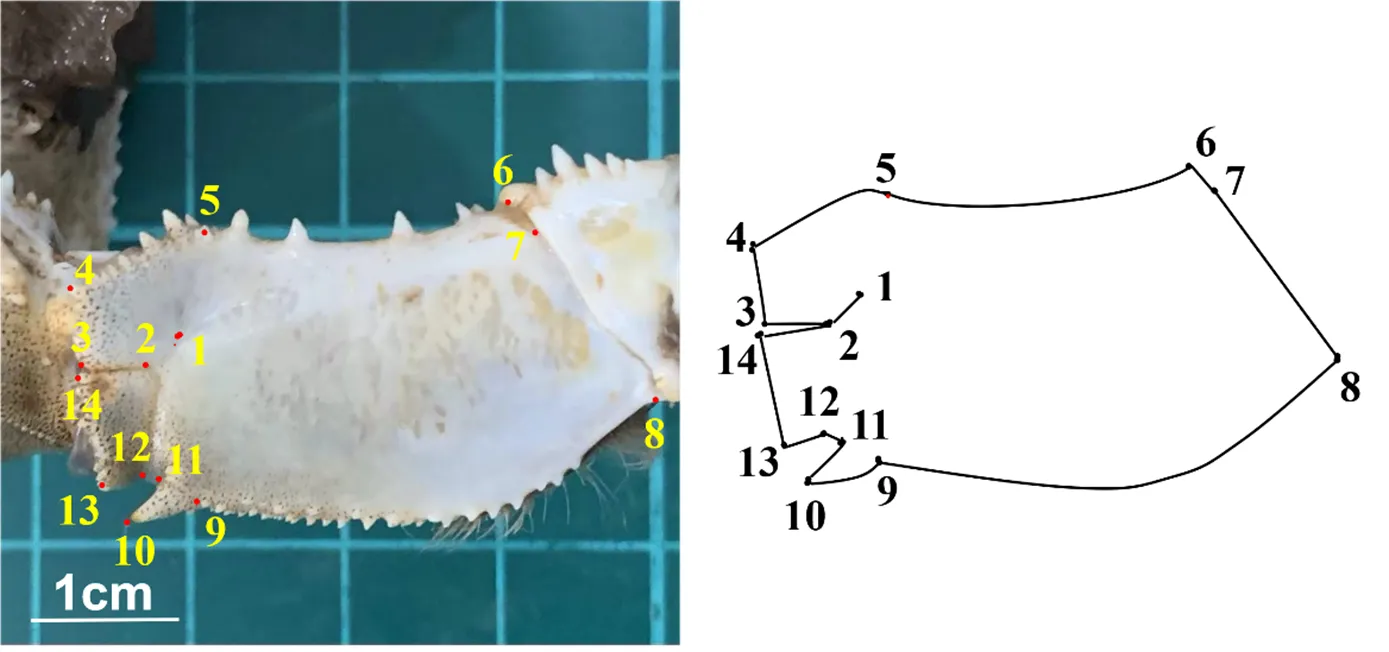
1: 长节内部凹陷点; 2:长节左端分叉点; 3、4、13、14:长节左端顶点;5:内部凹陷与上边缘相交点; 6、8:右边缘上下顶点;7:右边缘与内部突起相交点; 9、11:下边缘与最大侧齿交点;10: 最大侧齿顶点;12:内部凹陷与下边缘相交点。
Figure 1 Location map of landmark points for the cheliped merus of
平均形与薄板样条分析:地标点的原始坐标数据包含了许多非形态变异。利用tpsRelw软件[20]普氏叠加运算来消除上述非形态变异,得到标准化数据并求出平均型(图2),以用于后续的判别分析。依据标准化数据的相对扭曲分析获得14个地标点的贡献率[19]。利用tpsRegr软件进行薄板样条分 析[9,21],绘出各产地中华绒螯蟹网格变形图(图3,图4),进行差异比较。
判别分析:将经tpsRelw软件得到的相对扭曲得分导入Excel表格,采用判别错分概率及风险最小的Bayes方法[22]进行判别分析。将所有自变量纳入SPSS(23.0版)(美国IBM公司)判别函数进行统计分析。
2 结果与分析
2.1 螯足长节形态差异可视化分析
利用tpsRelw软件根据地标点数据文件计算得到螯足长节形态的平均形(图2),再通过绝对扭曲、回归分析和置换检验进行网格化和差异矢量化后的网格图(图3、图4)。
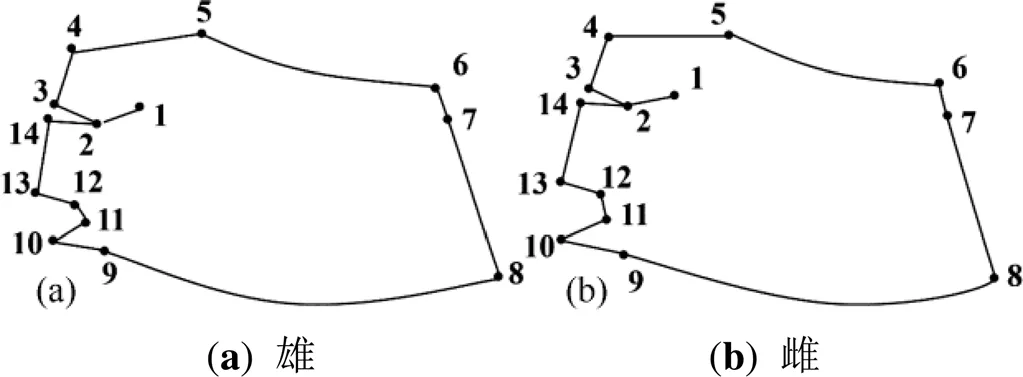
图2 中华绒螯蟹螯足长节平均形
Figure 2 Average shape of the cheliped merus of

图3 不同产地中华绒螯蟹雄蟹螯足长节变异可视化(变异扩大3倍)
Figure 3 Mesh deformation map and variation visualization (variation is 3 times larger) for the cheliped merus of malefrom different geographical origins
相比雄性中华绒螯蟹(图3),安徽黄陂湖蟹、女山湖蟹、沱湖蟹螯足长节与平均形相比长节内部凹陷点(第1地标点)向左下偏移;长节左端分叉点(第2地标点)向左偏移;左端最上顶点(第4地标点)向左下偏移、最下顶点(第13地标点)向右上偏移;右边缘上顶点(第6地标点)向右偏移,下顶点(第8地标点)向右上偏移;右边缘与内部突起相交点(第7地标点)向右下偏移;下边缘与最大侧齿交点(第9、11地标点)及最大侧齿顶点(第10地标点)均向右上方偏移,下边缘最大侧齿较平均形细小。三产地蟹螯足长节形态与平均形的扭曲变化程度为:黄陂湖蟹>女山湖蟹>沱湖蟹。
安徽无为长江江段(野生)蟹螯足长节形态与平均形相比较为接近,没有明显变异扭曲现象。
湖南大通湖蟹螯足长节与平均形相比其内部凹陷点(第1地标点)微向右上偏移。
江苏溱湖蟹、石臼湖蟹螯足长节与平均形相比长节内部凹陷点(第1地标点)向右上偏移;长节左端分叉点(第2地标点)向右偏移;左端最上顶点(第4地标点)向右上偏移、最下顶点(第13地标点)向左下偏移;右边缘上顶点(第6地标点)向左偏移;右边缘与内部突起相交点(第7地标点)向左上偏移;下边缘与最大侧齿交点(第9、11地标点)及最大侧齿顶点(第10地标点)均向左下方偏移,下边缘最大侧齿较平均形粗大;其石臼湖蟹各点偏移程度更大。

图4 不同产地中华绒螯蟹雌性螯足长节网格变形图与变异可视化(变异扩大3倍)
Figure 4 Mesh deformation map and variation visualization (variation is 3 times larger) for the cheliped merus of femalefrom different geographical origins
相比雌性中华绒螯蟹(图4),安徽黄陂湖蟹、女山湖蟹、沱湖蟹螯足长节与平均形相比长节内部凹陷点(第1地标点)向左下偏移;长节左端分叉点(第2地标点)向右下偏移;左端最下顶点(第13地标点)向右下偏移;右边缘上顶点(第6地标点)向右下偏移;下边缘与最大侧齿上交点(第11地标点)向右上偏移,下边缘与最大侧齿下交点(第9地标点)及最大侧齿顶点(第10地标点)均向左上方偏移,下边缘最大侧齿较平均形尖细。三产地蟹螯足长节形态与平均形比较的扭曲变化程度为:黄陂湖蟹>女山湖蟹>沱湖蟹。
安徽无为长江江段(野生)蟹螯足长节形态与平均形相比较为接近,没有明显变异扭曲现象。湖南大通湖蟹螯足长节与平均形相比其内部凹陷点(第1地标点)微向右偏移。
江苏溱湖蟹、石臼湖蟹螯足长节与平均形相比长节内部凹陷点(第1地标点)向右上偏移;左端最下顶点(第13地标点)向右上偏移;右边缘上顶点(第6地标点)向左上偏移;下边缘与最大侧齿上交点(第11地标点)向左下偏移,下边缘与最大侧齿下交点(第9地标点)及最大侧齿顶点(第10地标点)均向右下方偏移,下边缘最大侧齿较平均形粗短;其石臼湖蟹各点偏移程度更大。
2.2 螯足长节形态差异判别分析
对雄性中华绒螯蟹的螯足长节进行产地判别,准确率均为100%,判别准确率高;对雌性中华绒螯蟹的螯足长节进行产地判别,准确率也均为100%;判别准确率高(表2)。由图5中华绒螯蟹的判别分析散点图分析,中华绒螯蟹螯足形态地标点法的应用结果在视图上也能明显且有效的显示出判别结果良好。
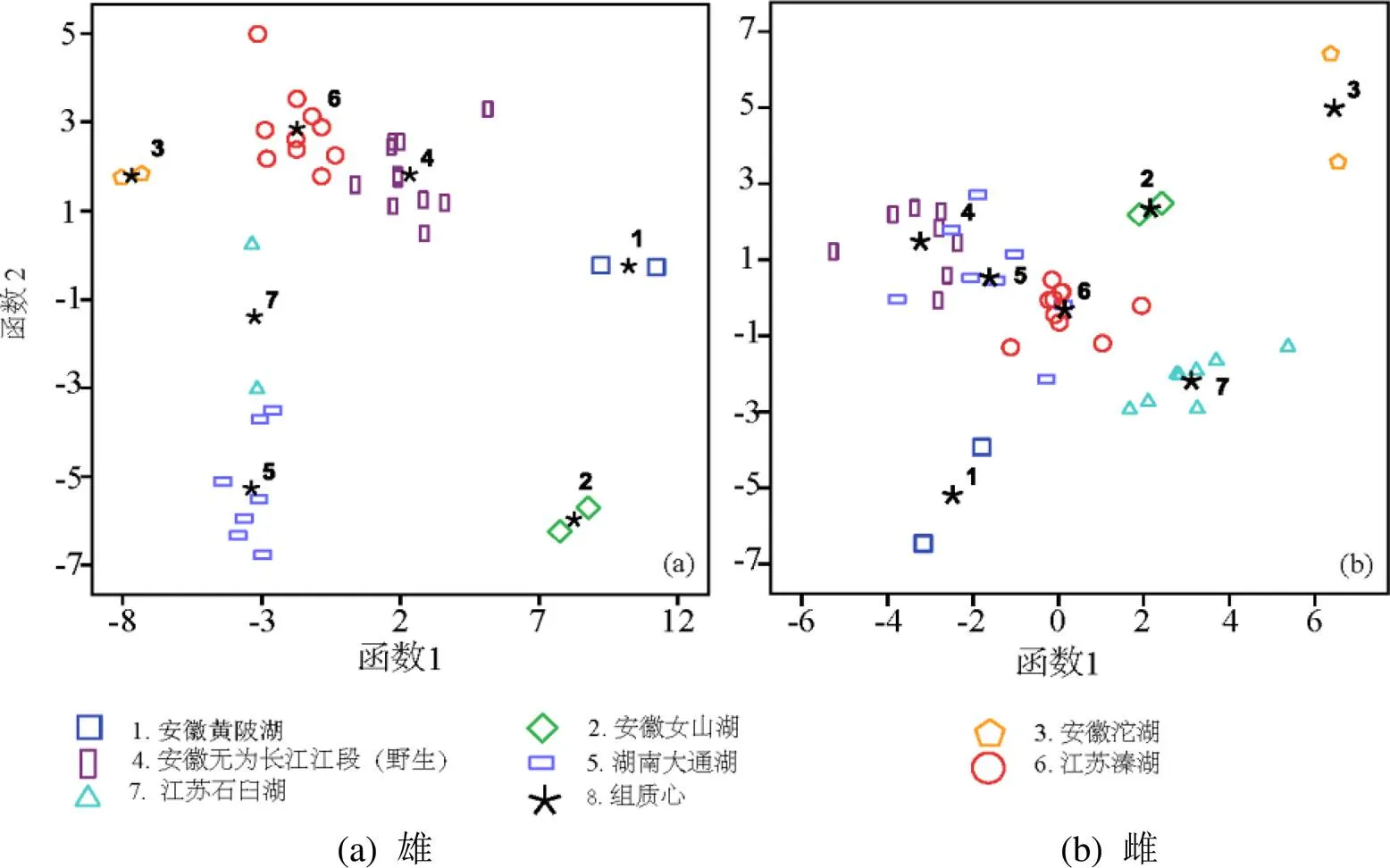
图5 不同产地中华绒螯蟹几何形态学特征的判别分析散点图,)
Figure 5 Scatter plot of discriminant analysis on geometric morphometrics offrom different geographical origins

表2 基于螯足长节形态的7产地雌雄中华绒螯蟹判别分析
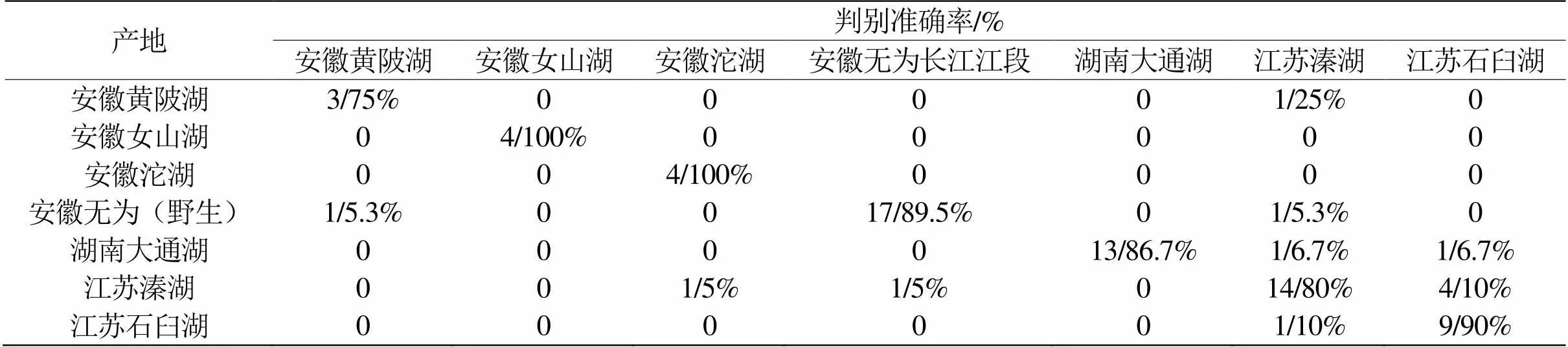
表3 基于螯足长节形态的7产地中华绒螯蟹判别分析
将每个产地的雌、雄蟹作为一个整体,进行仅7个产地间中华绒螯蟹螯足长节几何形态的判别分析,准确率为84.2%(表3)。由此可见,性别的差异对产地差异的准确判别会产生极大的干扰。
3 讨论与结论
在对7个产地中华绒螯蟹螯足地标点数据进行的地标点法分析中,雌雄蟹贡献率最高的地标点均为第3和11、13、14地标点,均共解释了89.28%的形态差异,其余10个地标点解释了10.72%的形态差异。本研究中选取的雌雄中华绒螯蟹螯足长节地标点的切空间距离和普氏距离的回归系数分别为0.999 351和0.999 172,均接近于1,这说明选取的地标点是有效的。本研究利用基于地标点的几何形态测量法来掌握同属长江水系7个不同产地养殖和野生中华绒螯蟹形态差异,取得了满意的结果。结果显示,与平均形比较,除无为野生蟹无明显的变化外,安徽省3产地蟹长节形态有同趋势变化,长节左下端向内缩小,长节内部凹陷点向左下端靠近;其余3个产地蟹长节形态有同趋势变化,长节左下端向外扩展,长节内部凹陷点向右上方向偏移。雌、雄蟹个体产地间的判别准确率均可达100%。由于包括大闸蟹在内的水产品生物的分布地及生长环境不同,其外部形态等方面可出现地理性的差异[23-27]。物种形态学变异可受遗传因素和环境因素等的双重影响,在敞水和浅水环境中养殖湖沼型和底栖型太阳鱼(),比较其遗传变异(genetic differentiation)和表型可塑性(phenotypic plasticity)对鱼形态变化权重时发现,养殖环境差所导致的表型形态可塑性变异的贡献率达53%,远大于由遗传分化所造成14%的贡献率[26];因此,更需重视养殖环境差异导致的形态学变异。针对中华绒螯蟹的大型外部结构背甲(头胸甲)的研究较为支持上述观点[26]。该近六边形钙质结构硬组织的边缘总体上可分为前缘、眼缘、前侧缘、后侧缘和后缘五部分。前缘正中为额部,有4枚齿突,被称为额齿。额齿中间的凹陷,以中央的一个为最深。左右前侧缘各有4个锐齿,被称为侧齿[28]。背甲后侧缘斜向内侧,后缘与腹部交界,比较平直。背甲中部有呈M型花纹[29]。Xue等[30]通过对1周年养殖期间阳澄湖大闸蟹背甲形态动态监测认为,背甲变化最大的部位主要在第3、4侧齿和M型花纹,影响背甲的可塑性因素主要因素应该为阳澄湖养殖环境的生物(如敌害物种)及非生物(如底质、水流、水质、养殖方式)因素。虽然造成本研究蟹精细结构部位螯足长节产地间差异的原因尚不知晓,但养殖环境差异的权重应该很大。这点尚待开展下一步的研究来加以证实。
与Xue等[5]所发现雌、雄蟹背甲存在显著差异的情况类似,本研究蟹的螯足长节形态之间也存在性别上的差异,而这种差异对产地差异的准确判别会产生明显干扰;因此,需要将雌、雄蟹分开进行多元统计分析。同样,潘鹏亮等[31]利用几何形态测量学技术对同属不同种金龟子的区分鉴别研究中发现,代表体躯背面特征的前胸背板、中胸小盾片、鞘翅等的长宽距离可以区分东北大黑鳃金龟性别,足部特征可以区分华北大黑鳃金龟性别,金龟子的翅脉特征可以区分所研究的各种金龟子的性别。来益同等[32]对大瓶螺()体色、性别与贝壳形态之间关系的研究发现,不同体色个体的贝壳形态存在着显著差异,而不同体色的贝壳差异均不及相同体色不同性别个体间贝壳的差异。这表明,贝壳形态的差异也存在性别上的区别。前人的研究表明了生物体的某些形态特征具有性二型,进一步佐证了本研究的结果,即中华绒螯蟹的背甲等大型结构及螯足长节等精细小型结构或部位形态应该普遍存在性别上的差异性。
虽然作为初步尝试,本研究中个别产地(如安徽黄陂湖、沱湖、女山湖和江苏石臼湖等)蟹样本量较小(如=4,雌雄各半),但并未掩盖螯足长节形态明显的产地差异(图5)。在前人研究中,小样本数量的判别结果依旧可以接受并具有学术价值。张秀秀等[33]在秦岭地区中华蜜蜂旬阳坝(=3)等5个地理种群翅形态差异的研究中,应用几何形态测量学方法发现了中华蜜蜂前翅和后翅形态在种群间变化规律不同,前翅形态变化更显著。在下一步的研究中,仍需加大样本数量来对螯足长节等精细小型结构形态的性别和产地差异性特征进行更深入地验证。
综上所述,本研究创新性地选用螯足长节作为产地鉴别的依据,发现了长江中下游7个产地的中华绒螯蟹螯足长节形态存在明显的产地差异,且确认了基于外部小型结构螯足长节地标点的几何形态测量学法对产地间雌、雄蟹个体的判别准确率均可达100%,更初步证实了中华绒螯蟹除背甲形态存在产地差异性外,其他精细结构或小型部位也存在因环境而发生的适应性变化。这为后续利用水产生物精细结构的形态来非致死致伤地研发产地溯源的手段提供新思路。但当前研究总体样本数量基数较小,其结果的实用性还有待进一步验证。在后续的研究中,将扩大样本数量及产地范围,进一步确证中华绒螯蟹螯足形态在产地溯源方面的应用价值。
[1] 李伟中.阳澄湖大闸蟹的经济与文化价值探析[J].扬州大学烹饪学报,2006,23(3):6-10.
[2] 宋黎黎. 越冬暂养对中华绒螯蟹生长、生理及品质的影响[D].上海:上海海洋大学,2021.
[3] 姬皓禹,赵伟进,白春雨.兴化大闸蟹产业发展现状及前景展望[J].江西水产科技,2021(6):41-43,45.
[4] 李微.江苏昆山大闸蟹全产业链发展现状与展望[J].科学养鱼,2021(12):1-2.
[5] 薛竣仁,姜涛,陈修报,等.阳澄湖原产及“洗澡”中华绒螯蟹背甲的几何形态分析[J].水生生物学报, 2020, 44(3): 587-594.
[6] 王婕,刘源.淮河沉积物中镉的历史变迁及污染评价研究[J].四川轻化工大学学报(自然科学版), 2021, 34(6): 47-54.
[7] LIU X L, WANG Q, HE J, et al. Occurrence, speciation analysis and health risk assessment of arsenic in Chinese mitten crabs () collected from China[J]. J Food Compos Anal,2020,94:103647.
[8] COOKE S B, TERHUNE C E. Form, function, and geometric morphometrics[J]. Anat Rec,2015,298(1):5-28
[9] BOOKSTEIN F L. Morphometric tools for landmark data: geometry and biology[M]. Cambridge: Cambridge University Press,1997:55-87.
[10] 郑朝臣,姜涛,骆仁军,等.不同产地中华绒螯蟹形态差异的地标点法分析[J].水产学报,2017,41(12):1896-1907.
[11] 姜晓东,成永旭,潘建林,等.中国长江与荷兰野生中华绒螯蟹的头胸甲形态特征比较[J].淡水渔业, 2020, 50(1): 38-43.
[12] ZHENG C C, JIANG T, LUO R J, et al. Geometric morphometric analysis of the Chinese mitten crab: a potential approach for geographical origin authentication[J]. N Am N J Fish Manag, 2021, 41(4): 891-903.
[13] 王超,方舟,陈新军.基于几何形态测量法的剑尖枪乌贼角质颚形态变化研究[J].渔业科学进展, 2023, 44(1): 58-69.
[14] 李晓慧,汤永涛,田菲,等.几何形态测量法结合Micro CT扫描对两水系花斑裸鲤的形态分析[J].水生生物学报, 2020, 44(4): 853-861.
[15] 张秀霞,朱巧莹,赵俊.利用几何形态测量学方法分析唐鱼群体的形态变异[J].水产学报,2017,41(9):1365-1373.
[16] 全国原产地域产品标准化工作组.地理标志产品阳澄湖大闸蟹:GB/T19957-2005[S].北京:中国质检出版社, 2005.
[17] XUE J R, JIANG T, CHEN X B, et al. Multi-mineral element profiles in genuine and “bathing” cultured Chinese mitten crabs () in Yangcheng Lake, China[J]. Fishes,2022,7(1):11.
[18] LAI J C Y, NG P K L, DAVIE P J F. A revision of the(Linnaeus, 1758) species complex (Crustacea: Brachyura: Portunidae), with the recognition. of four species[J].Raffles B Zool, 2010, 58(2): 199-237.
[19] ROHLF F J. Relative warp analysis and an example of its applications to mosquito wings[M]//MARCUS L F, BELLO E, GARCIA-VALDECASAS A. Contributions to Morphometrics. Madrid :Museo Nacional de Ciencias Naturales, 1993, 131-159.
[20] ROHLF F J. TpsRelw (version 1.46) [EB/OL]. (2021-01- 24)[2022-11-14].http://sbmorphometrics.org/index.html.
[21] 陈新军,方舟,苏杭,等.几何形态测量学在水生动物中的应用及其进展[J]. 水产学报, 2013,37(12):1873-1885.
[22] 王梦瑶,孙伟琪,陈冰冰,等.贝叶斯判别分析在财务预警中的应用[J].品牌与标准化,2021(1):59-61,64.
[23] MACLEAN J A, EVANS D O. The stock concept, discreteness of fish stocks, and fisheries management [J]. Can J Fish Aquat Sci,1981,38(12):1889-1898.
[24] 郑海波.中华绒螯蟹的品质分析与比较[D].无锡:江南大学, 2008.
[25] 谢辉,尹明雨,张玉非,等.不同产地雌性中华绒螯蟹感官品质与滋味品质的差异性[J].食品与发酵工业, 2021, 47(6): 114-120, 126.
[26] ROBINSON B W, WILSON D S. Genetic variation and phenotypic plasticity in a trophically polymorphic population of pumpkinseed sunfish () [J]. Evol Ecol, 1996,10(6):631-652.
[27] 郑朝臣,姜涛,骆仁军,等.不同产地中华绒螯蟹形态差异的地标点法分析[J].水产学报,2017,41(12):1896-1907.
[28] 于洪贤,周玲玲,于海波.黑龙江省大水面河蟹综合养殖技术研究[J].黑龙江水产,2003(1):28-33.
[29] 张列士,姜治忠,李军.日本绒螯蟹与不同水系中华绒螯蟹的形态比较[J].上海水产大学学报, 2002, 11(2): 110- 113.
[30] XUE J R., LIU H B, JIANG T, et al. Shape variation in the carapace of Chinese mitten crabs (H. Milne Edwards, 1853) in Yangcheng Lake during the year-long culture period[J]. Eur Zool J, 2022, 89(1): 217-228.
[31] 潘鹏亮,张方梅,洪枫,等.几何形态测量学在齿爪鳃金龟属成虫分类鉴定中的应用研究[M]//魏洪义,曾菊平,夏斌.华中昆虫研究(第十六卷), 杨凌:西北农林科技大学出版社, 2020, 336-337.
[32] 来益同,沈华,张奕祥,等.2个纯合体色品系大瓶螺的几何形态测量学研究[J].水生态学杂志,2016,37(2):88-95.
[33] 张秀秀,管德龙,覃凯华,等.秦岭地区中华蜜蜂5个地理种群翅形态差异的几何形态测量学研究[J].陕西师范大学学报(自然科学版),2017,45(2):71-74.
Geometric morphometric study on the origin difference of cheliped merus of
TANG Jing1, XUE Junren1, LIU Hongbo2, JIANG Tao2, CHEN Xiubao2, YANG Jian1,2
(1. Wuxi Fisheries College, Nanjing Agricultural University, Wuxi 214081; 2. Laboratory of Fishery Microchemistry, Freshwater Fisheries Research Center, Chinese Academy of Fishery Sciences, Wuxi 214081)
For revealing the origin difference on the morphological characteristics for a fine structure of cheliped merus, geometric morphological analysis was applied for the first time to comparatively investigate withcrabs from seven different well-known geographical origins in Hunan Province, Anhui Province and Jiangsu Province in the middle and lower reaches of the Yangtze River. Based on landmark selection principle of corresponding computer software image processing, a 14-landmark system was established, and then, the average cheliped merus shape of the crabs from the seven origins was obtained by standardized operation. The morphological variation vector was further visualized by thin plate spline analysis and mesh deformation. In comparison with the average cheliped merus shape, although no significant variation was found for that of Wuwei wild population, significant differences were documented for those from other six origins, i.e. the morphology of the merus of crabs from three origins in Anhui Province showed the same trend. The left lower end of the merus was narrowed inward, and the internal depression point of the merus was close to the left lower end. The morphology of the merus of crabs from remaining 3 producing origins of Hunan and Jiangsu Provinces showed the same trend. The left lower end of the merus extended outward, and the internal depression points of the merus shifted to the right upper direction. The discriminant analysis of the above standardized data showed that the discriminant accuracy of female and male crabs could reach 100%. The above results innovatively confirm that the landmark approach has great potential for nonlethal and non-wounded determination of the origin difference and traceability ofcrabs based on geometric morphology measurement by either large structure of carapace or fine structure/small part of cheliped merus.
; geometric morphometrics; landmark; the cheliped merus; origin traceability
S968.25
A
1672-352X (2023)05-0802-07
10.13610/j.cnki.1672-352x.20231030.005
2023-10-31 10:04:31
[URL] https://link.cnki.net/urlid/34.1162.S.20231030.0948.010
2022-09-09
昆山市阳澄湖大闸蟹产业研究院项目(HX2021509300),中央级公益性科研院所基本科研业务费专项资金项目(2021XT0704),国家自然科学基金面上项目(31772850)共同资助。
唐 静,硕士研究生。E-mail:2020813061@stu.njau.edu.cn
杨 健,博士,研究员,博士生导师。E-mail:jiany@ffrc.cn
猜你喜欢
小学生学习指导(低年级)(2021年12期)2021-12-31 08:09:26
辽宁省博物馆馆刊(2021年0期)2021-07-23 07:27:08
小学生学习指导(低年级)(2020年9期)2020-11-09 09:11:22
中国外汇(2019年22期)2019-05-21 03:14:56
意林·全彩Color(2018年9期)2018-10-12 01:06:54
中成药(2018年8期)2018-08-29 01:28:16
东方教育(2017年11期)2017-08-02 15:02:00
东方教育(2017年11期)2017-08-02 12:17:28
东方教育(2017年11期)2017-08-02 06:22:44
东方教育(2017年11期)2017-08-02 00:08:49
