
基于代谢组学和转录组学揭示郫县豆瓣中两株优势产风味菌株Bacillus amyloliquefaciens与Candida versatilis互作机制
2023-11-22 12:15:22刘欣宁卢云浩何强
中国调味品 2023年11期
刘欣宁,卢云浩,何强*
(1.四川大学 轻工科学与工程学院,成都 610065;2.成都大学 食品与生物工程学院,成都 610106)
郫县豆瓣,亦称为“四川豆瓣酱”,具有鲜辣醇厚、红棕油亮、黏稠绒实、酱香浓郁的特点[1-2],是川菜中必不可少的发酵调味品,被誉为“川菜之魂”[3]。近年来,随着川菜的风靡,郫县豆瓣产业逐步扩大,目前其年产值已近200亿元,在我国调味酱产业中占比高达20%,为促进地方经济增长发挥了重要作用[4]。郫县豆瓣的制备主要包括辣椒发酵、制曲-蚕豆发酵和混合后熟发酵3个阶段[5],并通过独特的“翻、晒、露”传统发酵工艺,最终形成独特的风味特征[6]。在此过程中,酱醅中的微生物群落在多相界面发生复杂的相互作用[7],从而实现信息传递、能量代谢、物质降解和分子转化等,为郫县豆瓣的风味形成作出了巨大贡献。
微生物间存在多种相互作用关系,包括互利共生、偏利共生、偏害共生、竞争等[8]。例如,在传统酸面团发酵中,Lactobacillussanfranciscensis和Kazachstaniaexigua可以相互协作利用麦芽糖,稳定微生物菌群,进而影响面团的品质[9]。在黄酒酿造过程中,Saccharomycescerevisiae的胞外代谢产物乙醇和有机酸会抑制Aspergillusflavus的生长,而添加A.flavus可促进S.cerevisiae的增殖,显著提高其产酯能力[10-11]。此外,浓香型白酒窖泥中Clostridiumfermenticellae生成的甲酸可被Novisyntrophococcusfermenticellae通过Wood-Ljungdahl途径利用,促进后者分泌乙酸,乙酸可进一步被C.fermenticellae通过逆β-氧化途径利用,从而积累丁酸和己酸[12]。前期研究表明[13-14],Bacillusamyloliquefaciens和Candidaversatilis是郫县豆瓣内源性优势细菌和酵母菌,能够高产呈酱香和果香的挥发性风味化合物,且在不同接种条件下,产品的风味具有显著差异,表明两株菌的互作可引起一系列代谢物的变化,但其互作机制仍需进一步揭示。
本文分析了体系中代谢物的变化及相关基因的差异表达,代谢组学和转录组学可揭示生物体调控机制,是生物学研究中的重要技术手段。因此,本文以B.amyloliquefaciens和C.versatilis为研究对象,基于非靶向代谢组学和转录组学技术,探究两株菌在共培养条件下代谢物和基因水平的变化,以揭示两株菌的互作机制。
1 材料与方法
1.1 材料与仪器
B.amyloliquefaciensMK063714、C.versatilisMK063708:分离自郫县豆瓣;甲醇、甲酸、醋酸铵(均为色谱纯)、Trizol RNA提取试剂盒、Ribo-Zero试剂盒:美国Thermo Fisher Scientific公司;其他试剂(均为分析纯):成都市科隆化学品有限公司。
LDZX-50FB立式压力蒸汽灭菌锅 上海申安医疗器械厂;1736R高速冷冻离心机 丹麦LaboGene公司;VanquishTMUHPLC色谱仪、Q ExactiveTMHF-X质谱仪 美国Thermo Fisher Scientific公司。
1.2 实验方法
1.2.1 发酵液的制备
添加50 g蚕豆粉、200 mL蒸馏水于500 mL锥形瓶中,混合均匀后灭菌,作为培养基备用。以活化后的B.amyloliquefaciens和C.versatilis制备4种发酵液:对照组(不接菌),单独接种B.amyloliquefaciens(BA组),单独接种C.versatilis(CV组),同时接种B.amyloliquefaciens和C.versatilis(CO组);菌株接种量均为2×107CFU/g。将培养基于30 ℃、150 r/min培养1 d后,加入30 g氯化钠,快速混匀后置于相同条件下培养7 d。培养完成后,在4 ℃下以5 000 r/min离心10 min,分别收集上清液及沉积物,然后用液氮速冻后于-80 ℃保存并尽快用于后续分析。上清液用于代谢产物测定,沉积物用于RNA提取。
1.2.2 非靶向代谢组学分析
100 μL液体样本利用400 μL甲醇沉淀蛋白后,加入500 μL含0.1%甲酸的80%甲醇水溶液,涡旋振荡后冰浴静置5 min,然后以15 000 r/min、4 ℃离心10 min,取一定量的上清液,加质谱级水稀释至甲醇含量为53%,并置于离心管中15 000 g、4 ℃离心10 min,收集上清液,利用UPLC-MS/MS进行分析[15]。以含0.1%甲酸的53%甲醇水溶液作为空白样品。
1.2.3 转录组学分析
RNA的提取参照Trizol说明书,并采用RNeasy Mini Kit试剂盒进行纯化。去除rRNA后,利用TruSeq®Stranded mRNA Library Prep Kit进行文库构建,并基于Illumina HiSeqTM2500平台进行双末端测序。文库构建及RNA-seq测序由北京诺禾致源生物科技有限公司完成。
1.2.4 数据分析
代谢组学中,所有结果均呈现为剔除空白和对照组样品后的数据,将同时满足以下条件的物质确定为差异表达代谢物:变化倍数(fold change,FC)≥2为上调,FC≤0.5为下调;P<0.05;VIP(variable important in project)≥1.0[16]。RNA-seq测序中,通过DEGSeq 1.18.0软件对基因表达进行差异分析,差异基因筛选标准为|log2(fold change)|>1,且校正后的P<0.001。将差异表达基因映射到Gene Ontology(GO)数据库和Kyoto Encyclopedia of Genes and Genomes(KEGG)数据库以分析其功能和途径[17]。
2 结果与讨论
2.1 非靶向代谢组学分析
样品BA组和CO组共有193种代谢物具有显著差异,而样品CV组和CO组的显著差异代谢物则有297种,这些物质均集中在脂类和类脂分子、有机酸及其衍生物、氨基酸及其衍生物、苯环型化合物等(见图1)。可以发现,共培养对酵母菌的影响强于对细菌的影响,这可能与B.amyloliquefaciens产蛋白酶等酶系的能力优于C.versatilis有关[18-19],表明该芽孢杆菌可为C.versatilis代谢形成风味物质提供前体物。

图1 差异代谢物富集气泡图Fig.1 Bubble chart of differential metabolite enrichment
主要差异代谢物见表1。

表1 主要差异代谢物比较Table 1 Comparison of main differential metabolites
样品CO组较样品BA组显著上调的代谢物有二十二碳四烯酸、油酸、棕榈酸甲酯和麝香酮,上调倍数均高于100倍;此外,香芹酮、可的松、十五烷酸、对羟苯基乙醇等物质含量也增加了20倍以上。这些物质主要为脂类化合物,表明C.versatilis可显著加速脂类的降解,这与C.versatilis分泌酯酶的能力远强于B.amyloliquefaciens的研究结果一致[20-21],并且在郫县豆瓣实物接种发酵模型中得到验证:相较于单独接种芽孢杆菌,同时接种上述两株微生物的样品中含有更高浓度的长链脂肪酸、脂肪酸甲酯或脂肪酸乙酯类化合物[22-23]。
而相较于样品CV组,CO组中显著上调的化合物具有相似的结果,主要集中在脂类和类脂分子,如油酸、棕榈酸甲酯、二十二碳四烯酸、可的松等化合物均上调30倍以上,这表明B.amyloliquefaciens和C.versatilis在这些化合物的形成中具有加和或协同效应。另外,许多多肽及氨基酸含量也上调了20倍以上,如Pro-Leu、Gly-Tyr-Ala、Gly-Tyr、羟脯氨酸、肌肽、L-酵母氨酸、O-乙酰丝氨酸、组氨酸等,但相较于样品BA组,所有鉴定到的二肽化合物均在CO组中显著下调。结果表明B.amyloliquefaciens较C.versatilis更利于形成与蛋白质相关的代谢物(B.amyloliquefaciens产蛋白酶和氨肽酶能力更强[24]),且后者可能会降低芽孢杆菌产相关酶系的能力。此外,C.versatilis较B.amyloliquefaciens可能更易形成氨基酸衍生物,该结论可以从样品CO组下调的氨基酸类物质中获得。相较于样品CV组,CO组中下调的氨基酸类物质主要为其衍生物,例如苯乙酰甘氨酸、甲硫氨酸硫氧化物、L-焦谷氨酸。
2.2 转录组学分析
2.2.1 基因表达差异对比分析
由于C.versatilis没有模式参考菌株,无法有效地进行基因表达量的比较,本节重点基于B.amyloliquefaciens的基因表达变化来剖析两株菌的互作机制,差异表达基因火山图和热力图见图2,B.amyloliquefaciens在样品BA组和CO组之间共有1 606个显著差异表达基因,其中上调表达基因有801个,下调表达基因有805个。

图2 差异表达基因火山图(A)和热力图(B)Fig.2 Volcano plot (A) and heatmap (B) of differentially expressed genes
2.2.2 差异基因GO富集分析
GO富集分析结果见图3,共发酵体系(CO组)中,B.amyloliquefaciens显著性表达的基因集中在生物过程和分子功能类别,上调表达以生物过程显著性更高,如核苷单磷酸合成及代谢、ATP代谢、嘌呤核苷单磷酸代谢等10个过程,且各类别差异基因的表达数量也很高(均>20个);而下调表达以分子功能类别更显著,且除作用于tRNA的催化活性外,其他类别基因下调数量均高于60个。
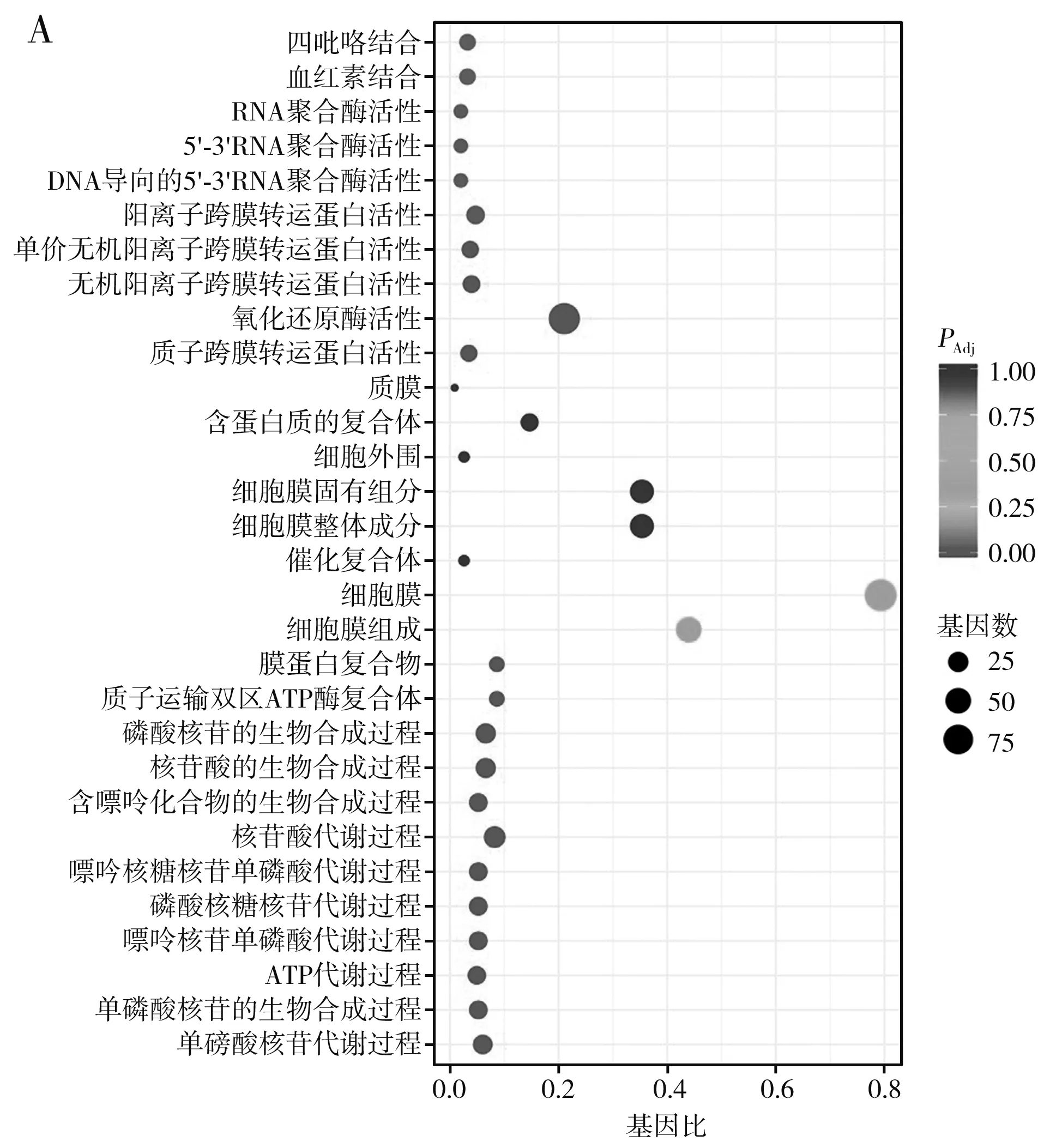
图3 B. amyloliquefaciens差异表达基因的GO富集分析散点图Fig.3 Scattering plot of differentially expressed genes in B. amyloliquefaciens by GO enrichment analysis
在下调表达基因中,与有机环状化合物代谢、杂环代谢和细胞芳香族化合物代谢过程相关的基因表达数量均超过100个,含硫化合物与吡咯化合物代谢也有相关基因下调表达,这说明相较于样品BA组,样品CO组与上述代谢相关的化合物的生成量会更低,该结果与前文基于代谢组学的研究结果一致,样品CO组中有机杂环化合物和苯环型化合物等化合物的下调数量及比例均更高,该结果也与Lu等[22]的研究结果相匹配,B.amyloliquefaciens与C.versatilis混合发酵时,郫县豆瓣样品中多数吡嗪、呋喃(酮)等杂环化合物以及含硫化合物含量均比单独接种B.amyloliquefaciens发酵的样品更低。不过,两种不同接种方式的样品在上述化合物中的含量差异较小,这与下调表达基因的显著性并不高(1.2<-log10(PAdj)<1.6)有关。
2.2.3 差异基因KEGG富集分析
对样品BA组和CO组中差异表达基因进行KEGG富集分析,其中20条富集通路见图4。
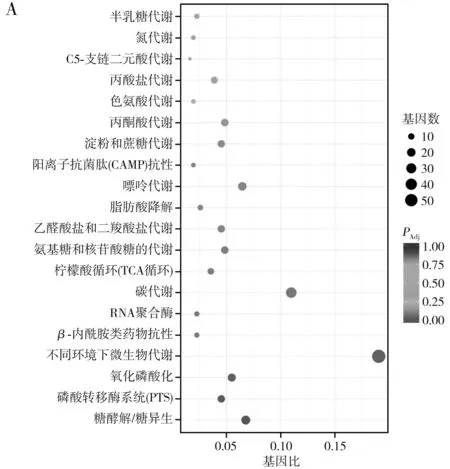
图4 B. amyloliquefaciens差异表达基因的KEGG富集散点图Fig.4 Scattering plot of differentially expressed genes in B. amyloliquefaciens by KEGG enrichment
B.amyloliquefaciens在样品CO组上调表达的基因中,注释到KEGG最显著的通路为糖酵解/糖异生途径(-log10(PAdj)≈2.5),其次为磷酸转移酶系统、氧化磷酸化和微生物代谢(-log10(PAdj)≈1.5),而包括碳代谢、脂肪酸降解、丙酮酸代谢在内的其他途径显著性相对较低(-log10(PAdj)<1.0)。而基因表达量方面,以微生物代谢活动相关的基因上调数量最高(59个),其次为碳代谢(34个)、糖酵解/糖异生(21个)。
下调表达基因中,最显著的通路包括叶酸合成和硫中继系统途径(-log10(PAdj)>2.0),其次为RNA降解、多种氨基酸(苯丙氨酸、酪氨酸、色氨酸、缬氨酸、亮氨酸和异亮氨酸)的生物合成及硫氨酸代谢等(-log10(PAdj)>1.0)。而基因表达量方面,以ABC转运蛋白相关的数量最多,为27个,其次为群体感应(17个)、叶酸合成(15个)和鞭毛装配(14个)等。
混合培养中B.amyloliquefaciensKEGG代谢途径分析见表2。
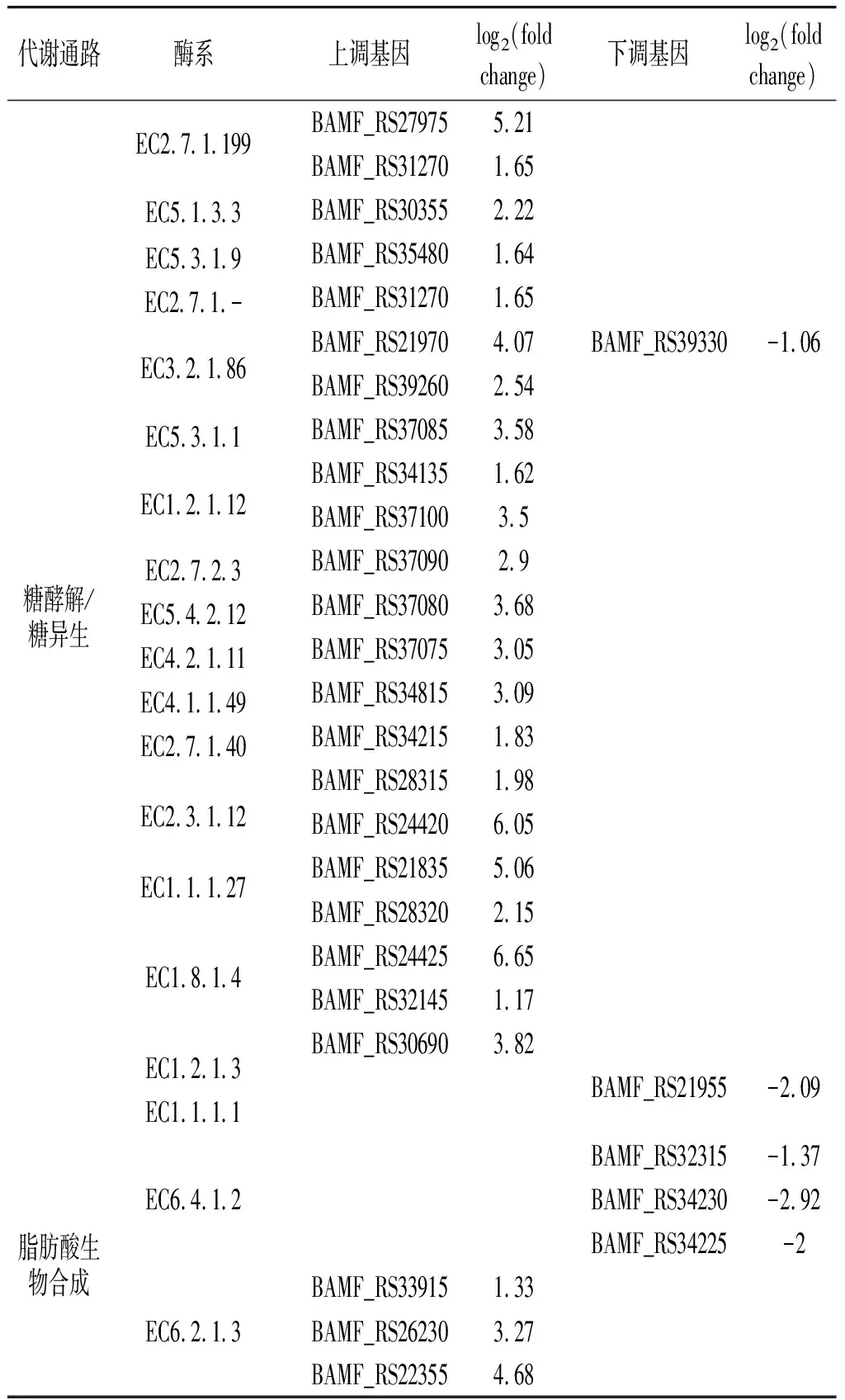
表2 混合培养中B. amyloliquefaciens KEGG代谢途径分析Table 2 Analysis of KEGG metabolic pathways of B. amyloliquefaciens in co-culture system
KEGG糖酵解/糖异生代谢途径中,除醇脱氢酶(EC1.1.1.1)外,其他几乎所有监测到的基因均在共发酵体系(CO组)中上调表达,这有助于乙醇的积累,进而合成更多的酯类化合物。另外,共培养体系下,除长链脂肪酸辅酶A连接酶(EC6.2.1.3)外,与脂肪酸合成代谢途径相关的基因均在B.amyloliquefaciens中下调表达;而除醇脱氢酶和酰基转移酶(EC1.1.1.1,EC2.3.1.9)外,其他与脂肪酸降解代谢途径相关的基因均上调表达。这些结果均与Lu等[22]的研究结果相似,B.amyloliquefaciens与C.versatilis共发酵的郫县豆瓣样品中醇类、酯类化合物含量显著高于单独接种B.amyloliquefaciens的样品,而脂肪酸类化合物的含量整体上更低。
从丁酸盐代谢通路来看,与乙偶姻前体物丙酸盐形成的相关基因(EC1.1.1.83)在共培养体系中下调最显著,而与其直接相关的基因(EC1.1.1.4,EC4.1.1.5,EC1.1.1.303)上调表达相对较弱,这可能是单独接种B.amyloliquefaciens的郫县豆瓣样品中乙偶姻的含量高于混合接种发酵样品的原因。另外,混合发酵体系下含硫化合物(3-甲硫基丙醛、3-甲硫基丙醇)的含量均低于单独接种B.amyloliquefaciens发酵的样品[22,25],这也与B.amyloliquefaciens中与蛋氨酸和半胱氨酸代谢相关的基因(EC2.5.1.47,EC4.4.1.2)在共培养体系中显著下调表达一致。
代谢组学和转录组学分析结果表明,B.amyloliquefaciens与C.versatilis共培养时,可引起B.amyloliquefaciens中大量基因表达的变化,其中,基因上调表达的显著性高于下调表达基因。显著上调的代谢途径主要包括糖酵解/糖异生、脂肪酸降解等,而与脂肪酸合成、氨基酸相关的代谢通路显著减弱,表明C.versatilis对B.amyloliquefaciens的生长和代谢具有双向作用,进而引起一系列代谢产物的差异。
3 结论
本文基于非靶向代谢组学和转录组学技术,探究了B.amyloliquefaciens与C.versatilis的互作机制。共培养时,C.versatilis对B.amyloliquefaciens的生长和代谢具有双向作用,且B.amyloliquefaciens基因上调表达的显著性高于下调表达基因。其中,与糖酵解/糖异生、脂肪酸降解等途径相关基因表达显著增强,而与脂肪酸合成、氨基酸相关的基因表达显著减弱,进而引起代谢产物的变化,两株菌共培养可使B.amyloliquefaciens上调表达脂类和类脂分子,下调表达有机酸及其衍生物类;可使C.versatilis上调表达脂肪酸或类脂化合物、多肽或氨基酸类代谢产物,下调表达氨基酸衍生物、苯环型化合物、苯丙素和聚酮类化合物。
猜你喜欢
食品与发酵工业(2023年19期)2023-10-18 00:28:36
现代临床医学(2022年4期)2022-09-29 07:36:10
宁夏医学杂志(2020年3期)2021-01-21 08:23:24
天然产物研究与开发(2018年9期)2018-10-08 03:25:32
天然产物研究与开发(2018年4期)2018-05-07 06:48:03
中国品牌(2016年1期)2016-03-19 06:49:08
分析测试学报(2015年7期)2016-01-13 06:19:16
摄影世界(2015年9期)2015-09-15 16:10:26
质谱学报(2015年5期)2015-03-01 03:18:37
中国医疗美容(2015年5期)2015-02-03 03:02:09
