
碱性亮氨酸拉链和W2结构域2对子宫内膜癌细胞生物学行为的作用及机制
2023-11-03 07:25徐姗姗江征慧未亚兰江飞云
实用临床医药杂志 2023年18期
唐 彬, 蔡 文, 徐姗姗, 江征慧, 未亚兰, 江飞云
(华东师范大学附属芜湖医院/安徽省芜湖市第二人民医院 妇科, 安徽 芜湖, 241000)
子宫内膜癌是发生于子宫内膜的常见妇科恶性肿瘤,多见于围绝经期及绝经后女性[1-2]。肥胖、糖尿病、高脂血症、高雌激素水平等在内的因素可增加子宫内膜癌的发病风险[3]。早期子宫内膜癌患者的生存率可通过手术、化疗、放疗得到改善,但晚期患者的预后仍较差[2,4]。因此,探究子宫内膜癌的发病机制并鉴别有效的治疗靶点对改善子宫内膜癌预后有重要意义[5]。bZIP转录因子家族成员碱性亮氨酸拉链和W2结构域2(BZW2)可参与调控细胞黏附,其机制可能与介导钙黏蛋白有关。BZW2在多种癌症中主要发挥致癌作用,但BZW2在子宫内膜癌中的表达和作用尚不明确。Wnt/β-连环蛋白(Wnt/β-catenin)信号通路是一条在生物进化中极为保守的通路,参与调控多种生物学功能,如细胞增殖、凋亡、分化等[6]。Wnt/β-catenin通路失调与子宫内膜癌的发生发展有关[7]。基于上述报道,本研究探讨BZW2对子宫内膜癌细胞生物学行为的作用及其机制。
1 材料和方法
1.1 材料
人子宫内膜基质细胞系ESC购自青旗(上海)生物技术发展有限公司; 人子宫内膜癌细胞Ishikawa购自北纳生物; 人子宫内膜癌细胞KLE、RL95-2、HEC-1-A购自美国菌种保藏中心; DMEM培养基、胎牛血清(FBS)购自Biowest公司; McCoy′s 5A培养基购自赛百慷(上海)生物技术股份有限公司; SKL2001购自大连美仑生物技术有限公司; siRNA-BZW2#1/2(siRNA-BZW2#1: 5′-GGCTCAAGATTAGATTATCGTCG-3′; siRNA-BZW2#2: 5′-CTGTGTGTTTTCAGCAAATGAAG-3′)和阴性对照siRNA-NC由武汉金开瑞生物工程有限公司构建; LipofectamineTM3000转染试剂购自美国Invitrogen公司; Steadypure 通用RNA提取试剂、Evo M-MLV反转录试剂盒、SYBR®Green Pro Taq HS预混型 qPCR试剂盒购自艾科瑞生物; RIPA细胞裂解液、BCA蛋白定量试剂盒、极超敏ECL化学发光液、碘化丙啶(PI)和核糖核酸酶(RNase)A均购自北京拜尔迪生物技术有限公司; CCK-8试剂购自北京百瑞极生物科技有限公司; 基质胶、Transwell小室购自美国BD公司; 抗BZW2(货号ab254771)、抗Ki-67(货号ab16667)、抗增殖细胞核抗原(PCNA, 货号ab92552)、抗基质金属蛋白酶2(MMP2, 货号ab92536)、抗基质金属蛋白酶9(MMP9, 货号ab76003)、抗周期依赖性激酶4(CDK4, 货号ab108357)、抗周期依赖性激酶6(CDK6,货号ab124821)、抗β-catenin(货号ab32572)、抗c-Myc(货号ab32072)、抗磷酸化糖原合成酶激酶3β(p-GSK3β, 货号ab75814)、抗GSK3β(货号ab32391)、抗GAPDH(货号ab9485)、抗Lamin B1(货号ab229025)、辣根过氧化物酶(HRP)标记的羊抗兔IgG二抗(货号ab6721)均购自英国Abcam公司。
1.2 方法
1.2.1 细胞培养、处理和质粒转染: 含10% FBS的DMEM培养基用于ESC、Ishikawa、KLE、RL95-2细胞的培养; 含10% FBS的McCoy′s 5A培养基用于HEC-1-A细胞的培养。孵育环境为37 ℃、5% CO2。将HEC-1-A细胞接种于6孔板中(5×104个/孔),严格按照LipofectamineTM3000说明书将siRNA-BZW2#1/2、siRNA-NC瞬时转染至汇合度为60%~70%的细胞中,转染持续48 h, 分为4组,分别为Control组、siRNA-NC组、siRNA-BZW2#1组、siRNA-BZW2#2组。为验证BZW2通过Wnt信号进而参与调控子宫内膜癌进展,加入40 μmol/L SKL2001对HEC-1-A细胞进行处理,分为4组, 分别为Control组、siRNA-NC组、siRNA-BZW2组、siRNA-BZW2+SKL2001组。
1.2.2 实时荧光定量聚合酶链反应(RT-qPCR): 收集人子宫内膜癌细胞(Ishikawa、KLE、RL95-2、HEC-1-A)及人子宫内膜基质细胞系ESC, 利用Steadypure通用RNA提取试剂和Evo M-MLV反转录试剂盒分别提取总RNA并进行cDNA合成。在PCR反应体系中按照说明书加入SYBR®Green Pro Taq HS 预混型qPCR试剂盒。采用2-△△Ct法进行分析并相对于GAPDH进行标准量化。
1.2.3 蛋白质印迹法(Western blot): 人子宫内膜癌细胞及人子宫内膜基质细胞系ESC或“1.2.1”分组HEC-1-A细胞加入100 μL预冷的RIPA裂解液进行充分裂解,并采用二喹啉甲酸(BCA)试剂盒检测蛋白浓度。通过10%十二烷基硫酸钠聚丙烯酰胺凝胶电泳(SDS-PAGE)分离后将蛋白转至聚偏二氟乙烯(PVDF)膜, 5%脱脂牛奶室温封闭2 h后,与一抗在4 ℃条件下孵育过夜。洗膜缓冲液(TBST)洗涤后,加入HPR标记的羊抗兔二抗,室温孵育1 h。加入ECL发光试剂盒对蛋白条带进行显影,并拍照观察。
1.2.4 CCK-8检测细胞活力: 在96孔板中配备转染后的HEC-1-A细胞,每孔约4×103个细胞。利用450 nm的酶标仪分析光密度(OD)值之前,滴入10 μL CCK-8进行2 h孵育。
1.2.5 克隆形成实验检测细胞增殖能力: 在6孔板中配备转染后的HEC-1-A细胞,每孔约200个细胞。2周后利用磷酸盐缓冲液(PBS)洗涤细胞,多聚甲醛固定、结晶紫染色,利用光学显微镜进行克隆数统计。
1.2.6 划痕实验检测细胞迁移: 在6孔板中配备转染后的HEC-1-A细胞,使用移液器在汇合度为90%的细胞中形成划痕,利用无血清培养基继续孵育经PBS冲洗后的细胞。分别于0、24 h时取样拍照,计算细胞迁移率。
1.2.7 Transwell实验检测细胞侵袭: 在铺有基质胶的Transwell上室和下室内分别配备转染后的HEC-1-A细胞和500 μL含10% FBS的McCoy′s 5A培养基。24 h后,拭去残留细胞,细胞经固定和染色后,于光学显微镜下观察。
1.2.8 流式细胞术检测细胞周期: 收集转染后的HEC-1-A细胞,加入预冷体积分数70%的乙醇进行固定,并将细胞重悬于PI/RNase A染色缓冲液中,黑暗中孵育30 min。流式细胞仪上机检测。
1.2.9 数据库分析: 通过GSEA分析BZW2在Wnt信号通路中的富集度。
1.3 统计学分析

2 结 果
2.1 BZW2在子宫内膜癌细胞中表达呈上升趋势
通过RT-qPCR和Western blot分析发现, BZW2在人子宫内膜癌细胞(Ishikawa、KLE、RL95-2、HEC-1-A)中的表达较人子宫内膜基质细胞系ESC增加,差异有统计学意义(P<0.001)(图1A、图1B)。后续选择BZW2相对表达量最高的HEC-1-A细胞进行实验。
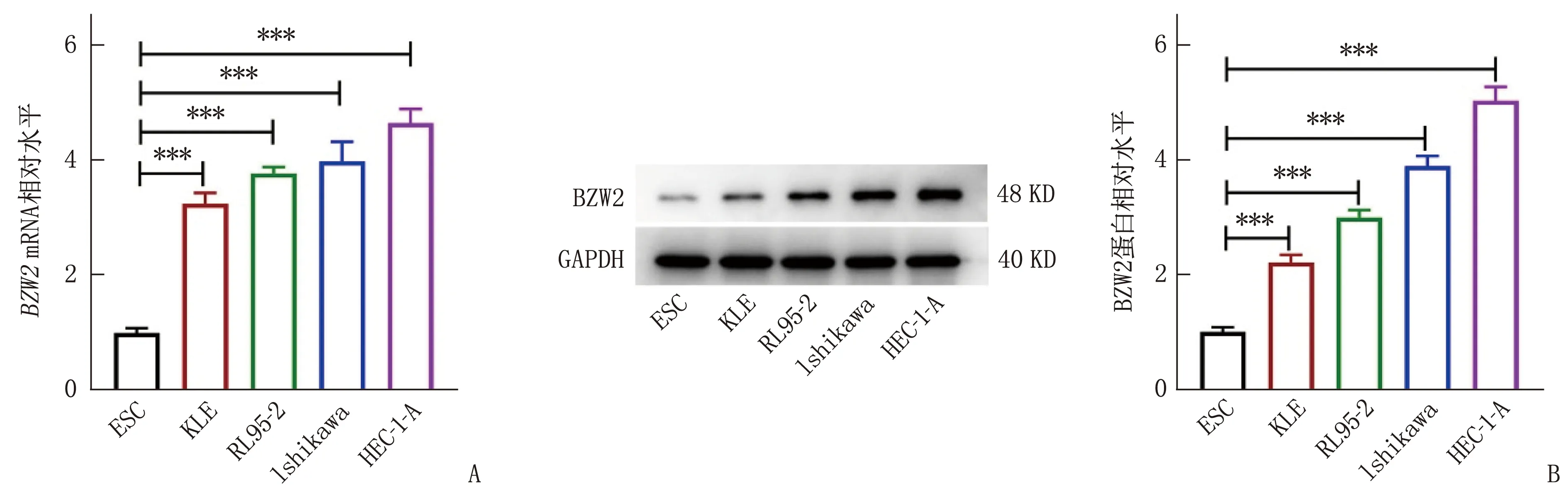
A: RT-qPCR检测子宫内膜癌细胞中BZW2 mRNA表达水平。B: Western blot检测BZW2在子宫内膜癌细胞中的蛋白表达水平。与ESC比较, ∗∗∗P<0.001。图1 BZW2在子宫内膜癌细胞系中表达增加
2.2 BZW2缺失抑制子宫内膜癌细胞增殖
为阐明BZW2对子宫内膜癌发生发展的影响,本研究构建BZW2干扰质粒并将其转染至HEC-1-A细胞,RT-qPCR和Western blot检测结果表明,与siRNA-NC组比较, siRNA-BZW2#1组、siRNA-BZW2#2组的BZW2mRNA和BZW2蛋白表达均下调,差异有统计学意义(P<0.001)(图2A、2B), 故后续实验选择干扰效率较为显著的siRNA-BZW2#2质粒。siRNA-BZW2组较si-NC组细胞活性降低,差异有统计学意义(P<0.001)(图2C)。克隆形成实验结果表明,与si-NC组相比, siRNA-BZW2组细胞克隆形成能力减弱,差异有统计学意义(P<0.001)(图2D)。Western blot检测发现,敲低BZW2导致增殖相关Ki-67和PCNA蛋白表达下降,差异有统计学意义(P<0.001)(图2E)。
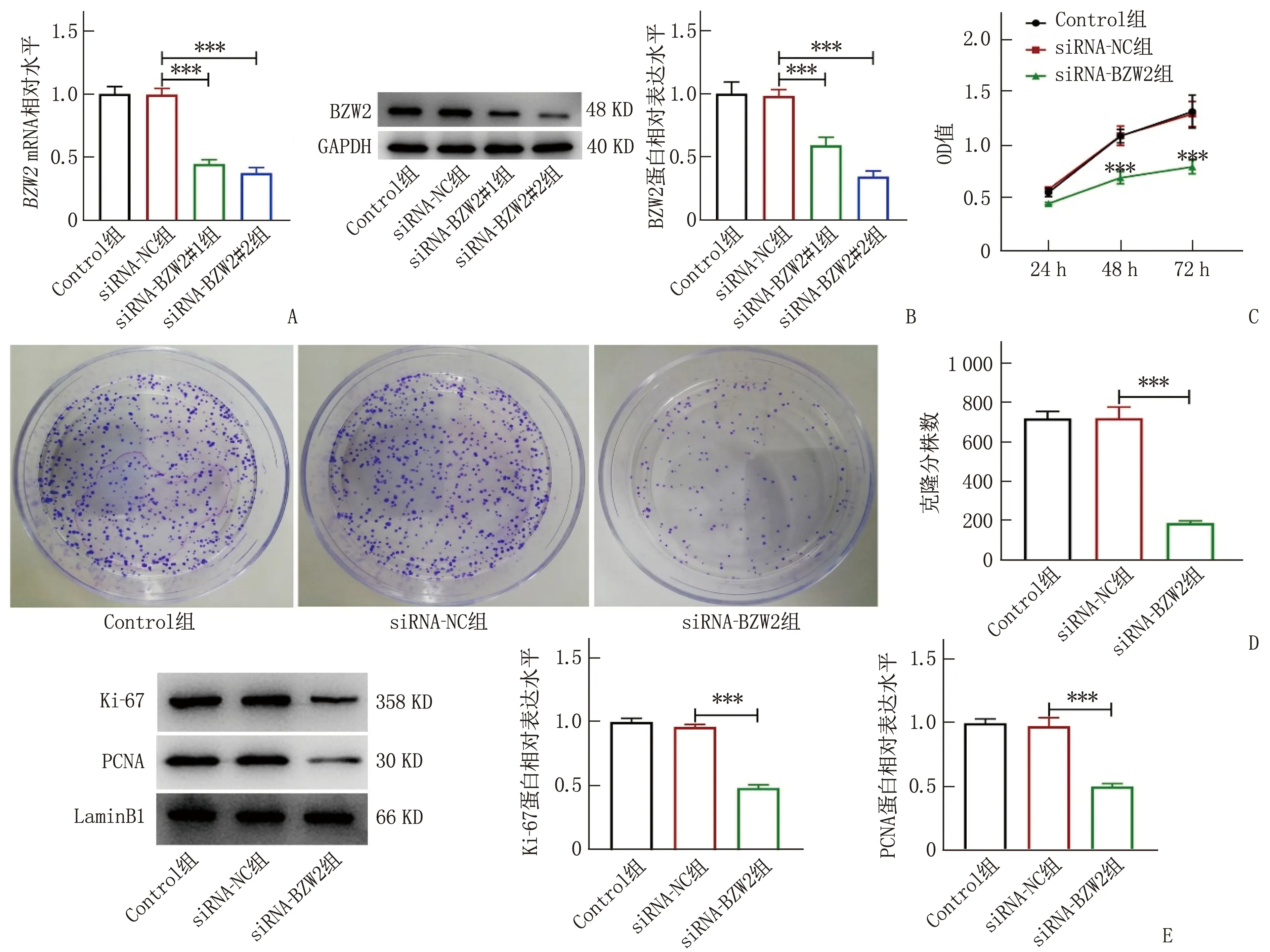
A: RT-qPCR检测质粒转染后BZW2的mRNA表达水平; B: Western blot检测质粒转染后BZW2的蛋白表达水平; C: CCK-8检测HEC-1-A细胞活力; D: 克隆形成实验检测HEC-1-A细胞克隆形成能力; E: Western blot检测增殖相关蛋白表达水平。与siRNA-NC组比较, ∗∗∗P<0.001。图2 BZW2缺失抑制子宫内膜癌细胞增殖
2.3 BZW2缺失抑制子宫内膜癌细胞侵袭和迁移
划痕实验显示,与si-NC组相比, siRNA-BZW2组迁移细胞数目大幅度下降,差异有统计学意义(P<0.001)(图3A)。此外, siRNA-BZW2组较si-NC组侵袭细胞数目降低,差异有统计学意义(P<0.001)(图3B)。Western blot分析发现,干扰BZW2可抑制转移相关蛋白MMP2和MMP9表达,差异有统计学意义(P<0.001)(图3C)。
2.4 BZW2缺失诱导G0/G1期细胞阻滞
如图4A所示,流式细胞分析结果表明, siRNA-BZW2组较si-NC组G1期细胞数量增加, S期和G2期细胞数量减少,差异有统计学意义(P<0.001)(图4A)。Western blot分析周期相关蛋白CDK4和CDK6表达,结果发现, siRNA-BZW2组较si-NC组CDK4和CDK6蛋白表达下调,差异有统计学意义(P<0.001)(图4B)。

A: 流式细胞术检测HEC-1-A细胞周期; B: Western blot检测周期相关蛋白表达水平。与siRNA-NC组比较, ∗∗∗P<0.001。图4 BZW2 缺失诱导G0/G1期细胞阻滞
2.5 BZW2缺失抑制Wnt/β-catenin通路
经GSEA分析发现, BZW2显著富集于Wnt信号通路中(图5A)。进一步Western blot分析结果揭示,与si-NC组比较, siRNA-BZW2组β-catenin、c-Myc、p-GSK3β蛋白表达减少,差异有统计学意义(P<0.001); 而添加Wnt/β-catenin信号通路激动剂SKL2001后,上述蛋白表达再次增加,差异有统计学意义(P<0.001)(图5B)。

A: GSEA分析检测BZW2在Wnt信号通路中的富集度; B: Western blot检测Wnt信号通路相关蛋白表达水平。与siRNA-NC组比较, ∗∗∗P<0.001; 与siRNA-BZW2组比较, ###P<0.001。图5 BZW2缺失抑制Wnt/β-catenin通路
2.6 SKL2001部分逆转了BZW2干扰对子宫内膜癌细胞增殖、迁移、侵袭的作用
为验证BZW2是否通过调控Wnt/β-catenin通路进而参与子宫内膜癌发生发展,本研究利用SKL2001对细胞进行处理,并分为Control组、siRNA-BZW2组及siRNA-BZW2+SKL2001组。CCK-8结果表明,与Control组相比, BZW2缺失导致细胞活性下降,差异有统计学意义(P<0.001); 与siRNA-BZW2组相比, siRNA-BZW2+SKL2001组中细胞活性有所提升,差异有统计学意义(P<0.001)(图6A)。克隆形成实验证明, siRNA-BZW2组较Control组克隆形成数减少,差异有统计学意义(P<0.001); siRNA-BZW2+SKL2001组较siRNA-BZW2组克隆形成数增加,差异有统计学意义(P<0.001)(图6B)。Western blot分析同样发现,与Control组比较, siRNA-BZW2组Ki-67和PCNA蛋白表达减少,差异有统计学意义(P<0.001); 与siRNA-BZW2组相比, siRNA-BZW2+SKL2001组Ki-67和PCNA蛋白表达再次上升,差异有统计学意义(P<0.05或P<0.001)(图6C)。此外,与Control组相较, siRNA-BZW2组迁移和侵袭细胞数减少,差异有统计学意义(P<0.001); siRNA-BZW2+SKL2001组较siRNA-BZW2组迁移和侵袭细胞数增加,差异有统计学意义(P<0.001)(图6D、6E)。Western blot分析结果同样表明, siRNA-BZW2组较Control组MMP2、MMP9蛋白表达下调,差异有统计学意义(P<0.001); 而siRNA-BZW2+SKL2001组较siRNA-BZW2组MMP2、MMP9蛋白表达增加,差异有统计学意义(P<0.001)(图6F)。
2.7 SKL2001部分逆转了BZW2干扰对子宫内膜癌细胞周期的作用
流式细胞术和Western blot分析结果表明,与si-NC组比较,siRNA-BZW2组G0/G1期细胞占比高, S期和G2期细胞占比低,且CDK4和CDK6蛋白表达降低,差异有统计学意义(P<0.001)。与siRNA-BZW2组比较, siRNA-BZW2+SKL2001组G1期细胞占比下降, S期细胞占比升高,且CDK4和CDK6蛋白表达上调,差异有统计学意义(P<0.01或P<0.001)(图7A、7B)。
3 讨 论
子宫内膜癌是引起女性死亡的重要原因之一[8]。虽然已有报道[9]揭示基因表达异常与子宫内膜癌的恶性生物学特征的相关性,但肿瘤侵袭转移相关机制仍亟待阐明。因此,探究潜在的新型肿瘤生物标志物对子宫内膜癌诊断和靶向治疗具有重要意义。
BZW2可在恶性肿瘤中作为致癌因子驱动特定的细胞生物活动。本研究同样表明,BZW2在子宫内膜癌细胞中表达上调,且敲低BZW2表达可导致增殖、迁移和侵袭细胞数目下降。增殖相关蛋白Ki-67和PCNA表达增加与子宫内膜癌不良预后相关[10]。此外,转移相关蛋白MMP2和MMP9[11]在子宫内膜癌组织中异常表达并与子宫内膜异位症的转移和侵袭有关[12]。同样,干扰BZW2可导致Ki-67、PCNA、MMP2和MMP9蛋白表达显著下降。以上实验结果均表明敲低BZW2可通过抑制子宫内膜癌细胞增殖、迁移和侵袭,从而发挥抑癌作用。
细胞周期的快速运转是肿瘤细胞增殖的重要机制[13]。细胞周期包括G1/S期、G2/M期重要检查点,任意阶段的细胞周期调控异常都会导致细胞周期停滞,并抑制细胞增殖[14]。研究[15]发现,BZW2缺失可导致肌层浸润性膀胱癌G1期细胞停滞; 研究[16]表明,敲低BZW2表达可促进骨肉瘤G2/M期细胞停滞。本研究结果揭示,抑制BZW2可导致G0/G1期细胞比例增加, S期和G2期细胞比例减少。CDK4和CDK6是细胞周期的核心调控因子,且CDK4/6抑制剂可抑制子宫内膜癌肿瘤生成[17-18]。本研究中,功能丧失实验结果表明,敲低BZW2后CDK4和CDK6蛋白表达呈现显著下降趋势。
GSEA分析数据表明, BZW2显著富集于Wnt信号通路中; 且沉默BZW2可显著减少子宫内膜癌细胞中Wnt/β-catenin通路相关蛋白β-catenin、c-Myc、p-GSK3β/GSK3β表达; 而Wnt/β-catenin信号通路激动剂SKL2001可再次促进以上蛋白表达。本研究结果显示,SKL2001可部分恢复BZW2缺失对子宫内膜癌细胞增殖、迁移、侵袭及细胞周期的影响。
综上所述, BZW2缺失可阻碍子宫内膜癌细胞增殖、迁移、侵袭,并诱导细胞周期阻滞,其机制可能与Wnt/β-catenin信号通路失活有关,表明BZW2可作为诊断子宫内膜癌的分子标志物及治疗子宫内膜癌的潜在分子靶标。
猜你喜欢
服饰导报·鞋世界(2021年4期)2021-05-17
奥秘(创新大赛)(2019年9期)2019-10-09
小哥白尼(趣味科学)(2019年1期)2019-04-12
中国药理学通报(2019年5期)2019-01-11
奥秘(2017年5期)2017-07-05
环境与生活(2016年6期)2016-02-27
山东医药(2015年14期)2016-01-12
当代畜禽养殖业(2014年10期)2014-02-27
生殖医学杂志(2013年4期)2013-03-11
河北医科大学学报(2011年9期)2011-03-25
