
基于SSR荧光标记的广西天冬种质资源遗传多样性分析
2023-11-22 13:36郑德波庾韦花
种子 2023年9期
郑德波, 石 前, 蒙 平, 庾韦花
(广西壮族自治区农业科学院生物技术研究所, 南宁 530007)
天冬(Asparaguscochinchinensis)是百合科天门冬属的藤本植物,是我国常用的传统大宗中药材之一,以干燥块根入药,始载于《神农本草经》,并被历代本草收录。天冬性寒、味甘、微苦,具有生津止渴、养阴润燥、清肺等功效,主治咽干口渴、内热消渴、肺热燥咳等症状[1]。Lei等[2]研究表明,天冬的一些水溶性提取物具有抗氧化延缓衰老的功效。长期以来,天冬以野生块根入药为主。随着市场需求的逐渐加大,过度采挖导致天冬野生资源日渐枯竭[3],天冬已成为国家重点保护的野生药材物种之一[4]。野生资源人工栽培是实现天冬资源可持续发展的重要途径之一,虽然天冬人工栽培技术研究在全国各地有所开展,但其种源以野生种质为主且来源混乱,导致天冬的产量和品质都无法保障。广西是我国天冬的重要产地之一,天冬种质资源十分丰富,遍布全区[5]。天冬种质资源遗传多样性、亲缘关系和系谱关系的研究是其资源的搜集保存和利用的理论基础,开展这方面的研究是非常必要和迫切的。
简单重复序列(Simple Sequence Repeat,SSR)标记是广泛分布于各个物种中的一类标记,具有共显性、多态性丰富、重复性好和操作简单等优点,在动植物分子标记遗传连锁图谱的构建、种质资源评估、基因定位、分子标记辅助选择、品种真实性和种子纯度鉴定等领域的研究都有很好的应用价值[6]。
荧光标记毛细管电泳技术是在PCR扩增时使用有荧光标记的引物替代普通引物,扩增出来的片段一条链带上荧光标记,可直接读取目标片段准确大小,大大降低了分辨扩增产物的难度,提高了研究的效率和准确性。目前,该技术已应用于粮食作物(玉米[7]、水稻[8]等)、果树(桃[9]、杧果[10]、火龙果[11]等)和中药材(当归[12]、三七[13]、三叶青[14]等)一些物种的分子标记研究中。相较于传统聚丙烯酰胺凝胶电泳,毛细管自动电泳荧光检测法[15]是较为先进的检测技术。该技术通过不同颜色标记荧光引物,检测结果可精确到1 bp,能够实现低成本、高效率地检测多个SSR标记、多个位点信息。
天冬的相关报道以其药用成分的研究为主,种质资源遗传多样性的研究报道较少。目前,国内仅有两篇关于天门冬种质资源ISSR遗传多样性的研究报道[16-17]。ISSR标记为显性标记,无法区分显性纯合基因型和杂合基因型,其标记数量有限,PCR扩增时对DNA模板的质量要求较高,而且其PCR产物检测自动化程度不高,这些不足都极大地限制了天冬种质资源遗传多样性研究的开展。SSR荧光标记则很好解决了这些问题。广西作为国内天冬的重要产地之一,天冬种质资源极其丰富,而前人的研究中鲜有广西天冬材料的报道。因此,开展广西周边地区的天冬种质资源的遗传多样性研究,有助于理清现有各种质间亲缘关系的远近,发掘广西的优异天冬资源,为广西天冬育种和生产提供优异种质,进而为广西的天冬产业的可持续发展提供理论依据和技术支持。目前,SSR荧光标记毛细管电泳技术用于天冬种质资源遗传分析的研究笔者尚未见报道。因此,本研究借助该技术对25份天冬种质资源进行遗传多样性分析,优化天冬SSR毛细管电泳荧光检测技术,为天冬种质资源鉴定及杂交育种亲本选择的遗传分析提供理论基础和技术支撑。
1 材料与方法
1.1 试验材料
本试验中所选用的25份天冬材料均为本单位药用植物利用团队经过多年收集整理,经中国医学科学院药用植物研究所广西药用植物园植物分类学专家鉴定,种植保存于广西农科院生物所大棚及里建基地。经鉴定,这些天冬材料均为百合科天门冬属天门冬种[Asparaguscochinchinensis( Lour. ) Merr.],采集地不同,植株性状稍有不同。主要包括广西及其周边省份、邻国越南等地的野生种及栽培种,详情如表1所示(共取样27份,其中样品18由于样品量太少而弃用,样品17由于在与其他25份样品多样性分析时数值异常而剔除)。取这些材料的叶片用于提取总DNA。主要仪器设备及试剂:PCR 仪(杭州晶格K960);水平电泳仪(北京君意 JY300C);凝胶成像仪(北京科创K8160 );测序仪(美国应用生物ABI 3730XL);Taq酶、dNTP、DL2000(宝生物TaKaRa)。
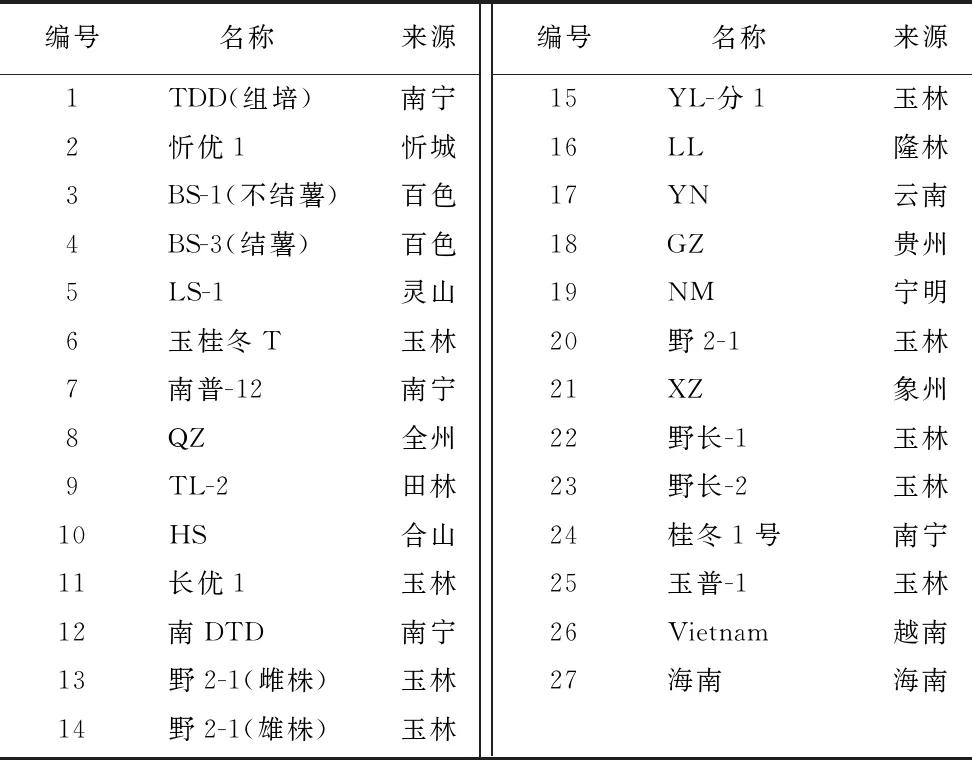
表1 25份天冬材料编号及来源
1.2 天冬基因组DNA提取
天冬叶片中多糖多酚等次生代谢物较多,DNA提取较困难,故参照韦荣昌等[18]的改良CTAB法进行DNA提取。
1.3 SSR引物筛选
普通引物参照Manish Kapoor等[19]报道的SSR 引物序列,由通用生物系统(安徽)有限公司合成。用17,24,25,26 这4个样本对24对引物进行筛选。多态性引物筛选PCR反应体系参考曾桂萍等[20]采用的20 μL体系稍作优化:dNTP 0.4 μL,Buffer 2 μL, SSR引物0.6 μL[其中,F链 0.3 μL(20 μmol/L),R链0.3 μL(20 μmol/L)],DNA模板2 μL,Taq0.2 μL,ddH2O 14.8 μL。扩增程序:94 ℃预变性5 min;94 ℃变性30 s,46~52 ℃复性35 s,72 ℃延伸40 s,共 35个循环;最终72 ℃延伸3 min。
1.4 SSR-PCR扩增及毛细管电泳检测
所有参试样品荧光引物PCR扩增时所采用的反应体系和扩增程序与多态性引物筛选完全相同,只是将之前的普通引物的F链换成加注FAM(蓝)荧光染料的F链。扩增产物在 3730 测序仪(ABI3730XL)进行毛细管电泳,筛选出多态性较好的位点。
1.5 数据分析
根据Convert1.31软件要求,将每个样品在各个等位基因位点的片段大小按格式录入到Excel软件中,然后用Convert1.31软件转化成POPGENE软件所要求格式。使用POPGENE1.32软件分别进行各引物及各种群的遗传多样性分析。用Nei(1972)的遗传一致度I(Genetic Identity)和遗传距离GD(Genetic Distance)来衡量各种群之间的遗传分化大小。基于遗传相似系数,使用NTSYS Version2.10e软件中UPGMA法对各样品进行聚类分析,构建系统发育树。
2 结果与分析
2.1 SSR引物的多态性分析
24 条 SSR 引物中,共筛选出 8条多态性好且条带清晰的 SSR 引物(表2)。

表2 多态性SSR引物信息
通过8对引物检测,25个天冬样本共检测出48个等位基因位点。等位基因数目中,最小为引物P8,多态性比率为100%;最大等位基因数目为12(引物P15),多态性比率为83%,平均每个位点等位基因数目为4.1。遗传多样性参数结果表明:观测等位基因数(Na)平均为6.0个,有效等位基因数平均为3.81;期望杂合度平均为0.66,PIC平均为0.60。以上结果表明,25份天冬种质资源间具有较为丰富的遗传多样性,部分引物对供试样品的检测结果(图1)。Shannon指数和基因多样性指数平均值分别为1.33和0.64,表明供试样品具有较高的遗传多样性(表3)。
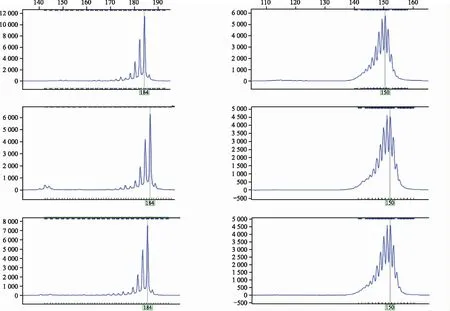
注:左为P15,右为P23;上为样品7,中为样品19,下为样品25。

表3 SSR多态引物相关指标
2.2 25份天冬种质资源的群体遗传结构分析
采用Evanno 等[21]的方法计算得到的最适 deltaK值为4(表4),表明 25 份样品来自 4 个基因库。由表4可知,当K=4时,样品2、样品10、样品11、样品15、样品16、样品21和样品27的基因主要来自基因库1,样品4、样品5、样品8和样品12的基因主要来自基因库2,样品,1、样品3、样品6、样品13、样品14、样品20、样品24和样品26的基因主要来自基因库3,样品7、样品19、样品23和样品25的基因主要来自基因库4,样品9的基因主要来自基因库3,基因库4,样品22的基因主要来自基因库1,基因库2,基因库3。说明这些天冬材料非均质、具有明显的遗传结构。
2.3 25份天冬种质资源的遗传相似性分析
利用 8对SSR引物标记,用 NTSYS 软件计算得到 25 份天冬样本的遗传相似性系数。结果(表5)表明,25份材料的遗传相似性系数为0.08~1.00,平均为0.44。其中遗传相似性系数最小的为样品16(LL,隆林)和样品20(野2-1),说明二者亲缘关系较远,遗传差异相对大;遗传相似性系数最大为样品7(南普-12)、样品19(NM,宁明)和样品25(玉普-1),说明三者遗传差异相对较小,亲缘关系比较相近。遗传相似性结果分析表明,测试的 25份天冬种质遗传多样性丰富。值得关注的是,样品16与样品4,5,8,9,12,14,20,26这8个样品的遗传相似系数均较小,遗传差异明显,在资源创制和新品种选育时,可以重点关注这个材料。

表5 25份天冬种质资源的遗传距离与遗传相似系数
2.4 25份天冬种质资源的聚类结果分析
利用 NTSYS V2.1 软件对 25份供试材料进行聚类分析,得到相应的聚类图(图2),在遗传相似系数0.32处,25份供试天冬材料可分为类群Ⅰ和类群Ⅱ两大类。其中, 21 份天冬样品在类群Ⅰ中, 剩余4 份天冬样品(长优1、XZ、海南和LL(隆林))归入类群Ⅱ中。在遗传相似系数0.52处,类群Ⅰ又可分为4个亚群,第一个亚群(ⅰ)包括:TDD组培苗、Vietnam(越南)、BS-1(不结薯)、桂冬1号、野2-1(雄株)、YL-分1、玉桂冬T、野2-1(雌株)和野2-1 9个样品;第二个亚群(ⅱ)包括:BS-3(结薯)、LS-1、QZ、南DTD和野长-1;第三个亚群(ⅲ)包含:南普-12、NM、玉普-1、野长-2和TL-2;第四个亚群(ⅳ)包含:忻优1和HS。

图2 25份天冬种质资源的聚类结果
3 讨 论
SSR引物具有数量多、多态性好、共显性和易于PCR操作等特点,因而被广泛应用于植物种质资源遗传多样性研究中。对于已测序完成的物种,其基因组序列可以在NCBI等数据库上查询到,广大科研工作者可以根据基因组序列开发大量的SSR引物,广泛应用到各研究领域。对于一些尚未完成全基因组测序的物种,由于没有已知的基因组序列,无法开发SSR引物。随着测序技术的进步,可以通过简化基因组测序等手段获得部分基因组片段序列,用于SSR引物的设计。另外,还可以根据近似物种已开发的SSR引物来筛选目标物种的多态性SSR引物。本研究中,由于天冬分子生物学研究起步较晚,没有参考基因组的数据,无法直接开发SSR引物,笔者通过查阅相关文献,根据Manish Kapoor等[19]在天门冬属植物研究中采用的24对SSR引物,筛选出8对多态性SSR引物用于天冬种质资源遗传多样性研究,取得了较好的效果。
所筛选的8对SSR引物中,其中5对的PIC值高于0.6,表明它们在本试验材料中有很好的多样性,与实际的结果相符。这个结果很好地验证了Caruso等[22]、Li等[23]、斯钦毕力格和包文泉[24]、白冬梅等[25]通过已知近缘物种开发SSR引物的可行性。
荧光标记毛细管电泳技术以其准确度高、易于自动化检测的特点,已被广泛应用于许多物种的遗传多样性研究中。如何准确地将不同批次、不同胶块的数据整合在一起进行比较分析,一直是SSR标记聚丙烯酰胺凝胶电泳检测方法的一个难题。在SSR标记聚丙烯酰胺凝胶电泳检测过程中,虽然每块胶上都设置了标准带型对照,待检样品的等位基因类型可以通过与标准带型对照比较确定的办法加以解决,但聚丙烯酰胺凝胶电泳的分辨率有限,当待检样品在凝胶泳道上的位置与标准带型对照较远,且标准带型对照相邻等位基因片段差异较小时,待检样品的等位基因类型难以确定,对数据的准确性有很大的影响。而SSR荧光标记毛细管电泳检测方法,则用内标代替标准带型对照,依据待检样品目标峰的位置,与同一毛细管泳道的内标比较,直接读出目标DNA片段的准确大小。与常规的凝胶电泳检测方法相比,荧光标记毛细管电泳检测数据显然更为精确,同时很好地解决了大规模不同批次数据有效整合和准确比较的问题。基于荧光标记毛细管电泳技术自身的特性,多年多点的数据可有效整合在一起,并可以准确比较。因此,荧光SSR毛细管电泳为今后天冬的跨年份、跨地域的协作研究提供了技术保障。
25份天冬种质资源的聚类结果与真实的分布有较大的出入,其地理来源出现明显的分化。自育品种“桂冬1号”与BS-1(不结薯)间的遗传关系最近,其遗传相似系数达0.80,遗传距离仅为0.22。种植在里建基地的野2-1(雌株)和野2-1(雄株)是从生物所大棚野2-1这个种质资源的种子实生苗人工选择的后代,可能是人工选择的缘故,其与原种质间的遗传相似系数分别为0.84,0.80,相应的遗传距离分别为0.18,0.22。而野2-1(雌株)和野2-1(雄株)也由于人工选择的原因发生较大的遗传分化,虽然聚类到一个大群但变异稍大,两者的遗传相似系数为0.696,遗传距离则为0.362。南普-12、NM和玉普-1三个样品的遗传相似系数为1,遗传距离为0,这就表明三者在所筛选的8对引物中的多样性表现完全相同。要想把这3个种质资源完全区分开,需要开发更多的SSR引物。在排除人为操作失误(取样混杂)的情况下,最可能的原因就是天冬在人工栽培的过程中由于三地间长期的相互引种造成了种质资源的混乱。值得进一步关注的是,来自云南的17号种质表型上与其他种质有较大的差异,且在本试验中8个多态性标记的电泳结果中也表现出极高的多态性,但在聚类分析中却不能与其他25个种质中的任何一个聚类在一起,具体原因有待进一步研究。
猜你喜欢
阅读(高年级)(2022年9期)2022-10-08
今日农业(2022年13期)2022-09-15
今日农业(2021年1期)2021-11-26
云南化工(2020年11期)2021-01-14
国际种业前沿动态(2020年18期)2020-12-23
中国麻业科学(2018年6期)2018-04-09
科学24小时(2016年11期)2016-11-08
西南农业学报(2016年5期)2016-05-17
广西林业科学(2016年3期)2016-03-16
杂草学报(2015年2期)2016-01-04
