
不同树龄107杨次生木质部解剖特征及径向变化
2023-11-20 07:59吕瑶刘智郭梦杰
林业与生态科学 2023年4期
吕瑶,刘智,郭梦杰
(河北农业大学 林学院,河北 保定 071000)
木材解剖特性与影响树木生长的内、外因素密切相关,如遗传、形成层年龄和造林措施等[1-3]。导管是树木次生木质部的重要组成部分,对树木水分传导起着关键作用,在树木生长过程中,导管细胞的生长发育与其生理活动关系紧密,其解剖特征对木材性质及其加工性能有重要影响,如会影响木材染色效果、染液渗透等[4-9]。而且,探究导管及其相关解剖特征的径向变化规律,有助于理解树木解剖结构特征与其水分输导能力、木材改性剂与渗透之间的关系[10]。
曾有学者通过研究不同树种次生木质部中导管细胞的解剖结构,来揭示其随生长轮龄增长的径向变化特征。研究结果表明,导管分子径向变化规律在不同木本植物间有一定相似之处,导管在整个树木中,导管细胞直径沿髓心向外随生长轮龄的增加逐渐增加后趋于稳定[11-12]。也有研究表明,不仅在不同树种的种源、家系和无性系间,在相同种源、家系和无性系内,不同株间和株内的径向上的不同部位,木材导管细胞均存在显著的变异性,导管细胞直径、壁厚、导管比量等均随轮龄的增长而变化[13-16]。
关于阔叶树种的导管细胞直径在径向上的分布变化,与其立地条件、气候变化培育措施等因素响应关系是近期研究热点方向之一[17-22]。以往关于针叶树、常绿阔叶树、人工林观光树等树木木质部解剖结构特征径向变化的研究有一定的局限性,如取样部位仅为胸径部位、没有考虑树龄对其径向变异的影响等方面[23-25];同时,对我国北方种植面积最广的主要用材树种107杨,关于不同轮龄木材解剖构造与森林培育之间的关系研究未见报道。本研究利用木材解剖显微测量系统分析了107杨树种4个不同树龄生长轮龄次生木质部中导管细胞解剖结构:早材和晚材的年轮宽度和晚材率、导管细胞径向直径和弦向直径、导管细胞双壁厚和壁腔比、导管比量以及管孔链径向直径和弦向直径,旨在揭示不同树龄107杨次生木质部导管细胞解剖结构径向变异规律,为107杨树的人工林培育技术及木材改性的研究提供一定的参考依据。
1 材料与方法
1.1 研究区概况
试验地位于河北省保定市满城区苗圃基地,地理坐标在E 114°43′20″~115°32′00″,N 38°43′20″~39°07′00″之间,属大陆性季风气候。满城区地处河北省中部,太行山东麓。年平均气温12.9 ℃,日照2 412.7 h,无霜期190 d,平均降水量546.5 mm。
1.2 试验材料
107杨(Populus×euramericanacv. “74/76”)试验样木于2021年11月进行采集,采集方法依据GB1927—2021《木材物理力学实验方法》中相关要求进行,分别选取3年龄(3 a)、5年龄(5 a)、7年龄(7 a)、10年龄(10 a)的107杨,每个相同林龄采伐2株,共8株,每2株试样都尽量控制在同一生长水平,以便各项参数指标测定时,减少误差。样木基本情况见表1。
表1 采样基本信息Table 1 Sample wood information
1.3 试验方法
1.3.1 显微切片的制作 根据木材圆盘南北方向,截取通过髓心的2 cm木条(如图1所示),水煮软化处理后,按年轮分成若干个连续木块。用木材切片机(YD-1508R)将软化好的试材切成厚度16~20 μm的切片,经过0.5%浓度番红染色(99.7,上海浦予工业科技有限公司)、蒸馏水脱色,制成临时切片。将切片放在生物显微镜(型号B204LED,重庆奥特光学仪器有限公司)下进行显微摄影和测量。
(a) 试样截取

(b) 切片制作图1 解剖试样选取Figure 1 Sketch map of wood anatomical properties
1.3.2 年轮宽度的测量 利用游标卡尺测量,沿髓心到韧皮部方向测量年轮总宽度和每年的年轮宽度,测量精度为 0.01 mm。
1.3.3 晚材率计算方法 根据GB/T1930—2009在试件年轮总宽度的范围内,测出每个年轮的晚材宽度,精确到0.01 mm。
(1)
式中:Lw为试件的晚材率,单位为%;∑Lb为测定范围内晚材总宽度,单位为毫米(mm);b为试件测定范围内整年轮总宽度,单位为毫米(mm)。
1.3.4 解剖性质 (1)导管细胞径向直径、弦向直径、双壁厚、壁腔比、管孔链径向直径和弦向直径测量:分别测量3年龄(3 a)、5年龄(5 a)、7年龄(7 a)、10年龄(10 a)4个不同树龄的107杨,各测量部位选在每个年轮中早材和晚材位置,测量导管细胞的大小尺寸。在生物光学显微镜(型号B204LED,重庆奥特光学仪器有限公司)下对其切片样品进行显微拍照,并在被测区域内随机观测导管细胞的相关结构参数(重复次数为10次),利用彩色图像计算机分析系统进行测量。壁腔比是导管细胞双壁厚和导管细胞腔径的比值。(2)导管比量测量:对图像进行“二值化”,通过“反向”“颗粒筛选”“粗化”“颗粒计算”,得到黑色颗粒总面积(导管总面积)占图像总面积的百分比,即导管比量[26]。
1.4 数据处理
利用 Excel 2019软件对木材导管细胞的解剖构造数量特征值进行数据统计,用Origin 2018绘图软件对图形进行绘制,对4个不同树龄107杨次生木质部导管细胞解剖结构随着生长轮龄的变化用多项式进行拟合,R2代表模型拟合的相关系数,用SPSS 27.0进行差异性分析。
2 结果与分析
2.1 年轮宽度与晚材率径向变化
年轮的宽窄是树木径向生长速度的标志,年轮中的晚材率与木材细胞形态、气候因子等密切相关[27]。107杨年轮宽度和晚材率的径向变化,见图2。
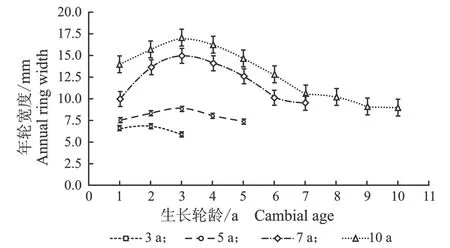
(a) 年轮宽度
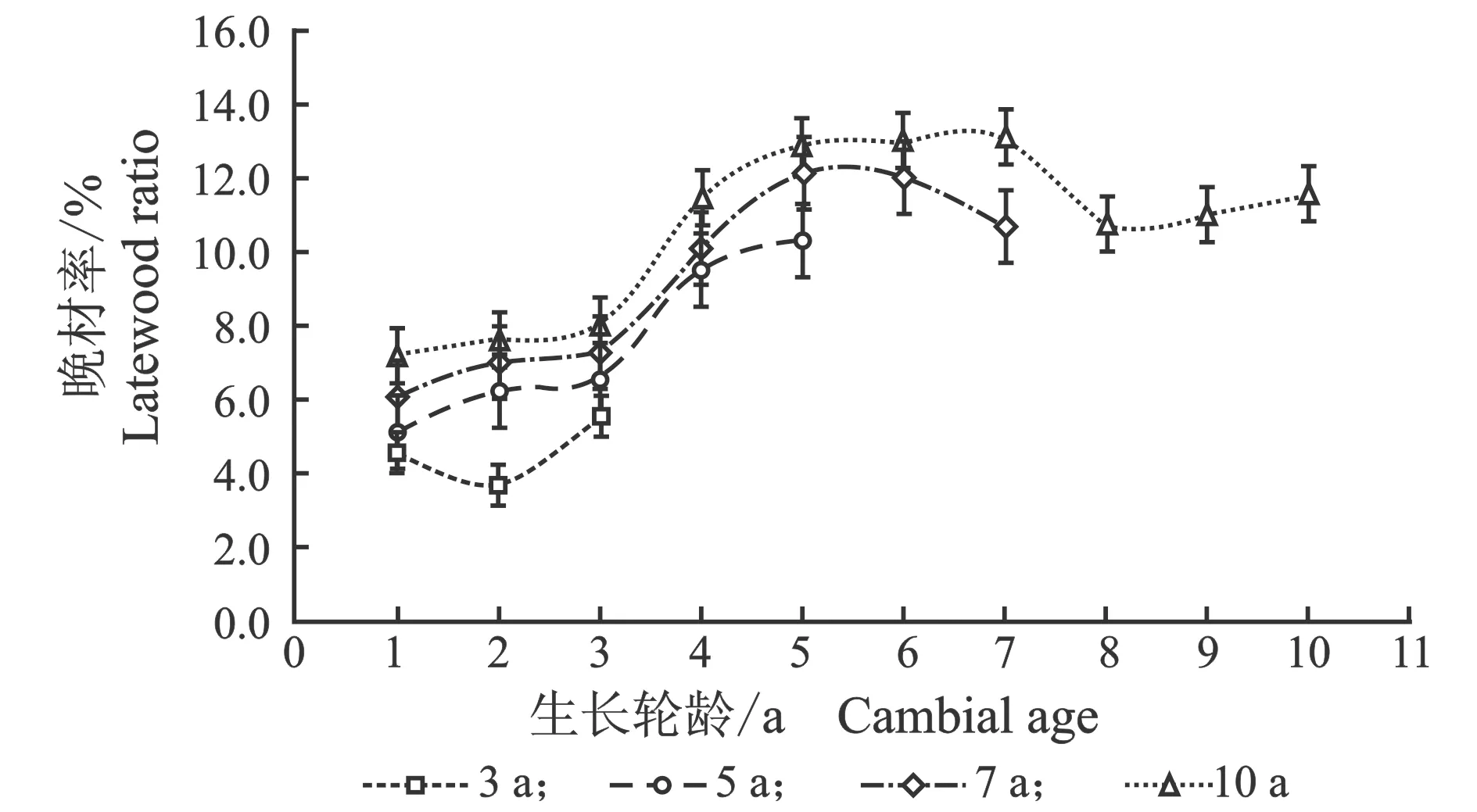
(b) 晚材率图2 107杨年轮宽度和晚材率的径向变异Figure 2 Variation of growth ring width and latewood percentage in the radial direction of Populus ×euramericana cv. “74/76”
由图2可知,3年龄、5年龄、7年龄、10年龄107杨的年轮宽度随生长轮龄增加呈先增大后减小的生长趋势。其中,10年龄107杨年轮宽度最大,3年龄107杨年轮宽度最小。107杨的年轮宽度在7年龄后处于相对稳定状态,表明107杨速生期约3-7年,3年龄、5年龄、7年龄、10年龄107杨的晚材率从髓心向韧皮部逐渐增大,变化规律表现为:10 年龄>7年龄> 5年龄> 3年龄。不同树龄107杨解剖形态特征,见表2。
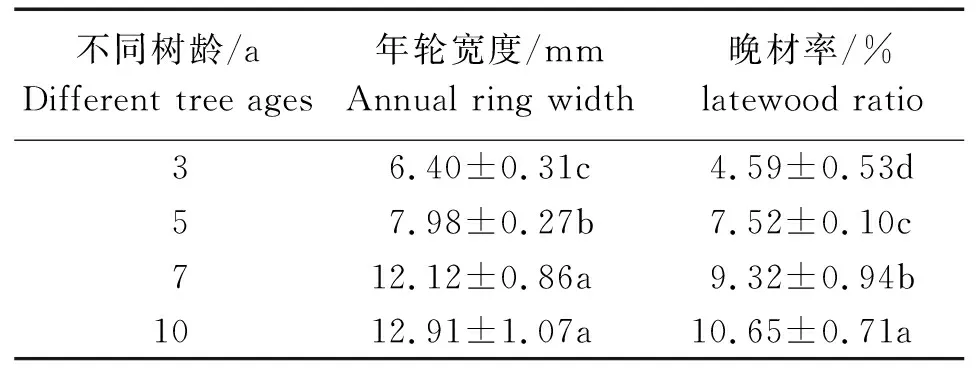
表2 不同树龄107杨解剖形态特征Table 2 Anatomical and morphological characteristics of Populus ×euramericana cv. “74/76” at different tree ages
由表2可知,10年龄107杨年轮宽度最大为12.91 mm,3年龄107杨年轮宽度最小为6.40 mm,标准差范围为0.31~1.07。不同树龄的生长轮龄之间年轮宽度有显著性差异,但7年龄和10年龄的差异不显著。107杨年轮内早材向晚材过渡形式为渐变,晚材率的大小可以作为衡量阔叶树材强度大小的指标,3年龄、5年龄、7年龄、10年龄107杨晚材率变化范围为4.59%~10.65%,材质较轻[28]。方差分析表明,晚材率与不同树龄的生长轮龄之间有显著性差异(P<0.05)。
2.2 导管细胞径向直径和弦向直径径向变化
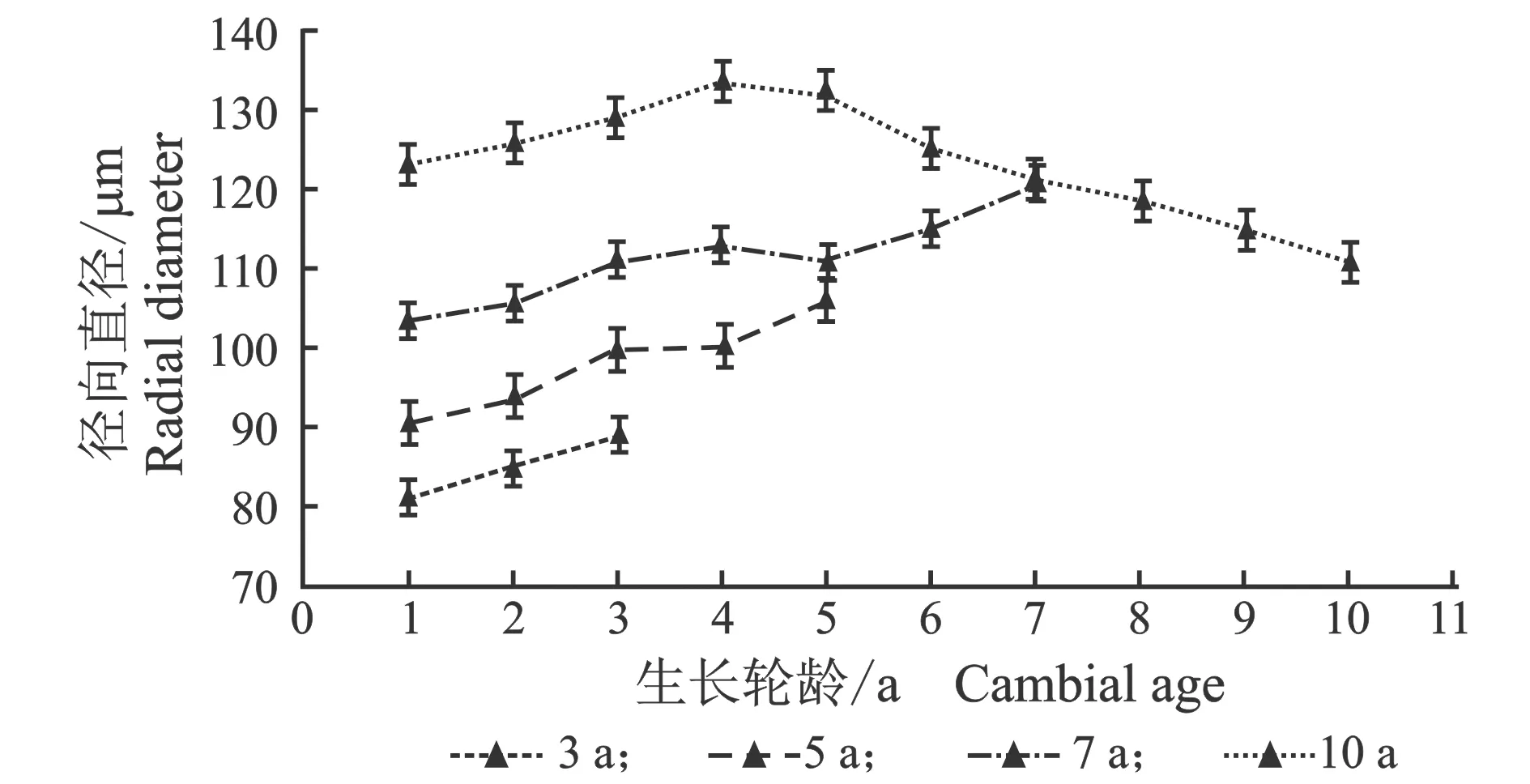
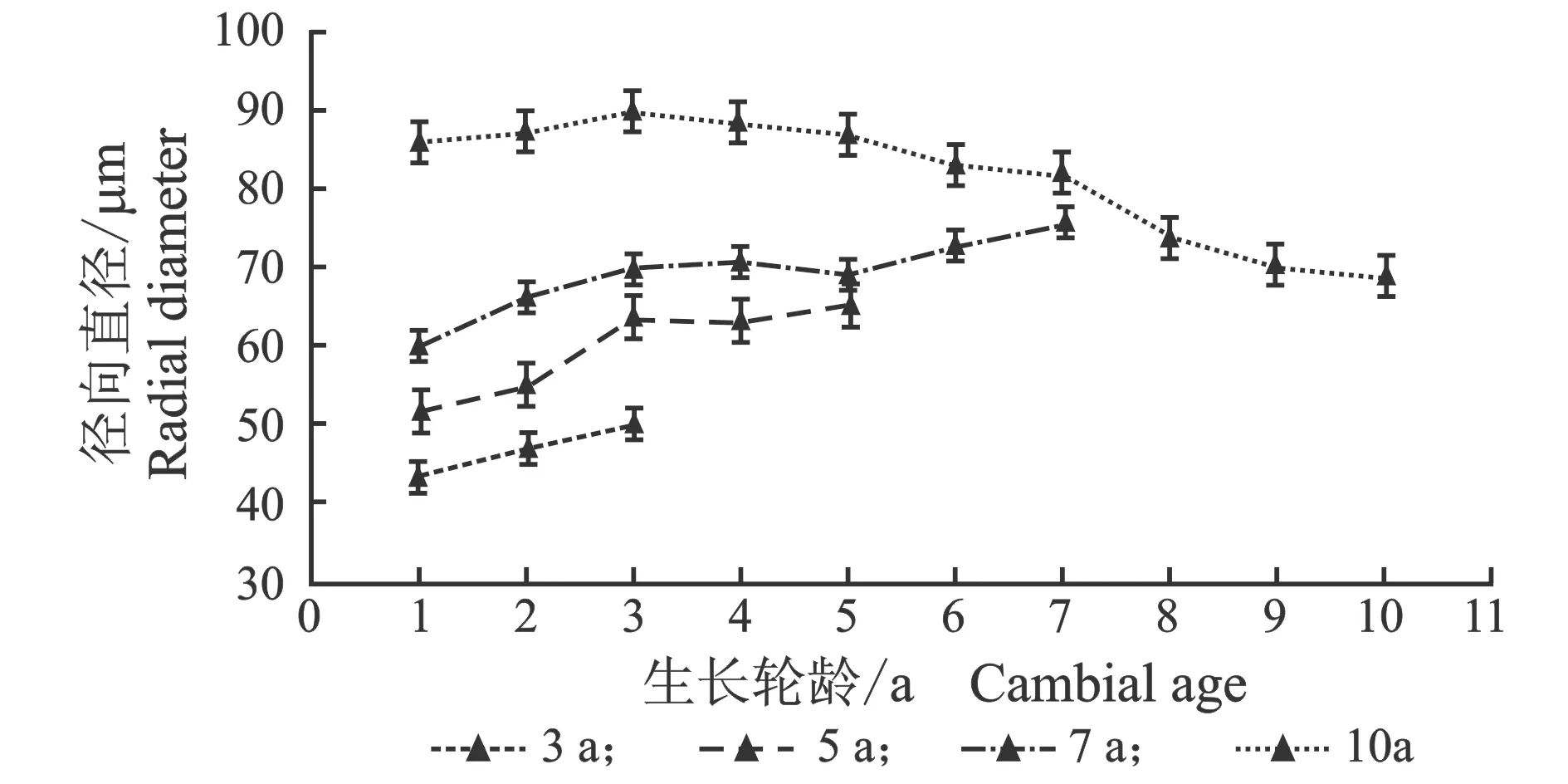
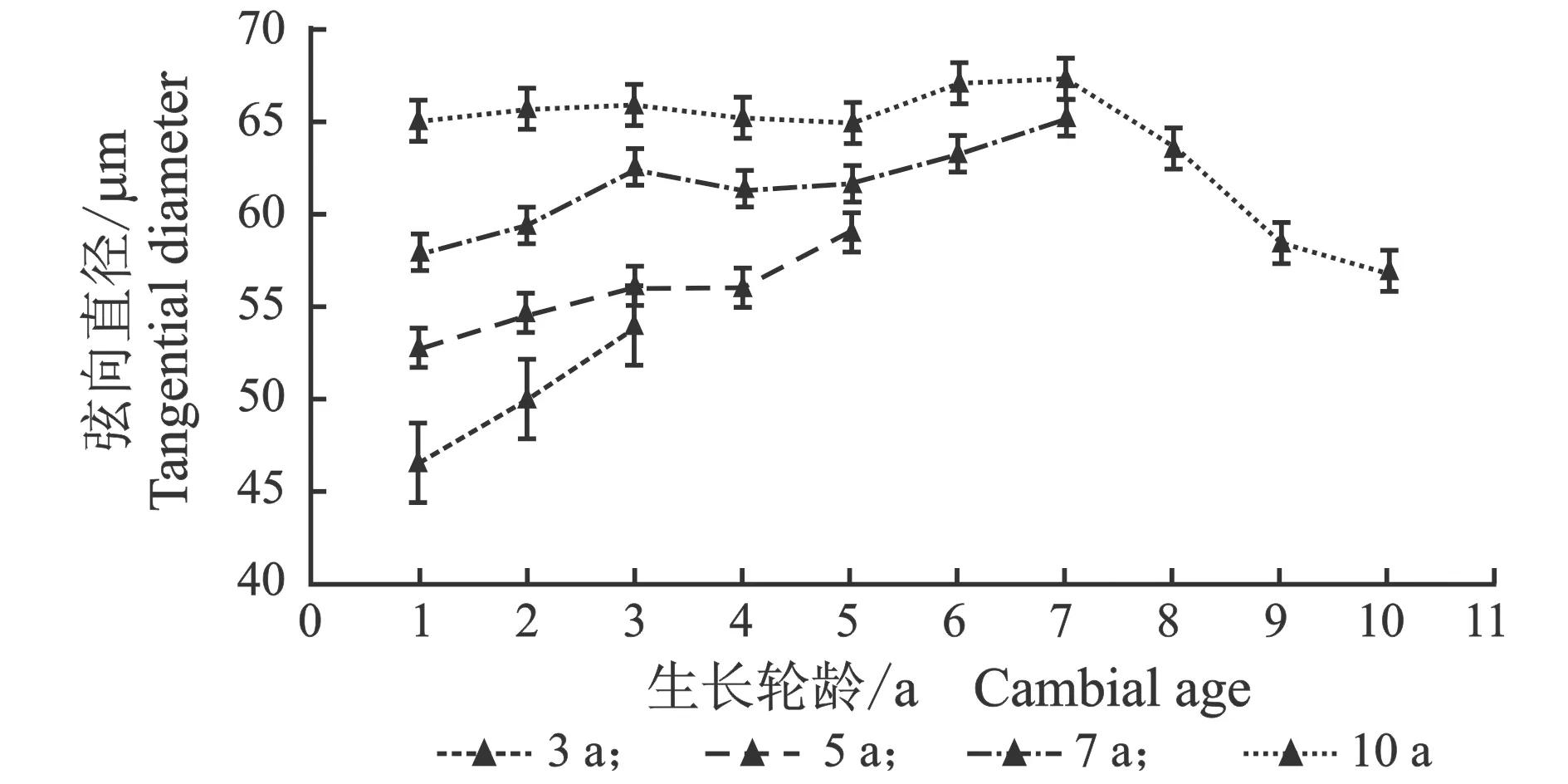
从髓心向韧皮部,早材导管细胞径向直径、弦向直径和晚材导管径向直径、弦向直径变化趋势相似,见图3。
(a) 早材
(b) 晚材
(c) 早材
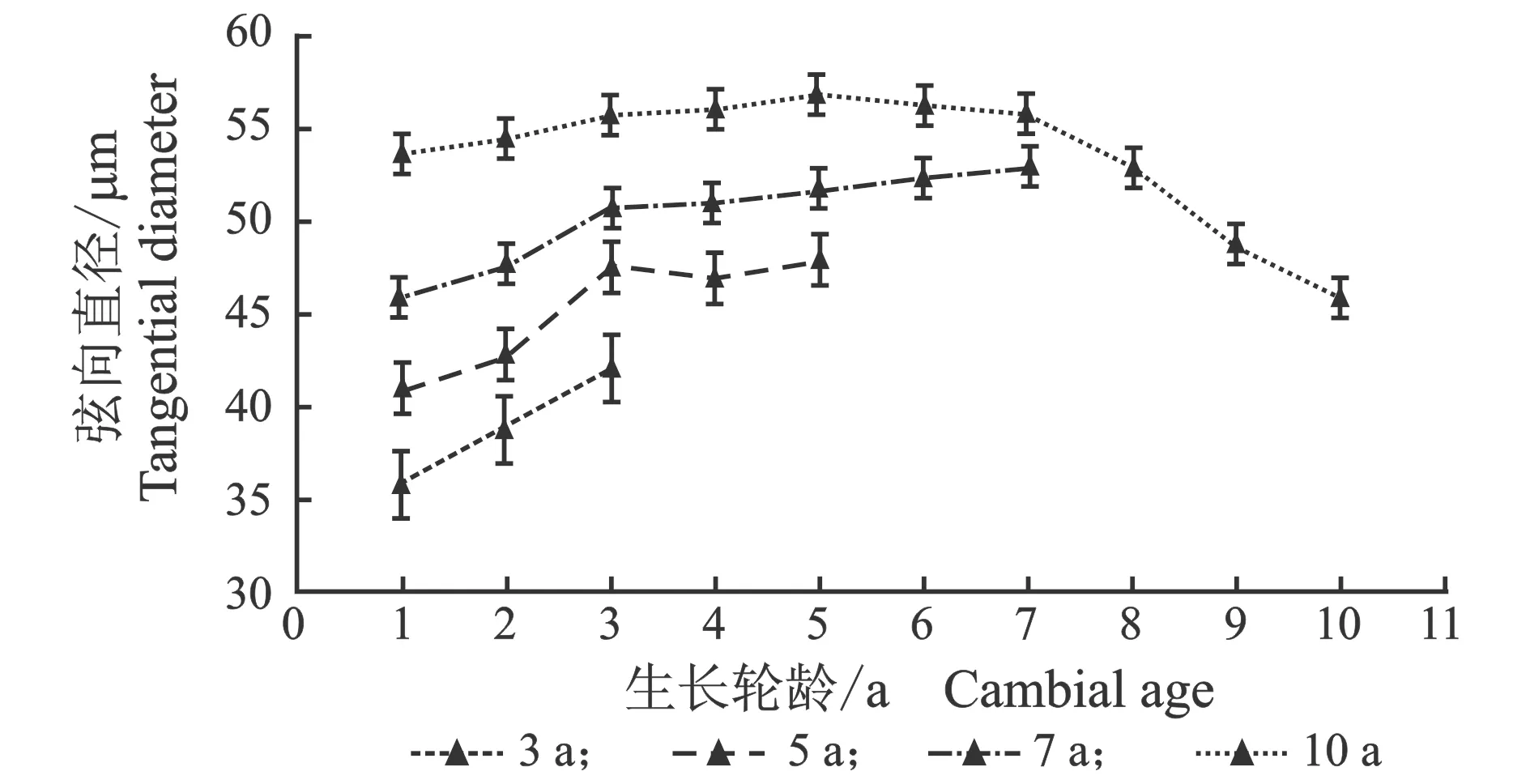
(d) 晚材图3 107杨早材、晚材导管径、弦向直径随生长轮龄的变化Figure 3 Variation of the radial diameter and tangential diameter of the vessel of early and late wood of Populus ×euramericana cv. “74/76” with the annual ring ages
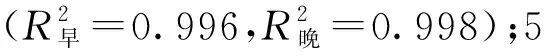
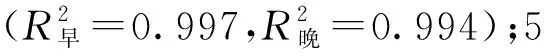
不同树龄107杨早材和晚材导管细胞解剖形态特征,见表3、图4。
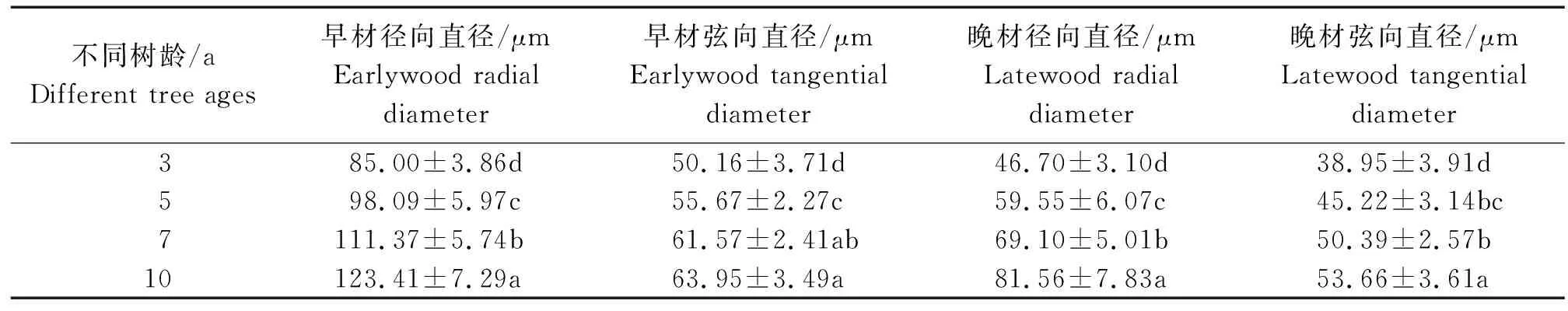
表3 不同树龄107杨早材和晚材导管细胞解剖形态特征Table 3 Anatomical and morphological characteristics of the vessel cell of Populus ×euramericana cv. “74/76” at different tree ages

(a) 3年龄早材 (b) 5年龄早材 (c) 7年龄早材 (d) 10年龄早材

(e) 3年龄晚材 (f) 5年龄晚材 (g) 7年龄晚材 (h) 10年龄晚材图4 107杨不同树龄早、晚材微观构造图片Figure 4 Microstructural pictures of early and late wood of Populus ×euramericana cv. “74/76” at different tree ages
由表3可知,7年龄107杨早材、晚材中导管细胞径向直径平均值分别为111.37、69.10 μm;3年龄、5年龄、10年龄107杨早材、晚材中导管细胞径向直径平均值分别为85.00、46.70 μm;98.09、59.55 μm;123.41、81.56 μm。7年龄107杨早材、晚材中导管细胞弦向直径平均值分别为61.57、50.39 μm;3年龄、5年龄、10年龄107杨早材、晚材中导管细胞弦向直径平均值分别为50.16、38.95 μm;55.67、45.22 μm;63.95、53.66 μm。方差分析表明,3年龄、5年龄、7年龄、10年龄107杨早材、晚材中导管细胞径向直径和弦向直径与生长轮龄均有显著性差异(P<0.05)。结合表3可以更直观地从图4看出,相同生长轮龄时,107杨早材和晚材中导管细胞径向直径和弦向直径变化规律表现为:10年龄>7年龄> 5年龄>3年龄。
2.3 导管细胞双壁厚和壁腔比径向变化
从髓心向韧皮部,导管细胞双壁厚和壁腔比径向变化规律,见图5。
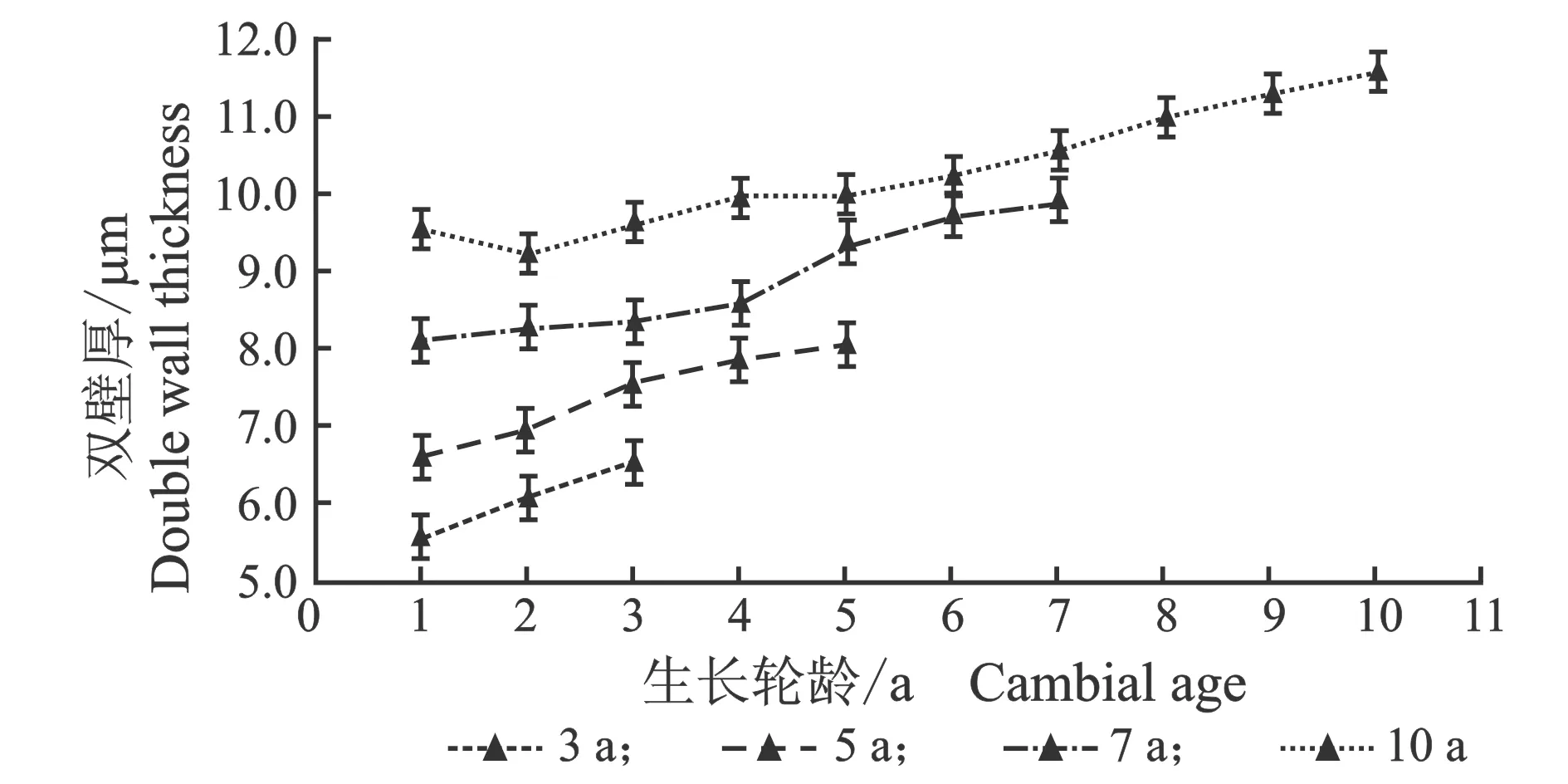
(a) 早材
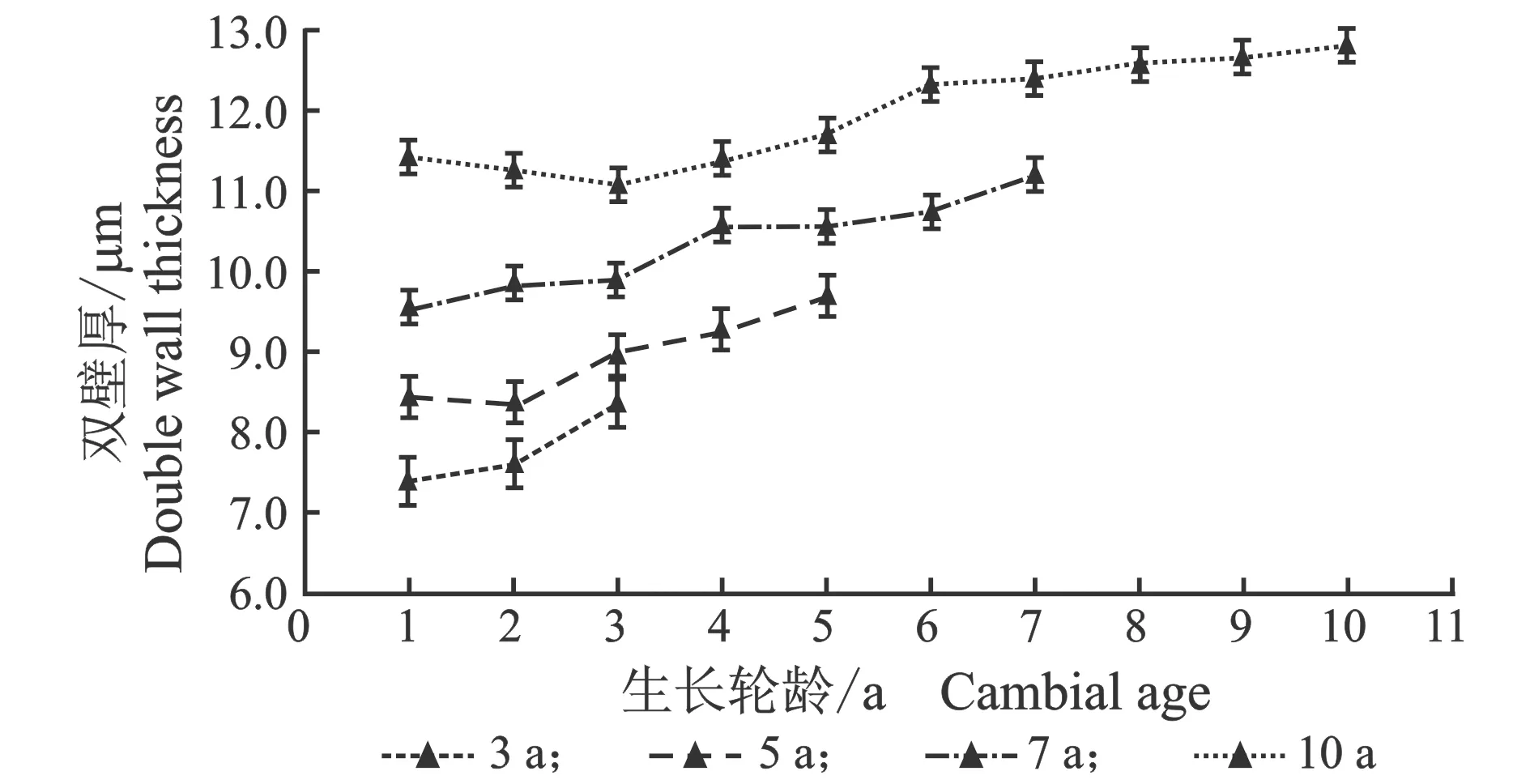
(b) 晚材
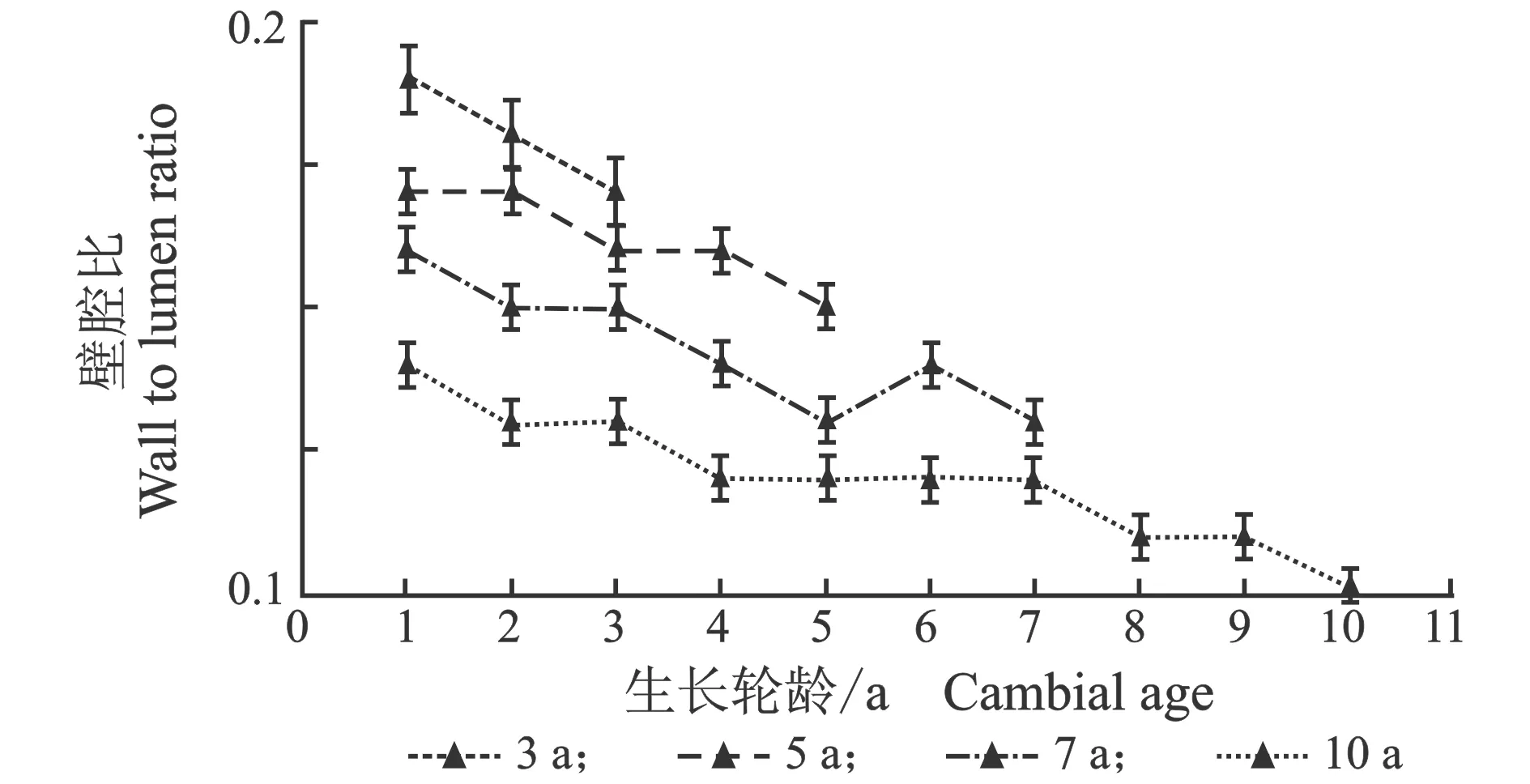
(c) 早材
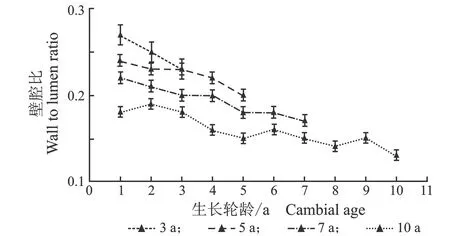
(d) 晚材图5 107杨早材、晚材导管双壁厚、壁腔比随生长轮龄的变化Figure 5 Variation of the double wall thickness and runkel ratio of the vessel of early and late wood of Populus ×euramericana cv. “74/76” with the annual ring ages
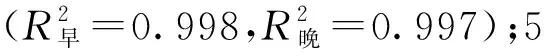
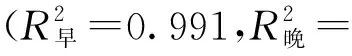
不同树龄107杨早材和晚材导管细胞解剖形态特征,见表4。

表4 不同树龄107杨早材和晚材导管细胞解剖形态特征Table 4 Anatomical and morphological characteristics of the vessel cell of Populus ×euramericana cv. “74/76” at different tree ages
由表4可知,3年龄107杨早材、晚材中导管细胞双壁厚平均值分别为6.08、7.71 μm;5年龄、7年龄、10年龄早材、晚材中导管细胞双壁厚平均值分别为7.40、8.93 μm ;8.89、10.32 μm;10.29、11.95 μm。相同轮龄时,107杨早材和晚材中导管细胞双壁厚变化规律表现为:10年龄>7年龄> 5年龄> 3年龄。方差分析表明,3年龄、5年龄、7年龄、10 年龄107杨早材、晚材中导管细胞双壁厚与生长轮龄之间差异显著(P<0.05)。
5年龄107杨早材、晚材导管细胞壁腔比平均值分别为0.16、0.22;3年龄、7年龄、10年龄107杨早材、晚材导管细胞壁腔比平均值分别为0.18、0.25;0.14、0.20;0.12、0.16。相同轮龄时,107杨早材和晚材导管细胞壁腔比变化规律表现为:3年龄>5年龄> 7年龄> 10年龄,晚材壁腔比大于早材壁腔比。方差分析表明,3年龄、7年龄、5年龄、10 年龄107杨早材、晚材导管细胞壁腔比与生长轮龄之间有显著性差异(P<0.05)。
2.4 导管比量径向变化
从髓心向韧皮部,107杨早材和晚材导管比量的径向变化规律,见图6。
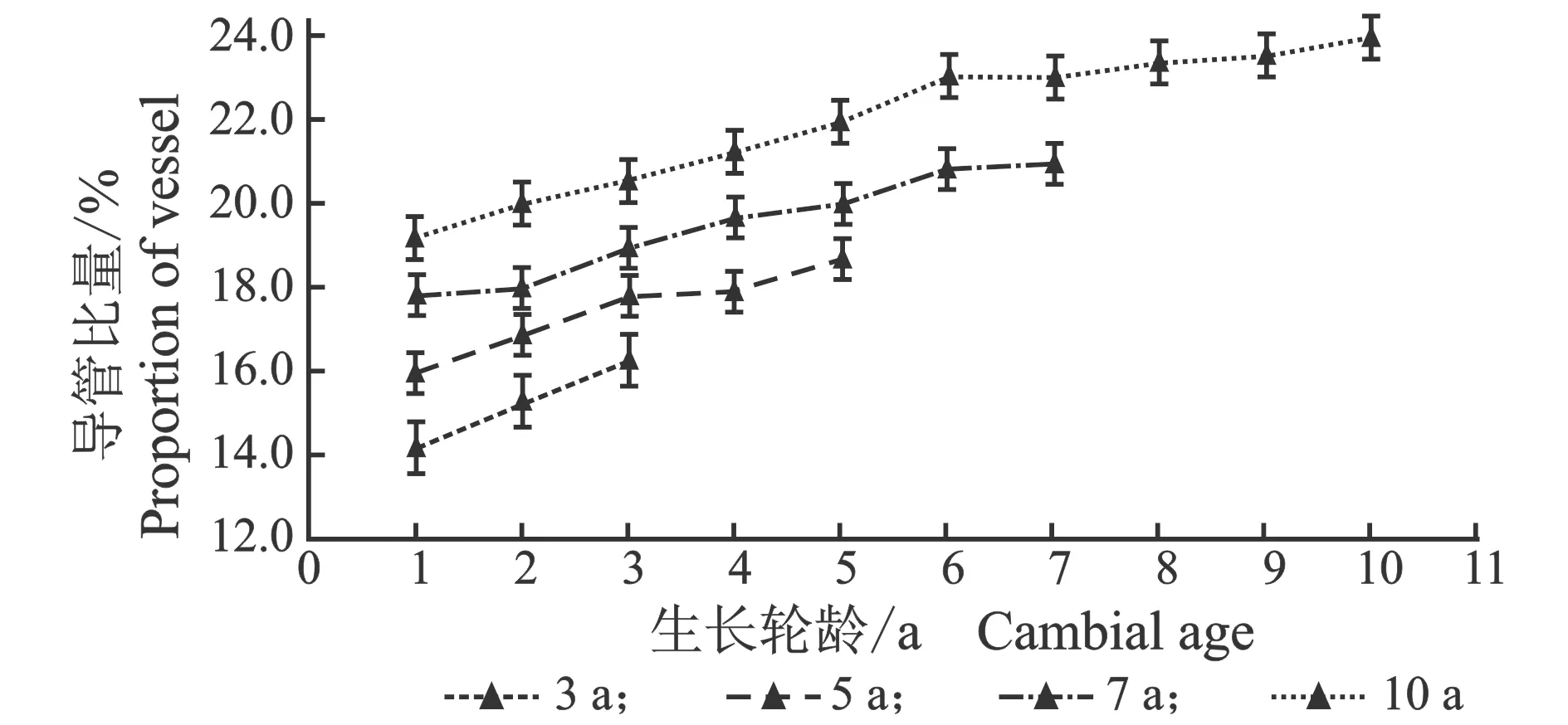
(a) 早材
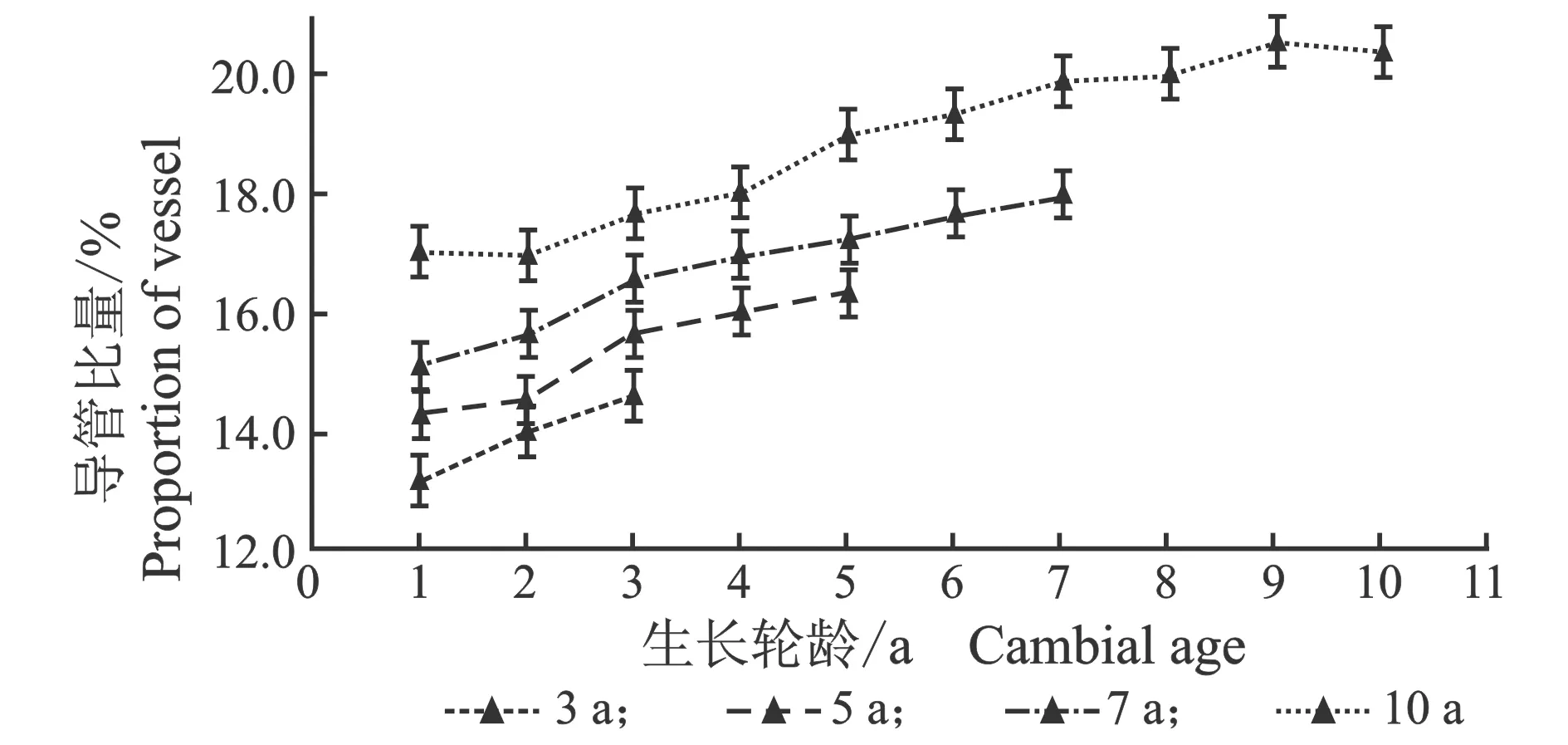
(b) 晚材图6 107杨早材、晚材导管比量比随生长轮龄的变化Figure 6 Variation of the proportion of the vessel of early and late wood of Populus ×
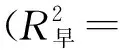
不同树龄107杨早材和晚材导管细胞解剖形态特征,见表5。
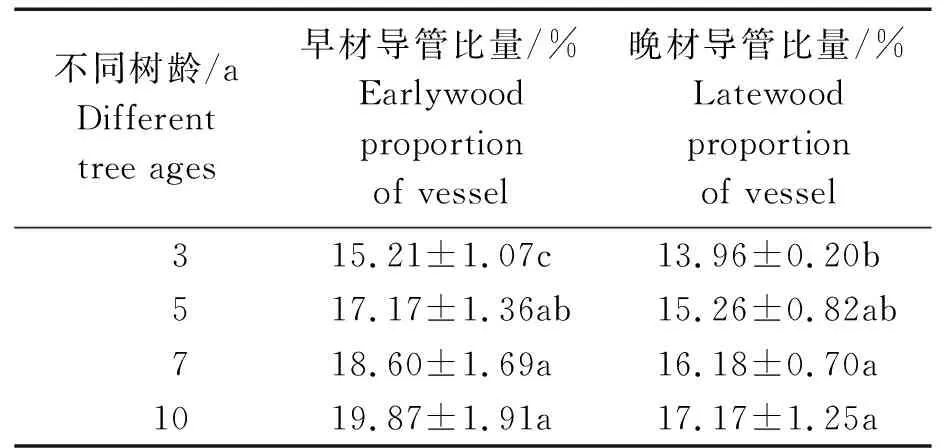
表5 不同树龄107杨早材和晚材导管细胞解剖形态特征Table 5 Anatomical and morphological characteristics of the vessel cell of Populus ×euramericana cv. “74/76” at different tree ages
由表5可知,107杨3年龄、5年龄早材、晚材中导管比量平均值分别为15.21%、13.96%;17.17%、15.26%。7年龄、10年龄早材、晚材中导管比量平均值分别为18.60%、16.18%;19.87%、17.17%。相同轮龄时,107杨早材、晚材中导管比量变化规律表现为:10年龄>7年龄>5年龄>3年龄,早材中导管比量大于晚材导管比量。方差分析表明,3年龄、5年龄、7年龄、10年龄107杨早材、晚材导管比量与生长轮龄之间差异显著(P<0.05)。
2.5 管孔链径向直径和弦向直径的变化
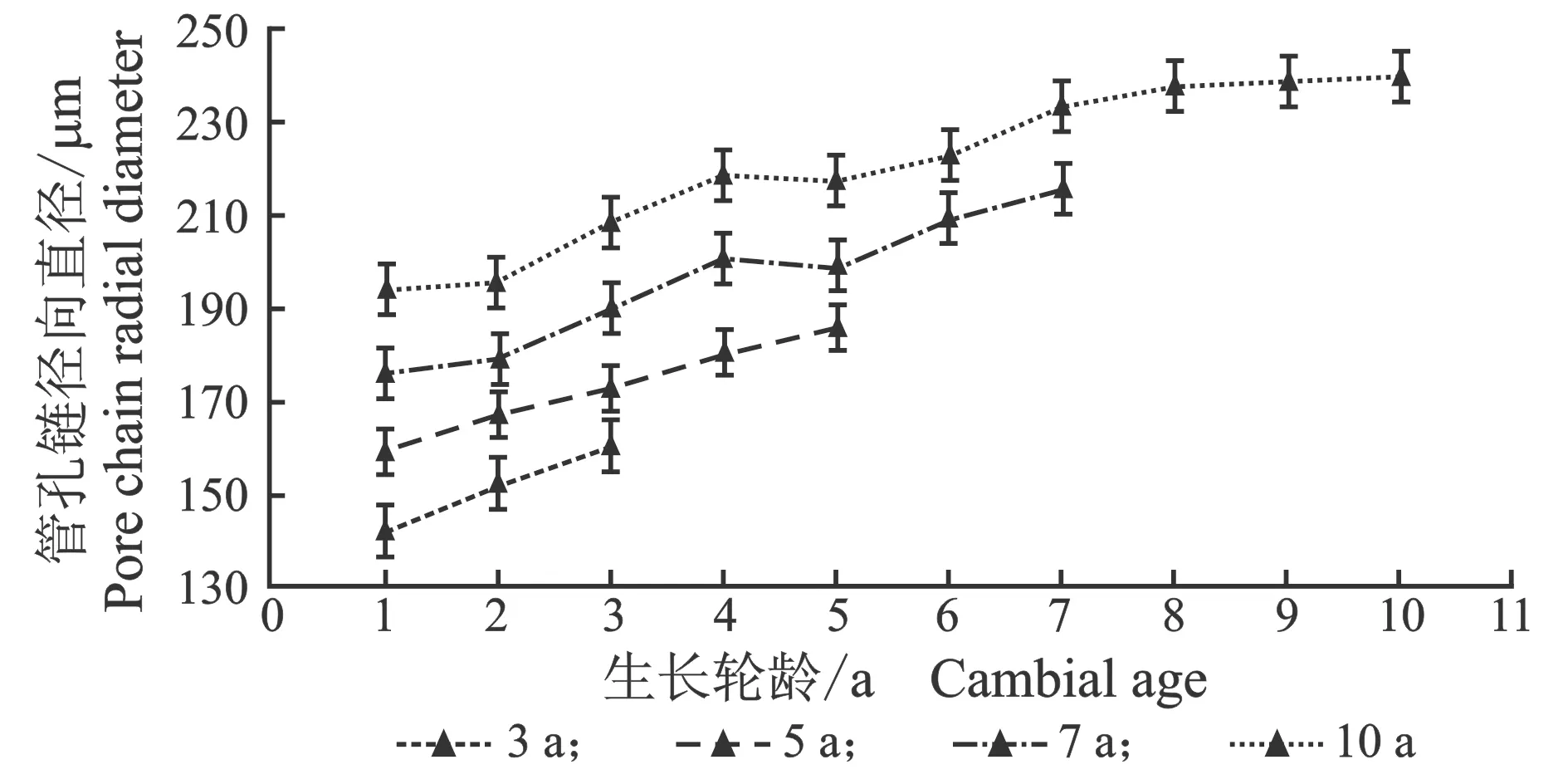
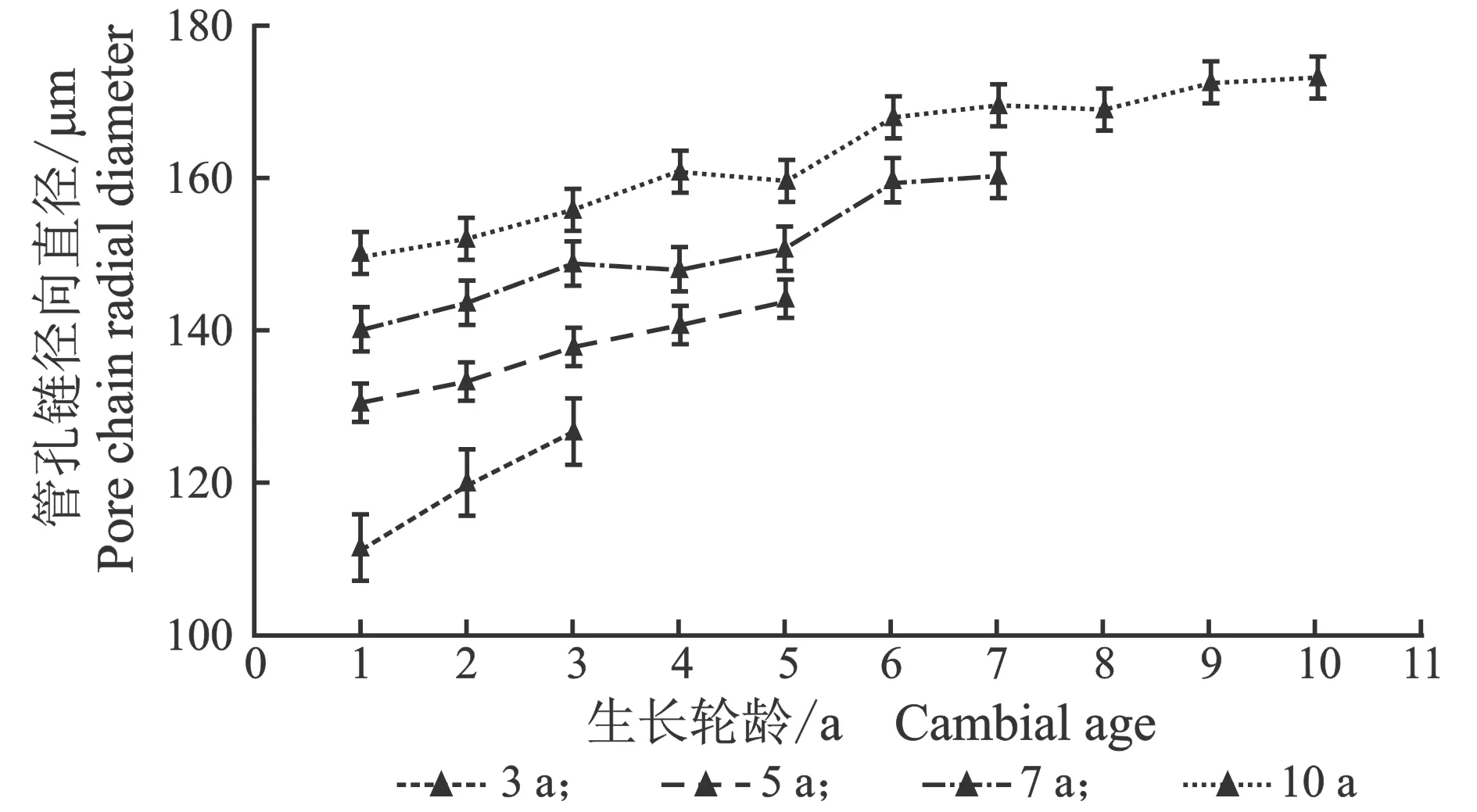
从髓心向韧皮部,107杨早材、晚材中管孔链径向直径和弦向直径的变化趋势,见图7。
(a) 早材
(b) 晚材
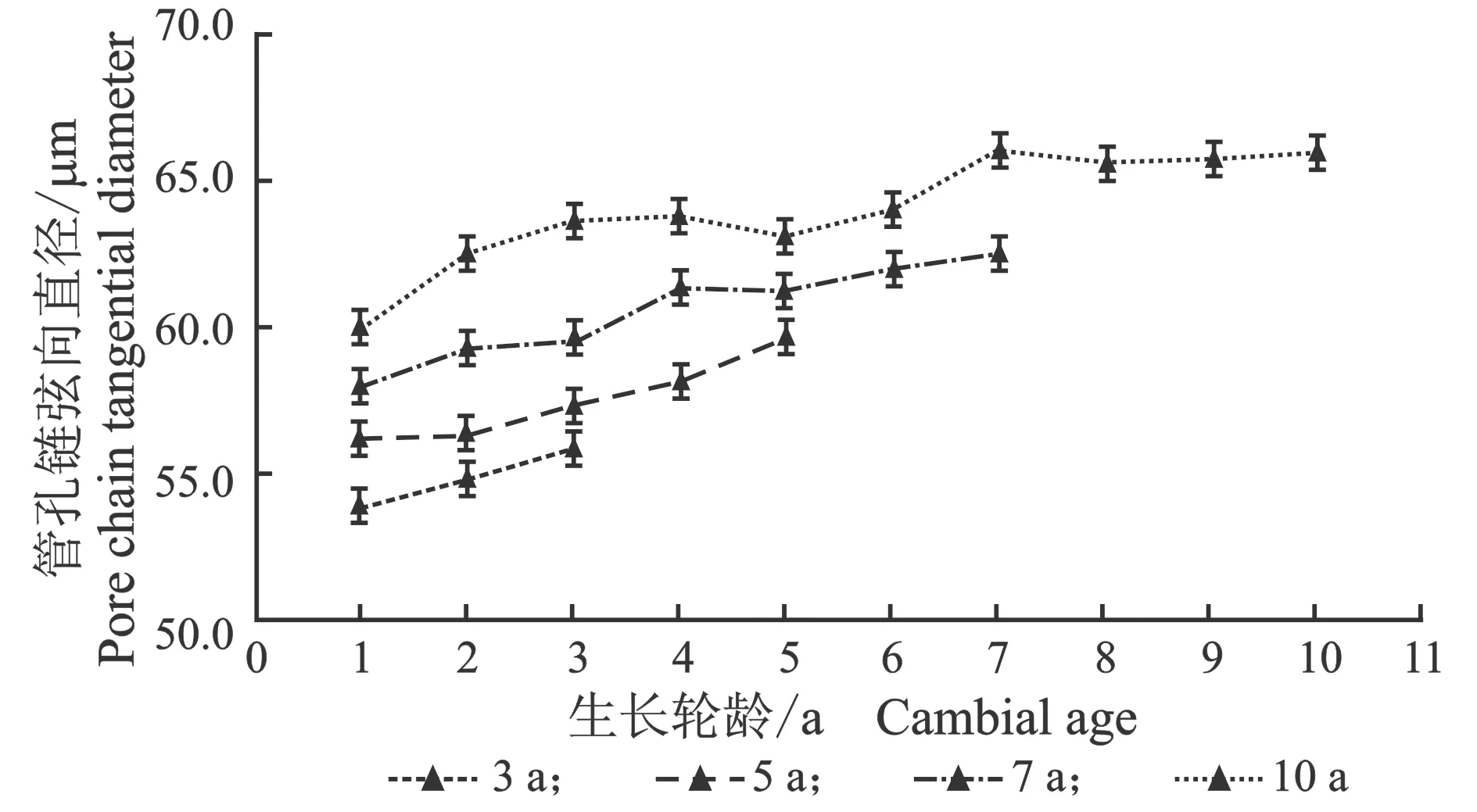
(c) 早材
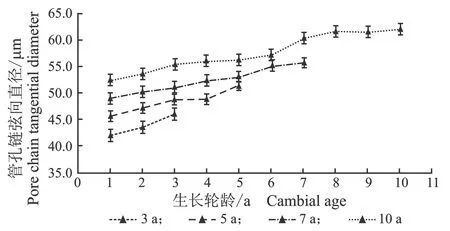
(d) 晚材图7 107杨早材、晚材管孔链径、弦向直径随生长轮龄的变化Figure 7 Variation of the radial diameter and tangential diameter of pipe hole chain of early and late wood of Populus ×euramericana cv. “74/76” with the annual ring ages
不同树龄107杨早材和晚材导管细胞解剖形态特征,见表6、图8。
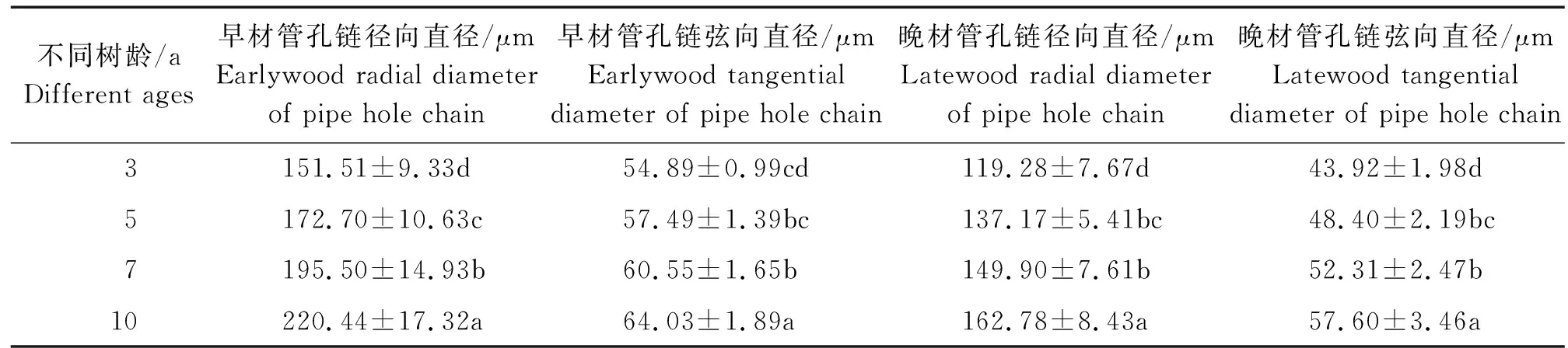
表6 不同树龄107杨早材、晚材导管细胞解剖形态特征Table 6 Anatomical and morphological characteristics of the vessel cell of Populus ×euramericana cv. “74/76” at different tree ages

(a) 3年龄早材 (b) 5年龄早材 (c) 7年龄早材 (d) 10年龄早材
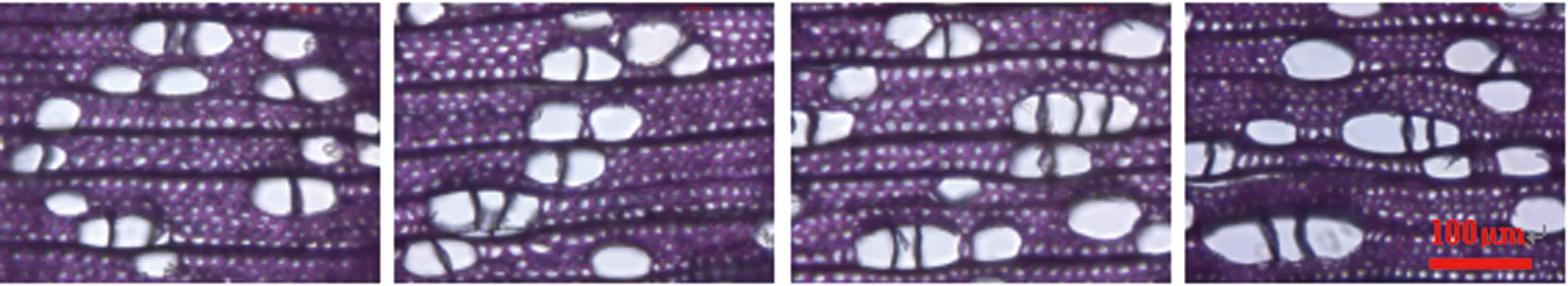
(e) 3年龄晚材 (f) 5年龄晚材 (g) 7年龄晚材 (h) 10年龄晚材图8 107杨不同树龄早、晚材的管孔链显微构造图片Figure 8 Microstructural pictures of pore chain of earlywood and latewood of Populus ×euramericana cv. “74/76” at different ages
由表6可知,3年龄107杨早材、晚材管孔链径向直径平均值分别为151.51、119.28 μm;5年龄、7年龄、10年龄107杨早材、晚材管孔链径向直径平均值分别为172.70、137.17 μm;195.50、149.90 μm ;220.44、162.78 μm。方差分析表明,3年龄、5年龄、7年龄、10年龄 107杨早材、晚材管孔链径向直径与生长轮龄之间差异显著(P<0.05)。
3年龄107杨早材、晚材管孔链弦向直径平均值分别为54.89、43.92 μm;5年龄、7年龄、10年龄107杨早材、晚材管孔链弦向直径平均值分别为57.49、48.40 μm;60.55、52.31 μm;64.03、57.60 μm。方差分析表明,3年龄、5年龄、7年龄、10年龄107杨早材、晚材管孔链弦向直径与生长轮龄之间差异显著(P<0.05)。根据表6和图8可以看出,相同轮龄时107杨中早材和晚材管孔链径向直径和弦向直径变化规律表现为:10年龄>7年龄> 5年龄> 3年龄,早材管孔链径向直径和弦向直径要大于晚材部分的管孔链径向直径和弦向直径。
3 讨论
随着森林培育和加工利用一体化、产业化的需要,林业及林产工业工作者从重视木材数量转向重视木材质量。107杨是我国北方地区重要的速生材和短周期人工林用材树种,其幼龄材与成熟材材质差异显著,对其木材质量相关解剖特征包括年轮宽度和晚材率、导管细胞直径、导管细胞双壁厚、木质部导管比量等的研究具有重要意义[29]。
本研究中,依据4个不同树龄107杨的年轮宽度和晚材率分析可知,107杨年轮宽度能反映出树木生长的速度,总体来看髓心至第3年年轮宽度增长较快,而后逐渐下降,这与石传喜等研究结果基本一致,其原因可能是:导管细胞弦向直径和导管细胞数量受到降雨、温度等气候条件影响所致[11,30]。107杨早材导管细胞腔大壁薄,晚材导管细胞腔小壁厚,晚材率较低为4.67%~8.70%,材质较轻[31]。生长轮龄对导管细胞早材、晚材径向直径和弦向直径有显著影响,其变化规律与赵西平等对大青杨的研究结果一致,与范泽鑫等研究中多个树种导管细胞直径从髓心到树皮逐渐增大的变异规律基本一致[24,32]。但10年龄生长变化有所起伏,前6年轮生长速度较快,7年轮之后呈下降趋势,之后波动较小。这是因为靠近髓心生长轮中的细胞还未成熟,细胞分裂得很快,生长得也很快[33]。当树木达到成熟期时或接近成熟时,细胞相对稳定,分裂缓慢,细胞长度的变化也相对稳定,树木的生长速度下降[34]。微观特征的观察过程中还发现,管孔组合当中管孔链占比较大,早材和晚材管孔链径、弦向直径和导管细胞径向、弦向变化规律相似,这与GUO、姜笑梅等对杨树无性系发现的生长特点相一致[35-37]。3年龄、5年龄、7年龄正处于快速生长期,生长速率快引起导管细胞横向生长速率相对较快,10年龄接近成熟,所以后期生长速度缓慢。而且,KEXIA等也认为导管细胞直径从髓部到树皮逐渐增大,第7年轮之后趋于稳定[38]。
早材和晚材中导管细胞双壁厚随生长轮龄的增加呈缓慢增大的变化规律,与邹明宏等对枫杨的研究结果一致,这种变异特点是形成层原始细胞的分裂和形成层的后生长作用造成的,但与梁善庆等对人工林米老排的研究结果相反,原因可能是所选树种、树种年龄、立地条件等因素所不同造成[39-42]。根据文献报道,不同树龄107杨早材和晚材中导管细胞壁腔比变化幅度和壁腔均比较小,有利于提高制浆得率和质量[43]。早材导管比量变化规律与周亮等对107杨导管比量的变异规律研究结果一致,而且与王嘉楠等对I-69杨、朱莹琦等对楸树、胡进波等对尾巨桉、邹明宏等对枫杨的研究结果也很相似[44-48]。导管比量的径向变异模式从生物学上分析其原因,是次生木质部母细胞分裂所必需的生长因子差异分布造成的结果;树木幼年时期细胞分裂较快,导致导管细胞生长受到外部因素的影响,导管分子的分化程度较弱,导管比量较小;到成熟期之后,细胞的分裂逐渐稳定,良好的细胞迅速扩展,并分化为导管细胞,故导管比量相对较多[44]。
由方差分析可知,不同树龄生长轮龄之间107杨中导管细胞解剖结构径向变化均存在显著性差异,根据这些差异特征和数据,结合用材林木木材微观解剖结构指标要求,可以作为107杨定向培育周期与采伐时间的判断依据[49];本研究结果表明,人工林杨树在幼龄林阶段如果以培育大径材林为目标,最佳轮伐期在10年龄[50];如果培育中、小径材林为目标,最佳轮伐期在7年龄[51]。本研究中,3年龄、5年龄的107杨次生木质部可作纸浆材及中小径民用材;而7年龄、10年龄的107杨次生木质部可作为干径材、板材的基材,细木工板材的芯板等方式加以利用[52-53]。
4 结论
通过对4个不同树龄107杨次生木质部中导管细胞的解剖结构及其径向变化规律进行了研究,表明随着生长轮龄的增加,导管细胞的解剖结构具有比较显著变化规律。
(1)107杨年轮宽度髓心至韧皮部的总体变异规律,是随生长轮龄增加呈先增大后减小的变化趋势,不同树龄年轮宽度的变化幅度为6.40~12.91 mm,生长轮龄对年轮宽度的影响达显著水平(P<0.05)。晚材率从髓心向韧皮部逐渐增大,晚材率较低,变化范围为4.67%~8.70%。3年龄、5年龄、7年龄107杨早材和晚材导管细胞径向、弦向直径随生长轮龄增加逐渐增大,10年龄随生长轮龄增加有先增大后减小的变化趋势(P<0.05)。不同树龄早材、晚材导管细胞径向直径和弦向直径的变化范围分别为85.00~123.41、50.16~63.95 μm;46.70~81.56、38.95~53.66 μm。4个不同树龄107杨年轮宽度、晚材率、早材和晚材导管细胞径向、弦向直径的变化规律表现为:10年龄>7年龄> 5年龄> 3年龄。
(2)3年龄、5年龄、7年龄、10年龄107杨早材和晚材导管细胞的双壁厚随生长轮龄增加逐渐增大,而壁腔比随生长轮龄增加逐渐降低(P<0.05)。不同树龄早材、晚材导管细胞双壁厚和壁腔比的变化范围分别为6.08~10.29 μm、0.12~0.18;7.71~11.95 μm、0.16~0.25。3年龄、5年龄、7年龄、10年龄107杨导管比量随生长轮龄增加呈上升变化的趋势(P<0.05)。不同树龄早材和晚材导管比量的变化范围分别为15.21%~19.87%、13.96%~17.17%。3年龄、5年龄、7年龄、10年龄107杨早材和晚材导管管孔链径向和弦向直径随生长轮龄增加逐渐增大(P<0.05)。不同树龄其导管管孔链细胞径向和弦向直径变化范围分别为151.51~220.44、54.89~64.03 μm;晚材分别为119.28~162.78、43.92~57.60 μm。4个不同树龄107杨早材和晚材中导管细胞的双壁厚、导管比量管孔链径向与弦向直径的变化规律表现为:10年龄 >7年龄> 5年龄> 3年龄;早材、晚材中导管壁腔比的径向变化规律表现为:3年龄>5年龄> 7年龄> 10年龄。
本研究结果揭示了107杨幼龄材次生木质部中导管细胞结构随生长轮龄增长的变异规律,为今后进一步研究107杨树木生长过程中的生理活动规律、确定最佳轮伐期和杨木木材高效、高值化加工利用提供一定的理论参考依据。由于区域限制,只对同一地区、同一树种不同树龄的107杨进行了试验,后续的研究工作应该侧重同一树种不同树龄的107杨在不同地区的解剖特征变异规律;同时,结合水分、温度等气候因子和人工抚育条件对107杨的影响,进一步深入研究其变化规律。
猜你喜欢
红蜻蜓·低年级(2021年12期)2022-01-19
中学生数理化·高一版(2021年3期)2021-06-09
数学物理学报(2021年1期)2021-03-29
重型机械(2020年3期)2020-08-24
果树资源学报(2020年3期)2020-05-13
新疆农业科学(2019年11期)2019-12-31
数学年刊A辑(中文版)(2019年3期)2019-10-08
小天使·一年级语数英综合(2018年7期)2018-09-12
滇池(2017年5期)2017-05-19
天然产物研究与开发(2016年11期)2016-06-15
