
利用CRISPR/Cas9技术改良三系杂交稻恢复系外观品质
2023-11-22 04:11高利军李经成周维永邓国富潘英华卿冬进陈韦韦戴高兴
西南农业学报 2023年9期
黄 娟,高利军,李经成,周维永,邓国富,潘英华,卿冬进,陈韦韦,伍 豪,戴高兴
(1. 广西农业科学院水稻研究所/广西水稻遗传育种重点实验室,南宁 530007; 2. 广西作物遗传改良生物技术重点开放实验室,南宁 530007;3. 广西农业科学院, 南宁 530007)
【研究意义】水稻粒型不仅影响稻谷产量,而且影响稻米品质,对水稻产量和品质形成均具有重要作用[1-2]。粒型包括粒长、粒宽和粒厚,与水稻的垩白度、垩白粒率和透明度等外观品质关系密切[3]。随着人们消费水平的提高,对稻米品质的要求也越来越高,特别是高端米市场对稻米外观品质要求严格,粒型细长、无心腹白和晶莹剔透是优质稻米的典型外观特征,因此,丝苗型稻米(糙米长≥6.5 mm,糙米长宽比≥3.5,T/GDSMM 001—2019)在市场上广受欢迎[4]。杂交稻品种的杂种优势能提高稻谷产量,但其品质提高上一直落后于常规稻[5]。杂交稻的品质由恢复系与不育系共同决定[6-7],当前恢复系选育仍以高产、恢复力强、配合力高和抗性强等为主要标准,存在垩白度和垩白粒率高的现象,利于杂交稻外观品质提高[8-9]。三系杂交稻的保持系、不育系和恢复系之间具有严格的恢保关系,传统育种过程通过杂交方式引入外源基因,有降低恢复系恢复强度的风险,极大降低了育种效率[10]。因此,借助基因编辑手段,加强亲本的粒型选择,是提高三系杂交稻外观品质的有效途径。【前人研究进展】水稻的外观品质包括米粒长、米粒宽、米粒长宽比、垩白粒率、垩白度和透明度等。垩白为稻米胚乳中白色不透明部分,是影响稻米品质的重要因素,决定着稻米的商品价值[11-12]。研究结果显示,垩白率和垩白度与粒宽和粒厚呈显著正相关,与粒长和长宽比呈极显著负相关[3],因此,粒型调控对垩白的形成有不可忽略的影响[13-18]。目前已有超过60个粒型相关基因被克隆[19-20],这些克隆的基因中,GS3和TGW3基因负责调控水稻籽粒的粒长和粒重,GW8基因正向调控水稻粒宽,是生产上重要的粒型调控基因[15,21-22]。传统杂交育种方式周期长,效率低下;粒型作为数量性状,增加了品种选育难度[23]。基因编辑技术可定向改造目的基因,为水稻的品种改良带来了契机[24]。Zhou等[25]利用CRISPR/Cas9技术,在籼稻温敏核不育系龙科638S中同时编辑3个广谱抗稻瘟病基因Bsr-d1、Pi21和ERF922,在T1代获得了无转基因纯合的单基因或三基因突变体,与野生型相比,所有突变体都对稻瘟病表现出更强抗性;Hui等[26]利用CRISPR/Cas9技术,在粳稻宁粳1号和籼稻黄华占中获得具有香味的BADH2等位基因;徐善斌等[27]利用CRISPR/Cas9技术在龙粳11中同时敲除GS3及GS9基因,T2代突变体粒长增加26.43%~27.01%,单株产量增加10.82%~12.11%,千粒重增加 18.34%~41.36%;韩政宏等[28]在多个粳稻品种中同时敲除GS3和GS9基因,T2代获得了籽粒细长和千粒重显著增加的突变体;赵春芳等[29]在南粳5055中敲除GS3和qGL3基因,获得了一系列大粒水稻新种质;谷玉娟等[30]创制了仅编码GS3基因 C 端功能结构域的突变类型,该类型的籽粒长度和千粒重有所增加;Wang等[31]利用CRISPR/Cas9对GS2基因的miR396识别位点进行编辑,突变体GS2E植株的千粒重和单株产量分别提高了23.5%和10.4%;Chen等[20]在日本晴中单独敲除GS3基因,突变单株的千粒重无显著差异,GS3及GL3.1(TGW3)的同时敲除使千粒重得到显著提高;Huang[32]等在籼稻保持系Mei1B中敲除GS3基因,gs3突变体及其测配组合(gs3/gs3)的千粒重比野生型以及杂合状态均得到显著提高。基因编辑技术在水稻产量、品质和抗性等改良上均取得显著效果。【本研究切入点】广西农业科学院水稻研究所杂交水稻遗传育种课题组自主选育的籼稻广亲和强恢复系桂恢582 (GH582)配组的超级稻组合桂两优2号为早晚兼用型超级稻品种,能适应华南稻区复杂的气候,早晚稻种植均能高产[33]。该品种的恢复系籽粒长约8.40 mm,宽约2.81 mm,籽粒短圆,在外观品质上与优质米有较大差距。针对恢复系的育种需求,本研究从粒型改良出发,快速获得外观品质提升的新恢复系。【拟解决的关键问题】利用CRISPR/Cas9技术,在GH582中定点敲除3个重要粒型调控基因GS3、TGW3和GW8,降低其籽粒宽度,提高籽粒长度,为快速提高三系杂交水稻的外观品质育种提供参考依据。
1 材料与方法
1.1 试验材料
供试材料为广西农业科学院水稻研究所水稻遗传育种课题组自主选育的籼稻广亲和强恢复系桂恢582 (GH582)。
1.2 试验方法
1.2.1 靶标设计 在粒型改良提高水稻品质的育种理念支持下,将增加供试品种粒长的同时减少籽粒宽度作为改良方案,选择粒长基因GS3和TGW3、粒宽基因GW8为目标基因。在NCBI数据库上查找序列并设计靶标,GS3基因的靶标为第一外显子上的靶标GS3-y1 cctcgaggaatccgatctcgcgg[32];GW8基因的靶标设计在第3外显子GW8-B1 agaccggaggcaagctggacagg;TGW3基因的靶标则设计在第1外显子TGW3-S1 gaatccatgtcccggccgagagg。
1.2.2 中间载体构建 根据靶标位点序列,分别合成带Eco31I酶切位点的引物(表1),然后将正向引物与反向引物等量混合,进行变性退火。反应程序:95 ℃变性10 min,55 ℃退火10 min,14 ℃延伸5 min。将获得携带Eco31I酶切位点的gRNA片段利用酶切连接方法分别连接到空载体上,操作程序:配置酶切连接体系10.0 μL,其中gRNA片段2.0 μL,空载体1.5 μL,Eco31I酶0.5 μL,T4-ligase 0.5 μL,T4-buffer 1.0 μL,H2O 4.5 μL,将配置好的体系放置于37 ℃培养箱2 h。然后利用酶切连接方法将3个构建的载体中包含编辑元件和双元载体部分进行重组,获得重组质粒CRISPR-Cas9-GS3-GW8-TGW3(图1)。
1.2.3 重组质粒转化 利用载体引物Yl-R+和Pbw2-(表1)对重组质粒中1100 bp的编辑元件进行PCR扩增及测序验证,确认无误后转入农杆菌EHA105中[34],并通过农杆菌介导的方法转化到水稻恢复系GH582的愈伤组织,用潮霉素筛选获得转基因T0代植株。
1.2.4 阳性植株靶位点突变及粒型变异分析 利用抗潮霉素引物Hyg-F/Hyg-R筛选阳性植株。以引物GS3-F1/R1、GW8-F2/R2和TGW3-F/R检测GH582材料获得的阳性植株的突变位点序列。将获得的阳性水稻苗与同时期野生型材料进行种植,成熟期采集籽粒,每个单株挑选10粒饱满度一致的籽粒,置于万深考种仪测量其长度,同时与野生型籽粒进行比对。
1.2.5 粒型突变新恢复系的获得及农艺性状考察 由于对恢复系GH582进行了多个基因、多个位点的编辑,T0代突变纯合单株的选择难度增大,可将T0代单株种子收获后扩大种植数量,在T1代或T2代选择突变位点纯合无标记基因且符合预期目标的单株鉴定为新恢复系。考察粒型突变新不育系的农艺性状,考察内容包括株高、有效分蘖、穗长、籽粒长、籽粒宽、籽粒长宽比、穗粒数、实粒数、千粒重、结实率、单株重量等。

插入片段区域包括U3启动子激活的具有敲除靶点的sgRNA表达盒,泛素启动子激活的Cas9基因和35S启动子激活的潮霉素磷酸转移酶基因。LB:T-DNA左边缘序列;RB:T-DNA右边缘序列。The inserted fragment region included sgRNA expression cassettes with knockout targets activated by U3 promoter, Cas9 gene activated by ubiquitin promoter and hygromycin phosphotransferase gene activated by 35S promoter. LB: T-DNA left border sequence; RB: T-DNA right border sequence.图1 重组质粒示意图Fig.1 Schematic map of the recombinant plasmid
1.2.6 粒型突变新恢复系的米质分析 对鉴定的新恢复系进行米质分析,包括整精米率、米粒长、米粒宽、米粒长宽比、透明度、垩白度、垩白粒率、胶稠度、碱硝值和直链淀粉含量等。
1.3 统计分析
利用GraphPad Prism 8对试验数据进行分析。
2 结果与分析
2.1 恢复系GH582阳性植株筛选及突变分析
潮霉素基因引物筛选获得供试材料的T0代阳性植株28株。由于GH582材料突变位点较多,在T0代较难找到多个位点都纯合且粒型符合需求的单株,将T0代收获的28个单株的籽粒按照10株/行×2行播种成为T1代植株,并用引物Hyg-F/R检测各单株的标记基因状况。选择无标记基因、有籽粒长和籽粒宽显著变化的80个单株进行突变位点测序分析。剔除测序结果为杂合状态的单株,获得突变稳定的21个单株进行基因突变分析和粒型检测。
2.2 基因突变类型分析
21个单株中GS3发生突变的类型为单碱基插入(A、C、T、A),GW8发生突变的类型为单碱基插入(A、T)及单碱基缺失,TGW3发生突变的类型为单碱基插入(G)、单碱基缺失和17个碱基缺失(图2)。这些突变类型都使得终止密码子提前出现,形成截断蛋白,导致籽粒变窄或变长。

靶标序列用蓝色标注,碱基插入用红色小写字母表示,碱基缺失用蓝色连字符表示。The targeted sequence was highlighted in blue, mutations with insertion were marked with red lowercase letters and the deleted sequences were presented by blue hyphens.图2 突变基因的测序分析Fig.2 Sequencing analysis of mutant genes
2.3 粒型基因突变组合分析
对GH582多个不同粒型突变单株的3个突变基因位点进行测序分析,结合粒型突变结果比对,最终得到3种基因型的4种不同突变组合,分别为GW8tgw3GS3、GW8TGW3gs3、gw8TGW3gs3和gw8tgw3gs3,对应的籽粒长和籽粒宽表型见图3。
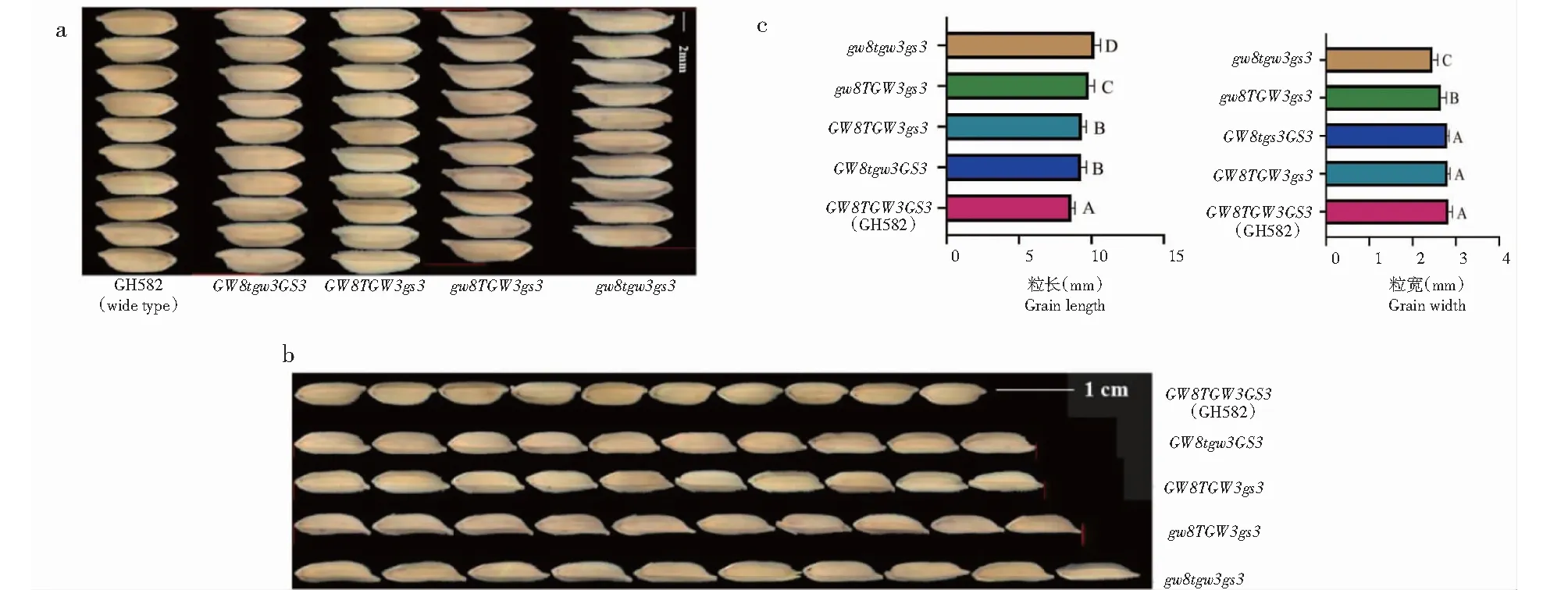
不同大写字母表示差异显著(P<0.05)。Different uppercase letters indicated significant difference(P<0.05).图3 GH582不同突变体籽粒长和籽粒宽差异比较Fig.3 Comparison of grain length and width of different mutants in GH582
由粒型突变及显著性差异分析可知,gs3和tgw3基因的单独突变(GW8TGW3gs3、GW8tgw3GS3)都显著增加了籽粒长(P<0.05,下同),且对籽粒长的贡献相当,而对籽粒宽无显著影响(P>0.05,下同);籽粒宽的显著下降主要来自gw8突变;相比gs3的单独突变,gw8和gs3同时突变(gw8TGW3gs3)使籽粒长度显著增加;三者同时突变(gw8tgw3gs3)时,籽粒长相比gw8和gs3同时突变(gw8TGW3gs3)显著增加,籽粒宽也显著下降。3个粒型基因对籽粒长的增加有叠加效应,在gw8突变的情况下对籽粒宽的影响也有一定的叠加效应。
2.4 粒型突变新恢复系选育及农艺性状测定
综合各突变类型的籽粒突变状态及植株整体表现,3基因突变体gw8tgw3gs3籽粒显著变窄变长,最符合育种目标要求,在T2代选择无标记基因的突变单株种子收获扩大繁殖,暂命名为恢复系GH582E。农艺性状统计显示,GH582E相较GH582,籽粒长度由8.62 mm增加至10.21 mm,增加18.4%,粒长极显著增加(P<0.01,下同);籽粒宽度由2.82 mm下降至2.45 mm,下降13.1%,籽粒宽极显著下降;籽粒长宽比由3.06增加至4.16,增加35.9%(图4)。除粒型变化外,其他农艺性状无显著差异,特别是千粒重、单株重等影响产量的性状无显著差异,说明3个基因的同时突变仅对恢复系GH582的粒型性状产生重要影响。
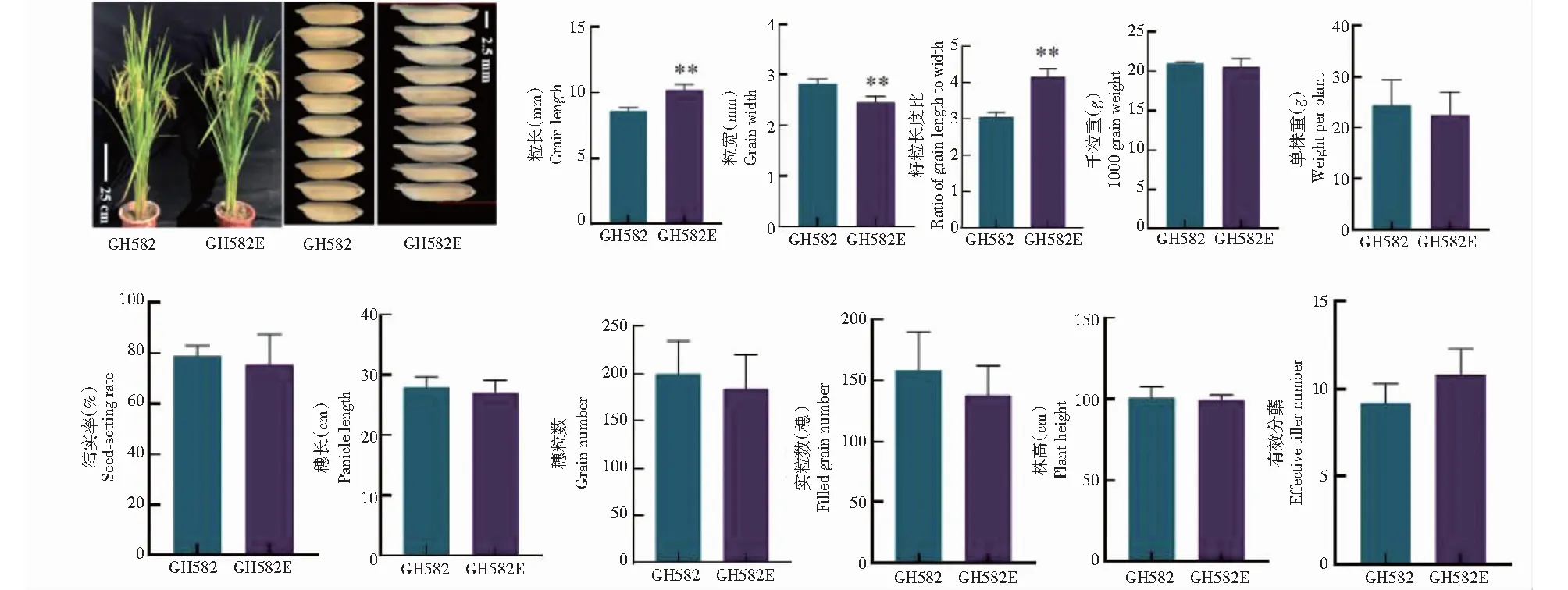
**表示各处理间差异极显著(P<0.01)。** indicated extremely significant difference among treatments(P<0.01).图4 粒型突变新恢复系GH582E与GH582的主要农艺性状对比(n=10)Fig.4 Comparison of main agronomic traits between the new restorer line GH582E and GH582(n=10)
2.5 粒型突变新恢复系的米质分析
米质分析结果(图5)显示,GH582E (gw8tgw3gs3) 相较GH582,米粒长从5.97 mm增加到6.46 mm,增加8.2%,米粒宽从2.37 mm减少到1.99 mm,减少16.0%,米粒长比从2.52增加到3.25,增加29.0%,变化均达极显著水平。透明度数值由4降低2,垩白粒率和垩白度极显著下降,整精米率由于米粒的增加而有所下降,其他米质相关性状没有明显变化,外观品质得到明显提升。由于外观品质的提升,该品种米质由原来的等外米提高到部颁优质米标准3级以上,商品价值得到提升。
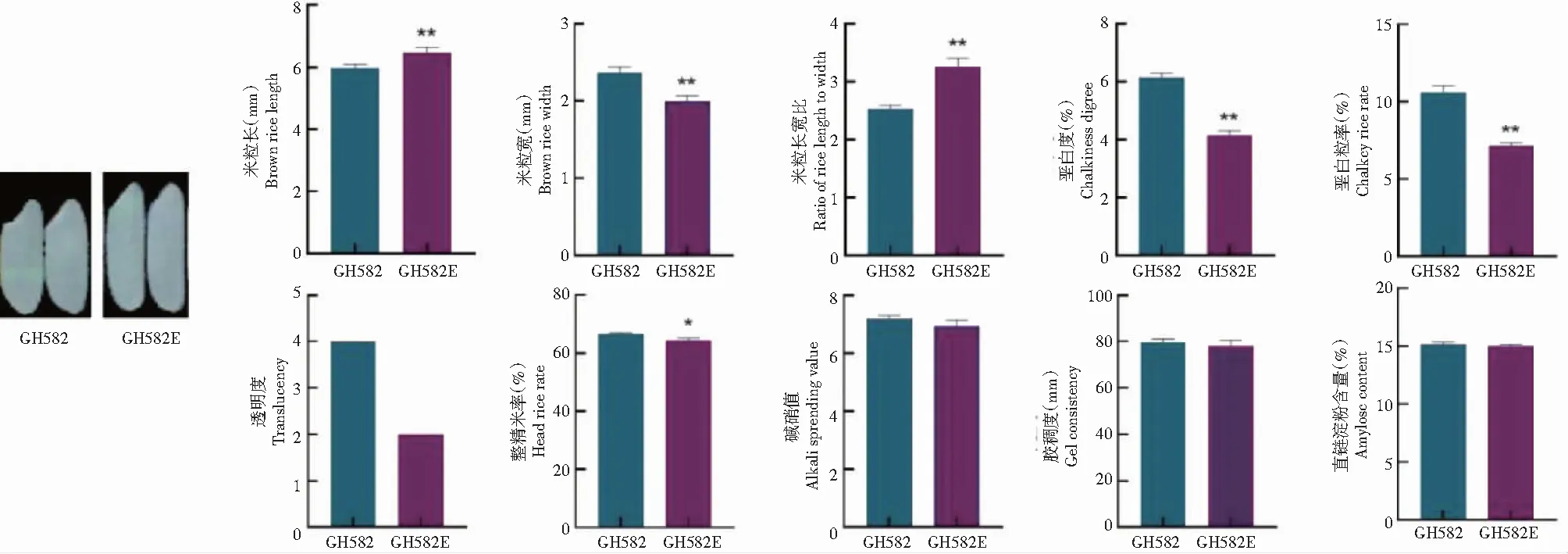
米粒长、米粒宽及米粒长宽比,n=10,其他性状n=3。*表示各处理间差异显著(P<0.05), **表示各处理间差异极显著(P<0.01)。n=10 for brown rice length, brown rice width and ratio of rice length to width and n=3 for the other traits.* indicated significant difference among treatments(P<0.05),and ** indicated extremely significant difference among treatments(P<0.01).图5 粒型突变新恢复系CH582和CH582E的米质分析Fig.5 Analysis of rice quality of new restorer line CH582 and CH582E
3 讨 论
传统水稻品种培育主要利用基因重组和突变等方法,经过多次选择来培育新品种,耗时长,工作量大;分子标记辅助育种 (MAS) 会因为多代回交及连锁累赘的问题而导致标记丢失,影响作物育种进程[35]。水稻粒型作为数量性状受到生育时期及栽培环境的影响,增加了其选育难度[23]。由于三系杂交稻恢复系和不育系之间的严格关系,传统杂交选育方式增加了外来基因渗入的风险[10]。CRISPR/Cas9基因编辑技术可以快速精确地获得期望性状,打破了传统育种方法的局限性,开创了作物改良新时代[35-36]。基因编辑技术在水稻粒型改良上的成功应用,创制了不同需求的水稻粒型种质资源[20, 26, 32, 37-39]。本试验利用CRISPR/Cas9基因编辑技术,对短圆粒三系杂交稻恢复系GH582的多个粒型基因同时编辑,T2代即可选择符合目标性状,无转基因成分且突变位点纯合的植株,获得新恢复系GH582E(gw8tgw3gs3)。GH582E相比较GH582,除籽粒变得细长外,千粒重、单株重量等重要产量性状及其他主要农艺性状无显著差别。籽粒长度增加可提高千粒重,而千粒重的减少主要来自宽度的降低[40]。本研究选用GW8基因[15]为目的基因进行敲除以降低籽粒宽度,利用gs3和tgw3突变使籽粒长度增加[21-22],获得理想粒型突变的同时保证了新恢复系GH582E(gw8tgw3gs3)产量。
垩白是影响水稻外观品质的重要性状,其形成受多基因控制[42-44]。在复杂的调控路径中,籽粒外形对垩白形成有不可忽略的影响。粒宽基因GW2编码具有E3泛素连接酶活性的环型蛋白,基因功能的丧失提高了籽粒宽度和重量,同时垩白也显著增加[13]。水稻直链淀粉含量基因FLO2在过量表达的情况下,导致稻谷籽粒增大,并呈现垩白状和面粉状,品质下降[14]。GW8基因的缺失变异使籽粒变细,垩白出现频率大大降低,稻米外观品质得到改善[15]。粒形基因GW7通过促进纵向细胞分裂、减少横向细胞分裂产生细长籽粒,同时增加了胚乳发育时几种淀粉合成基因的转化水平,降低垩白,提高籽粒外观品质[16]。qLGY3基因的选择性剪接蛋白OsMADS1lgy3会产生细长粒的水稻品种,籽粒透明度增加而垩白减少[17]。Zhao等[18]利用CRISPR/Cas9技术在盐稻8号中获得GS9基因的缺失突变体gs9使水稻品种获得细长籽粒,垩白降低;Chen等[20]在日本晴中敲除基因GS3,获得的gs3突变体籽粒变长,垩白度及垩白粒率降低,外观品质提升;Huang等[32]在保持系Mei1B中敲除GS3基因,获得的新保持系Mei2B(gs3)籽粒增长,产量增加,品质也得到相应提升。本研究中GH582E相较GH582,由于籽粒显著变长变细,籽粒长宽比增加,米质分析显示其垩白度及垩白粒率显著下降,外观品质得到显著提高,而影响产量的千粒重、单株产量等性状无显著差异,说明3个粒型基因的同时突变影响了粒型和品质,对产量变化无显著影响。水稻粒型与产量相关,以往研究认为在水稻品质育种过程中,特别强调细长粒型会影响粒重和单株重,导致所育成的优质米品种产量偏低[40]。从本研究及相关粒型基因编辑文献的报道可知,选择适当的粒型基因对水稻品种进行改良,可获得理想的类型突变,满足生产需要。
4 结 论
利用CRISPR/Cas9技术改良水稻恢复系的粒型快速有效,并且仅通过粒型的改变就可显著提高水稻外观品质,获得产量和品质兼顾的水稻新品种,促进现有资源有效利用,提高杂交稻亲本的育种效率。
猜你喜欢
北京航空航天大学学报(2022年8期)2022-08-31
阅读(快乐英语高年级)(2022年6期)2022-06-17
家庭影院技术(2021年10期)2021-11-20
少先队活动(2020年6期)2020-12-18
江苏农业学报(2020年2期)2020-05-27
山东农业科学(2018年11期)2018-01-17
紫禁城(2017年6期)2017-08-07
幼儿智力世界(2017年1期)2017-04-26
江苏农业科学(2016年6期)2016-07-25
小学科学(2015年9期)2015-09-28
