
西北印度洋卧蚕-1热液喷口偏顶蛤贝壳的超微结构和矿物组成
2023-11-03 11:38万煌旭刘吉强韩喜球梁金龙周亚东范维佳王叶剑邱中炎孟凡巍
光谱学与光谱分析 2023年11期
万煌旭, 刘吉强, 韩喜球, 梁金龙, 周亚东, 范维佳, 王叶剑, 邱中炎, 孟凡巍
1. 自然资源部第二海洋研究所自然资源部海底科学重点实验室, 浙江 杭州 310012
2. 成都理工大学地球科学学院, 四川 成都 610059
3. 中国矿业大学资源与地球科学学院, 江苏 徐州 221116
引 言
深海热液区自1977年在东太平洋戈拉帕格斯岛洋底被发现以来, 目前已在各大洋发现有上百处[1], 是热液区生物赖以生存的地方, 深海生命绿洲之一。 这些热液区喷口附近因适宜的温度(2~15 ℃)[2]等条件而发育不少数量的贻贝。 由于双壳类生物可以记录其生长环境颇多信息[3], 众多学者已经对生长在淡水[4-5]及浅海[6-9]的贝壳开展了大量的工作。 因样品获取难度高, 关于深海热液区贝壳[10-13]的研究工作还处于探索阶段, 主要集中在贝壳地球化学特征方面[12-13], 而对深海热液区偏顶蛤的壳体内部结构、 矿物组成及成因机制的研究鲜有报道。
长期以来, 对贻贝壳体研究通常利用显微鉴别、 理化鉴别、 扫描电镜等鉴别方法。 最近, 崔楠楠[14]等利用共聚焦显微激光拉曼原位分析技术对不同生长环境下的贻贝壳体矿物组成进行研究, 认为拉曼技术可以区分深海冷泉以及热液区不同理化环境下的贻贝。 张刚生[7]等利用不同壳层文石FTIR光谱位移特征, 表征了生物对不同壳层文石的选择性生物矿化控制作用。 这两种分析手段, 相比传统的显微鉴别、 理化鉴别、 扫描电镜等鉴别方法, 具有独特的优势。 傅里叶转换红外光谱分析是一种快速、 准确、 用样少的方法[4]。 共聚焦显微拉曼光谱是一种具有非侵入性与非破坏性原位光学分析技术[6], 适用于实验室矿物原位无损鉴定[15], 并能够鉴别矿物相(例如, 具CaCO3成分的方解石或文石)[6], 可以快速、 高效地用于研究不同环境下生存的贻贝壳体矿物组成。
采用共聚焦显微激光拉曼原位分析技术和傅里叶转换红外光谱分析技术, 结合扫描电子显微镜观察, 以来自西北印度洋卧蚕-1热液喷口的深海偏顶蛤生物样品(Bathymodiolusmarisindicus)为研究对象, 对其壳体内部的结构与矿物组成开展了分析, 并对这些矿物的成因及其指示意义进行讨论, 以期为揭示海底热液区软体动物壳体形成机制与生物诱导成矿过程提供参考依据。
1 实验部分
1.1 样品制备
样品采集于西北印度洋卧蚕1号热液区。 该热液区以弥散流为特征, 热液生物类群复杂多样。 本工作的深海偏顶蛤样品为蛟龙号载人深潜器第129次下潜获得, 编号为38I-DV129, 水深约3 000 m, 编码为贝(10), 采集出水后, 观察拍照, 用酒精固定以备研究。
游标卡尺测得该深海偏顶蛤的壳长: 80.280 mm; 壳宽: 33.281 mm; 壳高: 37.059 mm。 此外, 深海偏顶蛤壳体内外层(珍珠层与棱柱层)厚度分别为0.1~0.3与0.2~0.4 mm[图1(a)]。
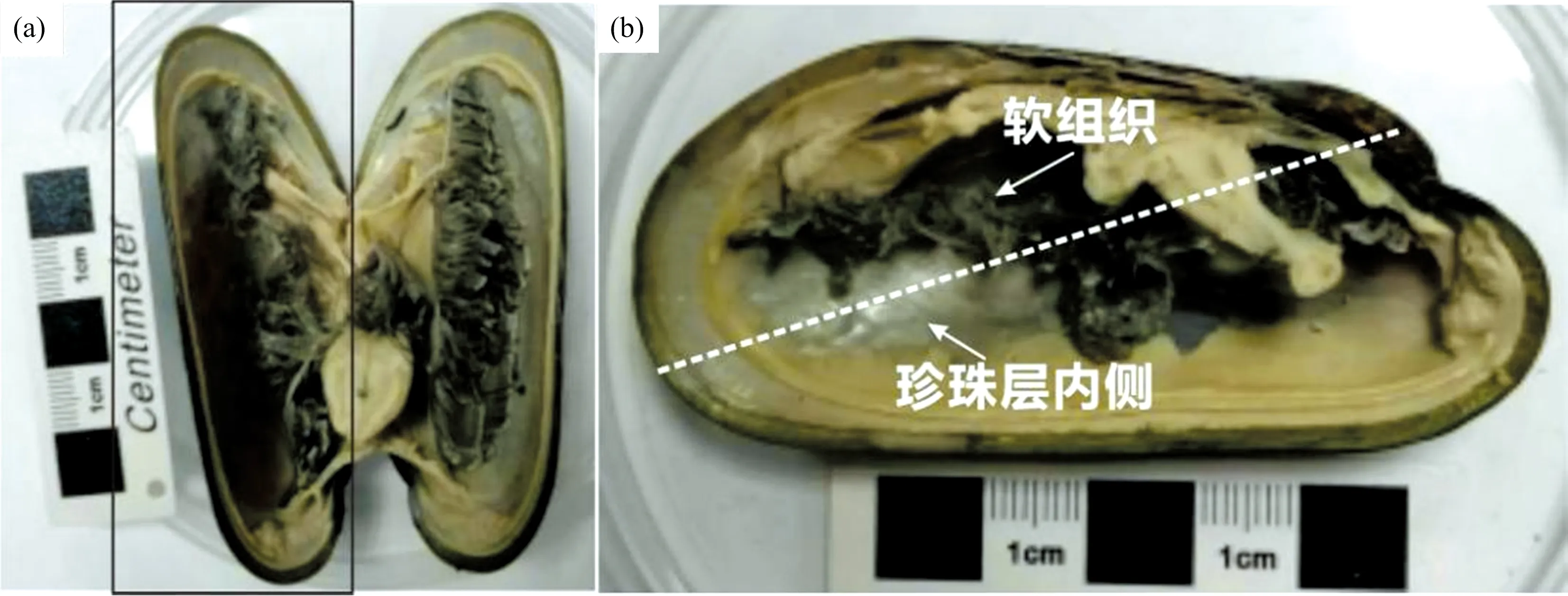
图1 卧蚕-1热液喷口深海偏顶蛤(Bathymodiolus marisindicus)壳体样品照片
实验过程中, 先取出内组织、 清洗风干、 清理后的贝壳样品放置在立方体碟子中, 置于真空玻璃皿中, 用环氧树脂固定24 h。 使用STX-603型金刚石线切割机沿贻贝的最大生长轴进行切割(切片的厚度为2~3 mm)[图1(b)]、 然后进行打磨抛光。 测试前, 用超纯水冲洗样品, 然后使用无水乙醇溶剂进行超声清洗, 最后用超纯水进行冲洗, 晾干。 在体视显微镜下观察贝壳样品切面并拍照。
1.2 方法
1.2.1 扫描电子显微镜(SEM)
深海偏顶蛤的天然断面形貌观察在复纳科学仪器(上海)有限公司完成。 测试仪器为荷兰Phenom生产的Phenom Pharos G2 SEM, 其腔室内部采用背散射电子、 二次电子、 能谱三合一设计结构。 电子源为肖特基场发射, 分辨率优于2.2 nm。 探测元素范围: B(5)—Am(95)号元素。 SEM观测前先将样品置于型号为Q150VES Plus(Quorum)喷金仪器中镀铂金膜2 min。
1.2.2 共聚焦激光拉曼光谱(CLRS)
深海偏顶蛤的壳层矿物成分分析使用自然资源部第二海洋研究所英国Renishaw公司的InVia型激光拉曼光谱仪完成。 系统在使用前利用单晶硅进行仪器校正。 激光波长514 nm, 分辨率1~2 cm-1, 测量范围100~1 500 cm-1, 积分时间10~30 s, 积分次数3次, 激发能量10 mW。
1.2.3 傅里叶转换红外光谱(FTIR)
在自然资源部第二海洋研究所完成, 所用测试仪器为德国Bruker公司生产的配有HYPERION2000型光学显微镜的Vertex80真空型红外光谱仪, KBr分束器, 检测器设置为LN-MCT。 光谱基线校正和峰位标识通过OPUS软件自动处理。 测试条件为室温, 采用反射模式测定, 扫描范围500~4 500 cm-1, 扫描次数128次, 分辨率4 cm-1, 扫描束斑大小为50 μm×50 μm~100 μm×100 μm。
2 结果与讨论
2.1 深海偏顶蛤壳体结构光学及SEM分析
双壳类贝壳分左右壳, 每个单壳包括壳顶、 铰合端及腹缘等[图2(a)], 其纵向和横向的生长方式分别为: 从外层到内层[16]; 从壳顶到腹缘[图2(a)]。
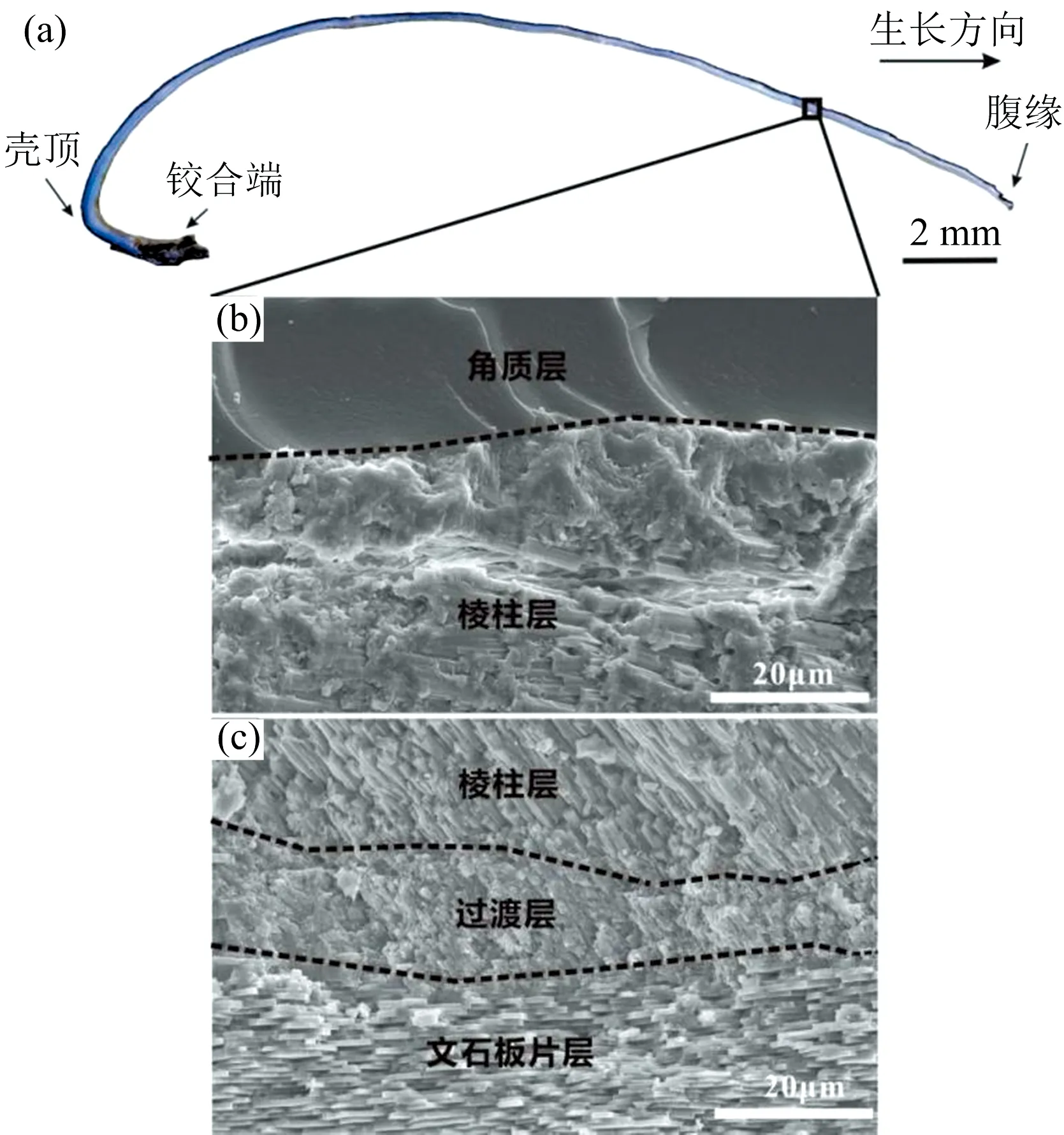
图2 卧蚕-1热液喷口深海偏顶蛤壳体结构分层
本样品B.marisindicus壳体的纵向生长按照其生长方式和内部组成主要可分为五层, 从外到内依次为角质层(有机质层)、 棱柱层、 过渡层、 文石板片层(珍珠层)[图2(b), (c)]及肌棱柱层(见图3)。 图2(c)中黑色虚线区域为从外层向内层的过渡区域, 即为过渡层。 棱柱层由很多呈纤维棱柱状的棱柱晶体构成, 其排列方式一致朝向角质层, 砖块状的文石板片层由平直的板片构成, 层叠生长[图2(b), (c)], 而陈道海和霍颖娴[5]观察研究13种双壳类贝壳微观结构特征发现贝壳棱柱层的晶体形状包括棱柱状、 短柱状、 片状和不规则形状, 珍珠层的晶体形状包括颗粒状、 砖块状、 圆形、 块状和不规则的多边形, 因此卧蚕-1热液喷口深海偏顶蛤B.marisindicus的珍珠层晶体形态与前人研究结果一致, 棱柱层晶体形状存在一定的差别。

图3 卧蚕-1热液喷口深海偏顶蛤壳体天然断面超微结构
据B.marisindicus壳体天然断面的扫描电子显微镜观察结果显示, 天然断面棱柱层棱柱晶体整体排列不规则[图3(b), (c)和(d)], 棱柱间排列紧密, 挤压严重, 导致菱柱体的c轴截面出现不规则状[图3(c), (d)]。 方解石棱柱垂直a轴截面的宽度不均, 约为818~960 nm[图3(b)], 并与文石层呈近45°斜交[图3(a)]。 这种结构与淡水三角帆蚌[17]和淡水粉红贝贝壳[18]的微结构存在显著差异, 后者文石棱柱层近垂直于文石片层。 此外, 邵浩彬等[17]指出棱柱层中小棱柱状晶体间存在少量有机质, 起到黏结作用, 使小棱柱状晶体紧密结合在一起, 引起整体协同效应, 在贝壳受到外力攻击时可以有效地缓解冲击, 从而达到保护内部软体组织的目的。 因此, 小棱柱状晶体整体协同效应结合方解石棱柱层斜交于文石板片层可更好地缓冲外界掠食者对贝壳的攻击。
另外, 卧蚕-1热液喷口深海偏顶蛤壳体棱柱层出现交错现象[图3(e)], 图左侧方解石棱柱体c轴向外部(角质层)生长, 图右侧棱柱体c轴向内部(珍珠层)生长。 欧晓娅等[19]研究发现浅海砗磲的文石棱柱体存在近120°相交的“编织状”交错结构, 但并未指明导致此现象的原因, 而是指出此现象可导致砗磲不同壳层的透明度不同, 并对砗磲壳层的成色产生一定影响。 在自然界中, 生物往往会进化出独特的生物结构或特殊的生物功能以便生存和适应周围的环境[20]。 软体动物的壳体通常具有单一或几种微观结构, 这些结构相互配合, 提供一系列复杂的多功能特性[21], 进而适应周围的水环境并存活。 然而, Génio等[22]却指出, 与喷口和渗漏处的腹足类动物一样[23], 化学共生贻贝壳体结构反映了它们所属的系统发育类群的结构, 也许不能代表它们对所栖息的极端环境或一般深海环境的适应, 且Bathymodiolus物种的壳体超微结构中纤维状棱柱体整体显示由水平到强烈倾斜的特征。 此外, Machado等[10]研究发现一定静水压力下的浅淡水贻贝壳体棱柱层和珍珠层的晶体和排列方式均发生相应的变化, 珍珠层变化较为强烈, 与深海热液喷口贻贝壳体结构存在相似之处, 从而得知贻贝壳体结构受深海静水压力的影响。 Ca2+可用性也可影响深海热液喷口贻贝壳层微观结构特征[17]。 控制壳体微观结构沉积和演化特征的机制还远未被完全了解[34], 在物种进化的不同时期, 不同的选择压力可能在不同的调控水平上起作用[22]。 因此, 我们认为本研究的棱柱层与珍珠层近斜交及棱柱层交错现象主要受贝壳遗传因素的影响。 具体原因有待进一步研究证实。
卧蚕-1热液喷口深海偏顶蛤壳体的过渡层形状极不规则, 它延续了棱柱层的生长取向, 既具有棱柱状形态也具有不规则的片状特征, 整体显示从棱柱状到文石板片状过渡的趋势, 无法估计其晶体厚度[图3(f)]。 印度洋深海偏顶蛤B.marisindicus的壳内层包括珍珠层和肌棱柱层。 从图3(g)和(h)中可观察到文石层具有片状结构, 即砖墙结构。 贝壳珍珠层中有机基质与文石板片组成了贝壳的“砖-泥”结构模式, 其中碳酸钙板片作为砖, 而在板片之间起粘结作用的有机质为泥[25]。 此外, 本研究B.marisindicus的壳体珍珠层微观结构与大西洋中脊(MAR)深海热液喷口收集的贻贝B.azoricus[26]壳珍珠层微观结构相似。 然而, 梁艳等[27]研究发现腹足纲香螺贝壳的文石层由多级结构构成, 不同于双壳贝类和鲍鱼文石层的“砖墙式”结构。 因此, 不同种属贝壳之间的文石层结构存在较大差异。 深海偏顶蛤B.marisindicus的文石形态较为规则、 平直, 为成熟的文石板片。 文石板片呈堆叠生长, 厚度不均[图3(h)], 极薄片层约为205 nm, 极厚片层约1 260 nm, 同一区域的文石板片厚度相似, 不同区域的厚度相差较大。 前人也曾在淡水贝壳珍珠层中发现片状文石晶体厚度不均的现象[17, 19], 此现象可能受贝壳自身的生理因素控制, 其文石板片厚度根据生长环境的变化而有所改变。 从图3(i)和(j)中可观察到肌棱柱层具有简单的棱柱状微观结构, 棱柱层和珍珠层均覆盖在其之上。 在深海热液喷口贻贝[22]中均可发现文石型晶体构型的肌棱柱层。
综上所述, 不同种属的贝壳以及生长于不同环境的贻贝, 其壳体棱柱层和珍珠层的微观结构特征均存在显著差异。
2.2 深海偏顶蛤壳层矿物组成分析
2.2.1 激光拉曼光谱分析
拉曼光谱分析显示, 深海偏顶蛤壳体外层拉曼谱线的特征谱峰为282、 713和1 086 cm-1(图4), 深海偏顶蛤壳体内层拉曼谱线的特征谱峰为155、 207、 704和1 086 cm-1(图4)。 方解石和文石晶体结构的拉曼特征峰处于~1 086 cm-1为碳酸盐(CO3)对称拉伸(ν1振动), 300 cm-1以下的波段为晶格模式, 700 cm-1附近的波段为CO3的面内弯曲(ν4振动)[28]。 数据与前人研究结果一致(表1),B.marisindicus的壳体成分为碳酸钙(CaCO3), 壳体珍珠层和棱柱层分别为文石和方解石。 此外, 贝壳的矿物成分可完全为文石质或方解石质, 也可在单独的单矿物层中同时含有文石和方解石[29]。 李青梅等[30]指出浅海翡翠贻贝贝壳棱柱层、 珍珠层及其过渡区域均由文石组成。 淡水三角帆蚌的棱柱层和珍珠层均为文石型碳酸钙[18]。 浅海扇贝和珍珠母贝的碳酸钙成分分别为方解石及方解石和文石[31]。 因此, 不同种属的贝壳壳体包含不同的矿物成分。
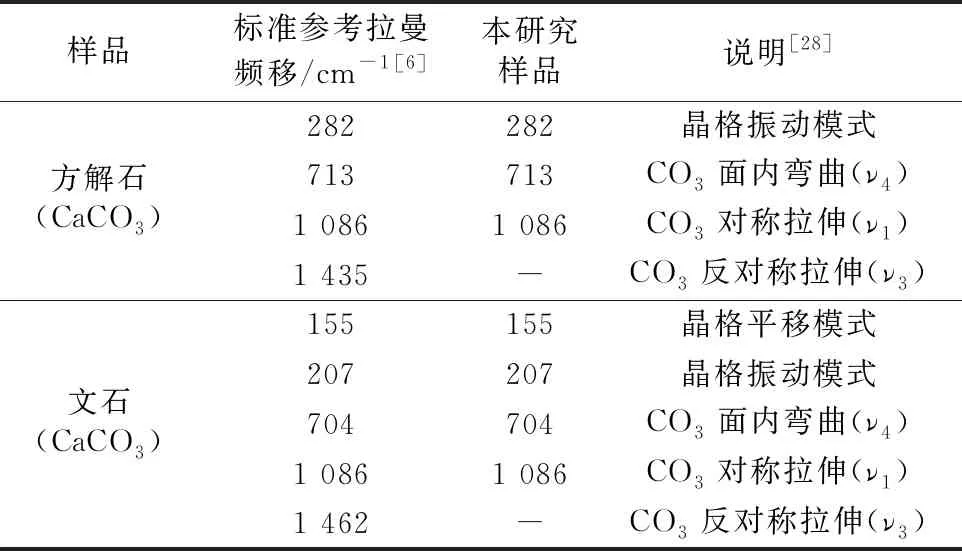
表1 本研究样品与碳酸盐矿物标准拉曼光谱
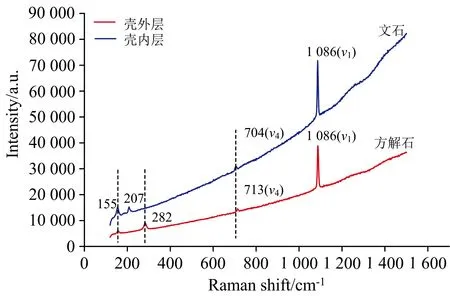
图4 卧蚕-1热液喷口深海偏顶蛤壳内外层激光拉曼光谱图
激光拉曼光谱技术在室内和户外(远洋深海、 野外露头)均可开展原位测试。 Xi[32]等在对南海冷泉碳酸盐岩的激光拉曼光谱原位探测时发现, 冷泉生物群落密集分布区和无生物覆盖区内都普遍存在文石(207、 703和1 086 cm-1)和方解石(281、 713和1 086 cm-1), 方解石随着冷泉系统的变化较稳定, 文石晶体结构却遭受明显破坏, 由此认为自生碳酸盐岩的原位拉曼光谱数据可以反映冷泉系统的演化过程。 此外, 在同一激发能量和曝光时间下, 从拉曼谱峰的强度和半高宽可观察到, 卧蚕-1热液喷口深海偏顶蛤壳体文石的结晶度略高于方解石。 对这一现象解释有两种, 其一, 由于深海偏顶蛤的纵向生长方式是从外层向内层, 故内层文石层受保护度远高于外层方解石层; 其二, 在动力学方面, 文石的力学性能优于方解石[14], 因此文石层受周围海水环境的影响相对较小。
2.2.2 傅里叶转换红外光谱(FTIR)分析
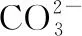
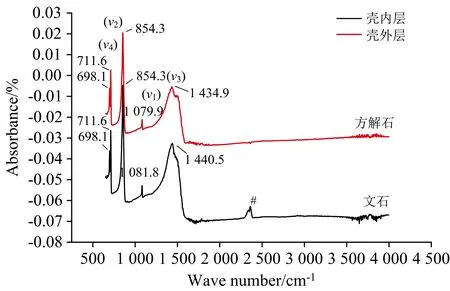
图5 卧蚕-1热液喷口深海偏顶蛤壳内外层傅里叶红外光谱图#有机质引起的衍射峰
本文深海偏顶蛤壳外层方解石的相对强度之比(Iν2/Iν4b)为6.7(表2), 且壳外层方解石的各项参数与无机方解石所知参数相差甚远。 无机成因方解石的Iν2/Iν4b为3.0左右[34]。 因此, 本研究深海偏顶蛤壳体方解石的Iν2/Iν4b远大于无机成因方解石的Iν2/Iν4b, 由此可知西北印度洋卧蚕-1热液区深海偏顶蛤壳外层方解石为生物成因(有机成因), 外层形成受严格的生物控制作用, 且生物矿化控制程度极高。 然而, 本研究深海偏顶蛤珍珠层文石ν2带振动频率值为854.3 cm-1, 且ν1、ν3及ν4带的频率值(1 081.8、 1 440.5、 698.1及711.6 cm-1)与无机文石的频率值(表2)非常接近。 张刚生和李浩璇[35]对16种贝壳生物成因文石的FTIR光谱进行测量发现文石的ν2带可作为指纹带鉴别生物和无机成因文石, 其中生物和无机成因文石ν2带频率平均值分别为863.4和855.5 cm-1。 因此, 本研究的印度洋深海偏顶蛤的珍珠层文石为无机成因。 此外, 深海偏顶蛤B.marisindicus的壳体珍珠层存在有机质。 贝壳珍珠层中含有约95%的文石碳酸钙和约5%的蛋白质、 多糖等大分子物质[36]。 因此, 深海双壳类贝壳壳体是一种潜在的研究生物矿化过程中有机质与无机质材料复合机制的优良载体。

表2 深海偏顶蛤壳内外层与无机碳酸盐矿物碳酸根的振动红外吸收频率(cm-1)
3 结 论
通过SEM、 激光拉曼和傅里叶转换红外光谱分析了西北印度洋卧蚕-1热液喷口深海偏顶蛤(Bathymodiolusmarisindicus)壳体天然断面的形貌与矿物组成, 结果表明:
(1)印度洋深海偏顶蛤(B.marisindicus)壳体的纵向生长从外到内依次为角质层(有机质层)、 方解石棱柱层、 过渡层、 文石板片层(珍珠层)及肌棱柱层。
(2)深海偏顶蛤壳体棱柱层棱柱晶体呈纤维状, 排列紧密, 挤压严重, 导致棱柱体的c轴截面出现不规则状, 方解石棱柱垂直a轴截面的宽度约为818~960 nm, 并与文石层呈近45°斜交。 同时, 卧蚕-1热液喷口深海偏顶蛤壳体棱柱层出现交错现象。 壳体的过渡层形状极不规则, 延续了棱柱层的生长取向, 整体显示从棱柱状到文石板片状过渡的趋势。 文石层具有片状结构, 即砖墙结构; 文石形态较为规则、 平直, 厚度约为205~1 260 nm。 肌棱柱层具有简单的棱柱状超微结构特征。
(3)深海偏顶蛤壳体珍珠层和棱柱层矿物分别为文石和方解石, 其中方解石和文石分别为生物成因和无机成因, 文石的结晶度略高于方解石。 贝壳壳体可能是一种潜在的研究深海生物矿化过程中有机质与无机质复合机制的优良载体。
猜你喜欢
排灌机械工程学报(2022年6期)2022-06-23
航空维修与工程(2022年12期)2022-02-04
宝藏(2021年8期)2021-09-15
——详解淄博文石皴纹及赏石文化
宝藏(2021年1期)2021-03-10
宝藏(2019年9期)2019-09-25
数学物理学报(2019年2期)2019-05-10
宝藏(2018年12期)2019-01-29
新世纪智能(数学备考)(2018年9期)2018-11-08
疯狂英语·新读写(2017年7期)2017-08-08
制造技术与机床(2017年12期)2017-02-02
