
ESET通过lncRNA GMDS-AS1调控肝癌细胞增殖、凋亡、糖酵解的研究
2023-11-21 06:15黄俊玲钟腾猛黄森平高鑫艳
安徽医药 2023年12期
黄俊玲,钟腾猛,黄森平,高鑫艳
作者单位:1右江民族医学院附属医院消化内科,广西壮族自治区 百色533000;2百色市人民医院肝胆外科,广西壮族自治区 百色533099;3右江民族医学院临床医学院,广西壮族自治区 百色533000
肝细胞癌(HCC)是世界范围内最常见的人类癌症之一,其死亡率居所有癌症之首[1]。HCC的发病是由于表观遗传的改变导致癌基因的激活和抑癌基因的失活,进而诱导的肝细胞癌[2]。表观遗传的改变包括异常甲基化、组蛋白修饰和核糖核酸(RNA)干扰,这不会改变遗传密码,但会影响mRNA的转录[3]。组蛋白甲基转移酶集合域分叉1(SET domain, bifurcated 1,ESET)基因位于染色体1q21上,编码143 kDa的蛋白质,具有多个功能域。大量研究证明,ESET在多种恶性肿瘤中高度表达,与肿瘤细胞的增殖、侵袭紧密相关[4]。ESET的过度表达与HCC进展、肿瘤侵袭性和HCC病人的低生存率显著相关,ESET的失活降低HCC细胞的增殖和迁移能力,表明ESET是HCC中的重要癌基因。遗憾的是,ESET发挥作用的潜在调控机制研究甚少。长链非编码RNA(lncRNA)起初被认为作为是多余的,现在很多研究已证实,lncRNA在生物功能调控中具有重要作用,尤其肝细胞癌[5]。多种多样的lncRNA在肿瘤的发生和转移中起关键作用[6],但仍有部分新发现的lncRNA的作用仍不完全清楚。lncRNA甘露糖4,6-脱水酶反义RNA1(GDP-Mannose 4,6-Dehydratase Antisense RNA1,lncRNA GMDS-AS1)是近期在肺腺癌中新发现的lncRNA[7],其在肝癌中的作用尚未可知。本研究拟以肝癌细胞为研究对象,观察ESET、lncRNA GMDS-AS1在肝癌组织中的表达及对肝癌细胞增殖、凋亡、糖酵解的作用,揭示ESET、lncRNA GMDS-AS1在肝癌细胞中发挥调控作用的潜在关系,为肝癌的治疗提供新的治疗靶点。
1 资料与方法
1.1 一般资料 样本来自右江民族医学院附属医院和百色市人民医院2019年1月至2020年1月期间接诊的行肝癌手术切除的49例病人的癌组织及对应的癌旁组织。所有病人均签署知情同意书,本研究符合《世界医学协会赫尔辛基宣言》相关要求。
人永生化正常肝细胞THLE-2、肝癌细胞HepG2购自美国菌种保藏中心;DMEM培养液、胎牛血清购自美国Gibco;Lipofectamin 3000试剂购自美国Thermo Fisher Scientific;TRIzol液购自美国Invitrogen;反转录试剂盒、定量聚合酶链反应(qPCR)试剂盒购自日本Takera;鼠抗ESET(A-1)单抗、ESET siRNA质粒购自美国圣克鲁斯生物;兔抗细胞增殖抗原标志物Ki-67单抗、兔抗葡萄糖转运蛋白1(GLUT1)多抗、兔抗乳酸脱氢酶A(LDHA)单抗、兔抗活化胱天蛋白酶-3(C-caspase-3)单抗均购自Cell Signaling Technology;HRP标记的二抗购自北京索莱宝公司;细胞计数试剂盒(CCK8)购自上海东仁化学研究所;膜联蛋白-V异硫氰酸荧光素/碘化丙啶(Annexin V-FITC/PI)细胞凋亡检测试剂盒购自上海翌圣生物;RNA结合蛋白免疫沉淀(RIP)试剂盒购自Millipore公司。
1.2 方法
1.2.1 细胞培养 THLE-2、HepG2细胞使用混有10%胎牛血清的DMEM培养液培养。培养条件为37 ℃、5%二氧化碳的恒温细胞培养箱中培养,隔天更换一次培养液。
1.2.2 细胞转染与分组 正常培养的THLE-2、HepG2细胞设为THLE-2、HepG2组。将HepG2细胞分为空白组(不做任何处理)、空载体组(转染pcDNA或si-con)、敲减ESET组(转染si-ESET)、过表达lncRNA GMDS-AS1组(转染pcDNA-GMDS-AS1)。具体的转染方法:使用3倍的Lipofectamin 3000转染试剂与各组待转染物质(DNA或质粒)混合与HepG2细胞共转染8 h,更换新培养液继续培养。实时荧光定量逆转录聚合酶链反应(qRT-PCR)或蛋白质印迹法确认转染效率。
1.2.3 qRT-PCR实验检测ESET、lncRNA GMDSAS1的表达 收集细胞,Trizol液提取总RNA,反转录试剂盒合成互补DNA(cDNA),-20 ℃保存待用。以cDNA为模板qPCR实验的模板,按照qPCR试剂盒要求操作,检测分析ESET、lncRNA GMDS-AS1的表达。以甘油醛-3-磷酸脱氢酶(GAPDH)为内参,2-ΔΔCt法计算ESET、lncRNA GMDS-AS1的表达水平。仪器程序设置为:95 ℃,2 min;95 ℃,30 min;60 ℃,30 s;72 ℃,30 s;72 ℃,5 min,41个循环。ESET,正向引物5'-AAGACCAGAAGCTCCGTGAA-3',反向引物5'-CCTGGGAACTGCTCTTCTTG-3';lncRNA GMDS-AS1,正向引物5′-AATGCTTTGAGGCCAAGCTA-3′,反向引物5′-TGGGTTCATAAGGGTTGCAT-3′;GAPDH,正向引物5'-GACAACAGCCTCAAGATCATCAG-3',反向引物5'-GTGGCAGTGATGGCATGGA-3'。
1.2.4 蛋白质印迹法检测ESET、Ki-67、GLUT-1、LDHA、C-caspase-3的蛋白表达 收集细胞或组织匀浆,冰上放射免疫沉淀法裂解30 min,提取总蛋白,测定蛋白浓度,变性。取变性后的蛋白上清做蛋白电泳上样模板。十二烷基硫酸钠-聚丙烯酰胺凝胶电泳蛋白电泳结束后,用转膜仪将蛋白从凝胶转移至NC膜,转模环境需在0 ℃以下。使用含2.5%脱脂奶粉的封闭液对膜封闭处理(37 ℃孵育2 h)。洗膜3次,将膜浸入稀释的一抗溶液中(鼠抗ESET(A-1)单抗,1∶800;兔抗Ki-67单抗,1∶1 000;兔抗GLUT-1多抗,1∶1 500;兔抗LDHA单抗,1∶1 000;兔抗C-caspase-3单抗,1∶2 000),4 ℃孵育过夜。充分洗膜,将膜转入稀释的二抗溶液中,37 ℃孵育2 h。洗膜,用ECL发光液显影,曝光。Image J软件分析蛋白灰度值,以目的蛋白灰度/内参GAPDH的灰度表示蛋白的表达量。
1.2.5 CCK8法检测细胞活性 收集细胞,调至0.5×105个/毫升,取200 μL细胞接种至96孔板,更换新鲜培养液,分别培养24、48、72、96 h。取出培养24、48、72、96 h的各组细胞,向细胞中加入10 μL的CCK8反应液,避光孵育20 min。在490 nm波长下检测细胞吸光度[D(λ)490]。
1.2.6 Annexin V-FITC/PI检测细胞凋亡 收集细胞,洗涤后用结合缓冲液制成悬液,取500 μL至于反应管,再依次加入Annexin V-FITC(5 μL)、PI(5 μL),避光孵育20 min,上流式细胞仪分析细胞凋亡。总凋亡率(%)=早期凋亡率(%)+晚期凋亡率(%)。
1.2.7 RIP实验检测ESET与lncRNA GMDS-AS1的相互作用 收集细胞,调至1×107个/毫升,取1.0 mL至EP管,用磷酸缓冲盐溶液洗涤3次,加入裂解液冰上裂解。结束后,分别向EP管中加入5 μL异染色质相关小RNA、免疫球蛋白G(IgG)抗体、Argonaute 2蛋白(Ago2)抗体,再加入2.5 μL的RNA酶抑制剂,轻轻吹打混匀。4 ℃低温离心10 min(12 000 r/min),洗涤3次,取上清置于磁珠中,4 ℃孵育过夜。用洗涤液洗涤磁珠3次,将磁珠结合复合物用于qRT-PCR实验。
1.3 统计学方法 实验中的数据均用SPSS 22.0进行专业的统计分析。计量资料使用表示。两组数据比较使用独立样本t检验,多组数据之间的比较使用单因素方差分析联合LSD-t检验。P<0.05为差异有统计学意义。
2 结果
2.1 肝癌组织和癌旁组织中ESET mRNA和lncRNA GMDS-AS1表达的影响 与癌旁组织相比,肝癌组织中ESET的mRNA和蛋白表达均显著升高,lncRNA GMDS-AS1的表达显著降低(P<0.05),肝癌组织中lncRNA GMDS-AS1的表达与ESET的表达之间呈负相关性,见表1。THLE-2组相比,HepG2组细胞ESET的mRNA和蛋白表达均显著升高,lncRNA GMDS-AS1表达显著降低(P<0.05),见表2。
表1 ESET mRNA和lncRNA GMDS-AS1在肝癌组织和癌旁组织的表达/
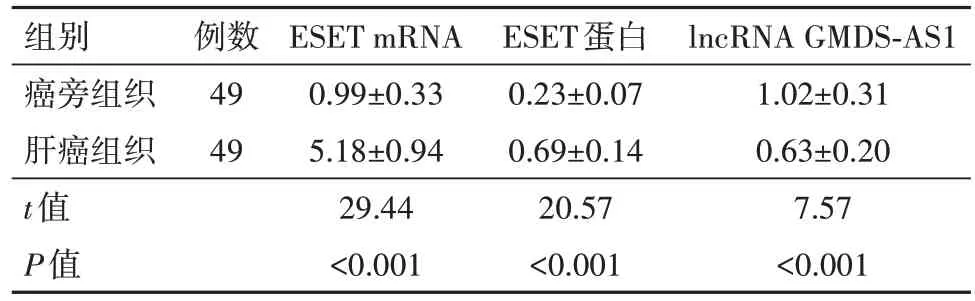
表1 ESET mRNA和lncRNA GMDS-AS1在肝癌组织和癌旁组织的表达/
注:ESET mRNA为组蛋白甲基转移酶集合域分叉1微小RNA,lncRNA GMDS-AS1为长链非编码RNA甘露糖4,6-脱水酶反义RNA1。
lncRNA GMDS-AS1 1.02±0.31 0.63±0.20 7.57<0.001组别癌旁组织肝癌组织t值P值例数49 49 ESET mRNA 0.99±0.33 5.18±0.94 29.44<0.001 ESET蛋白0.23±0.07 0.69±0.14 20.57<0.001
表2 ESET mRNA和lncRNA GMDS-AS1在THLE-2、HepG2中的表达/

表2 ESET mRNA和lncRNA GMDS-AS1在THLE-2、HepG2中的表达/
注:ESET mRNA为组蛋白甲基转移酶集合域分叉1微小RNA,lncRNA GMDS-AS1为长链非编码RNA甘露糖4,6-脱水酶反义RNA1,ESET为组蛋白甲基转移酶集合域分叉1。
lncRNA GMDS-AS1 1.01±0.13 0.51±0.08 9.83<0.001组别THLE-2组HepG2组t值P值重复次数9 9 ESET mRNA 1.00±0.17 4.15±0.32 26.08<0.001 ESET蛋白0.24±0.03 0.67±0.10 12.36<0.001
2.2 敲减ESET抑制肝癌细胞的增殖 与空载体组相比,敲减ESET组细胞ESET的mRNA和蛋白表达均显著降低,Ki-67蛋白表达显著降低,细胞在48、72、96 h的活性显著降低(P<0.05)。见表3。
表3 敲减ESET抑制肝癌细胞增殖/

表3 敲减ESET抑制肝癌细胞增殖/
注:ESET为组蛋白甲基转移酶集合域分叉1,Ki-67为细胞增殖抗原标志物。①与空载体组比较,P<0.05。
组别空白组空载体组敲减ESET组F值P值细胞活性(A490)96 h 0.96±0.10 0.95±0.13 0.77±0.09①8.82 0.001重复次数48 h 0.46±0.08 0.44±0.06 0.32±0.06①11.38<0.001 ESET mRNA 0.98±0.22 0.96±0.03 0.50±0.24①18.62<0.001 ESET 9蛋白0.58±0.07 0.60±0.09 0.24±0.04①75.70<0.001 Ki-67 0.70±0.11 0.69±0.05 0.35±0.11①40.15<0.001 24 h 0.25±0.04 0.26±0.02 0.24±0.03 0.93 0.408 72 h 0.71±0.13 0.68±0.07 0.46±0.07①18.84<0.001 9 9 9
2.3 敲减ESET对肝癌细胞糖酵解和凋亡的影响与空载体组相比,敲减ESET组细胞GLUT-1、LDHA的蛋白表达均显著降低,C-caspase-3蛋白表达显著升高,细胞凋亡率显著升高(P<0.05)。见表4,图1。

图1 敲减组蛋白甲基转移酶集合域分叉1(ESET)的肝癌细胞凋亡图及GLUT-1、LDHA、C-caspase-3的蛋白表达:A为肝癌细胞的凋亡图;B为GLUT-1、LDHA和C-caspase-3的蛋白图
表4 敲减ESET对肝癌细胞糖酵解和凋亡的影响/

表4 敲减ESET对肝癌细胞糖酵解和凋亡的影响/
注:ESET为组蛋白甲基转移酶集合域分叉1,GLUT-1为葡萄糖转运蛋白1,LDHA为乳酸脱氢酶A,C-caspase-3为活化胱天蛋白酶-3。①与空载体组比较,P<0.05。
组别空白组空载体组敲减ESET组F值P值凋亡率/%4.19±0.59 3.74±0.51 14.41±1.5①329.97<0.001重复次数9 9 9 GLUT-1 0.66±0.04 0.64±0.08 0.47±0.10①16.35<0.001 LDHA 0.72±0.13 0.76±0.05 0.43±0.10①29.79<0.001 C-caspase-3 0.25±0.04 0.27±0.02 0.45±0.18①9.53 0.001
2.4 过表达lncRNA GMDS-AS1对肝癌细胞增殖的影响 与空载体组相比,过表达lncRNA GMDSAS1组细胞lncRNA GMDS-AS1的表达显著升高,Ki-67蛋白表达显著降低,在48、72、96 h的活性显著降低(P<0.05)。见表5。
表5 过表达lncRNA GMDS-AS1抑制肝癌细胞的增殖/

表5 过表达lncRNA GMDS-AS1抑制肝癌细胞的增殖/
注:lncRNA GMDS-AS1为长链非编码RNA甘露糖4,6-脱水酶反义RNA1,Ki-67为细胞增殖抗原标志物。①与空载体组比较,P<0.05。
组别空白组空载体组过表达lncRNA GMDS-AS1组F值P值细胞活性(A490)96 h 1.03±0.11 0.98±0.12 0.72±0.08①22.73 0.001重复次数48 h 0.45±0.08 0.46±0.06 0.34±0.06①8.80<0.001 lncRNA GMDS-AS1 1.00±0.14 1.10±0.09 5.19±0.43①725.89<0.001 Ki-67 0.65±0.09 0.63±0.08 0.42±0.06①24.25<0.001 24 h 0.29±0.04 0.28±0.05 0.27±0.03 0.54 0.590 72 h 0.68±0.09 0.65±0.08 0.47±0.07①17.95<0.001 9 9 9
2.5 过表达lncRNA GMDS-AS1对肝癌细胞糖酵解和凋亡的影响 与空载体组相比,过表达lncRNA GMDS-AS1组细胞GLUT-1、LDHA的蛋白表达均显著降低,C-caspase-3蛋白表达显著升高,细胞凋亡率显著升高(P<0.05)。见表6,图2。

图2 过表达长链非编码RNA甘露糖4,6-脱水酶反义RNA1(lncRNA GMDS-AS1)的肝癌细胞凋亡图及GLUT-1、LDHA、C-caspase-3的蛋白表达:A为过表达lncRNA GMDS-AS1的肝癌细胞凋亡图;B为GLUT-1、LDHA、C-caspase-3的蛋白图
表6 过表达lncRNA GMDS-AS1抑制肝癌细胞糖酵解、诱导细胞凋亡/

表6 过表达lncRNA GMDS-AS1抑制肝癌细胞糖酵解、诱导细胞凋亡/
注:lncRNA GMDS-AS1为长链非编码RNA甘露糖4,6-脱水酶反义RNA1,GLUT-1为葡萄糖转运蛋白1,LDHA为乳酸脱氢酶A,C-caspase-3为活化胱天蛋白酶-3。①与空载体组比较,P<0.05。
凋亡率/%3.89±0.84 3.92±0.86 14.67±1.94①200.23<0.001组别空白组空载体组过表达lncRNA GMDS-AS1组F值P值重复次数9 9 9 GLUT-1 0.61±0.04 0.60±0.08 0.42±0.10①17.15<0.001 LDHA 0.74±0.11 0.75±0.09 0.43±0.07①35.61<0.001 C-caspase-3 0.21±0.04 0.23±0.05 0.49±0.08①62.74 0.001
2.6 ESET与lncRNA GMDS-AS1的结合 与空载体组相比,过表达ESET组Ago2抗体的细胞中lncRNA GMDS-AS1表达显著升高(P<0.05)。见表7。
表7 RIP实验结果/

表7 RIP实验结果/
注:RIP为RNA结合蛋白免疫沉淀,lncRNA GMDS-AS1为长链非编码RNA甘露糖4,6-脱水酶反义RNA1,IgG为免疫球蛋白G,Ago2为Argonaute 2蛋白,Input为异染色质,ESET为组蛋白甲基转移酶集合域分叉1。
Input 8.84±0.39 8.79±0.92 0.15 0.883组别空载体组过表达ESET组t值P值重复次数Ago2 2.14±0.21 4.97±0.52 15.14<0.001 lncRNA GMDS-AS1 IgG 0.89±0.10 0.91±0.13 0.37 0.719 9 9
3 讨论
ESET是人类肝癌中表达上调最显著的表观遗传调节因子,ESET的上调与肝癌病人的转移形成、预后较差密切相关[8-9]。近期,Shao等[10]在肝癌的研究中报道,ESET在肝癌组织和细胞中低表达,作为miR-621的下游靶基因调控肝癌的放射敏感性,揭示miR-621/ESET通路通过激活p53信号通路的活性提高肝癌细胞的放射敏感性。本研究发现,ESET在肝癌组织中的表达异常升高,并且敲减ESET具有抑制肝癌细胞增殖、糖酵解,促进凋亡的作用,这均与前人的研究相呼应。但是ESET发挥致癌作用的潜在机制仍需继续研究。此研究还发现了lncRNA GMDS-AS1在肝癌组织中的表达与ESET的表达具有明显的负相关性,于是猜测ESET在肝癌细胞中的作用可能与lncRNA GMDS-AS1有一定关系。
lncRNA已被证明是癌症进展的重要调节因子,包括肝细胞癌[11-12]。lncRNA DEAD/H盒蛋白11反义RNA 1调控肝癌细胞的增殖、迁移、侵袭、糖代谢和凋亡过程[13]。lncRNA远端同源框6反义RNA1(lncRNA DLX6-AS1)的沉默,可抑制肝癌细胞的增殖、迁移和侵袭,促进凋亡的发生[14]。但是,lncRNA GMDS-AS1在肝癌中的作用仍有待进一步研究。Zhao等[15]在肺腺癌的研究中发现新的失调lncRNA,lncRNA GMDS-AS1作为肿瘤抑制基因在肺腺癌中低表达,过表达lncRNA GMDS-AS1能够在体内外抑制癌细胞的增殖并促进凋亡;深入研究还发现,GMDS-AS1是miR-96-5p的靶基因,GMDS-AS1与miR-96-5p相关,调节LUAD细胞的增殖和凋亡,说明lncRNA GMDS-AS1作为肺腺癌治疗的潜在靶点的前景。但是,lncRNA GMDS-AS1作为新发现的癌症调控因子,其在肝癌中的功能尚未有人研究。本研究使用qRT-PCR实验检测了肝癌组织和癌旁组织中lncRNA GMDS-AS1的表达发现,lncRNA GMDS-AS1在肝癌组织中表达下调,这与Zhao在肺腺癌中的研究结果相吻合[16]。进一步研究,通过构建过表达lncRNA GMDS-AS1的肝癌细胞,检测细胞增殖、凋亡、糖酵解发现,过表达lncRNA GMDS-AS1抑制了癌细胞的增殖、糖酵解,促进癌细胞的凋亡,说明lncRNA GMDS-AS1在肝癌中也扮演抑癌基因的角色[17]。此研究还通过RIP实验发现,ESET与lncRNA GMDS-AS1之间存在结合力[18]。
综上所述,甲基转移酶ESET促进肝癌细胞的增殖、糖酵解,抑制凋亡,发挥促进癌症恶化的作用,产生这种作用的潜在机制与负向调节抑癌因子lncRNA GMDS-AS1相关,为肝癌的精准治疗提供新靶点。
猜你喜欢
中南医学科学杂志(2022年6期)2022-12-04
小学生学习指导·低年级(2021年6期)2021-09-10
考试与评价·七年级版(2021年4期)2021-08-14
昆明医科大学学报(2021年4期)2021-07-23
中国循证心血管医学杂志(2020年11期)2020-01-08
小学阅读指南·低年级版(2018年5期)2018-11-02
中华老年口腔医学杂志(2016年4期)2017-01-15
中国医学装备(2016年6期)2016-12-01
中国塑料(2015年10期)2015-10-14
医学研究杂志(2015年4期)2015-06-10
