
一株白木香不同组织内生菌的分离鉴定与多样性分析
2023-11-01 02:24唐冰沣黄文政陈卫国刘学林陈欣强符韵林
安徽农业科学 2023年20期
邓 力,唐冰沣,黄文政,陈卫国,刘学林,陈欣强,符韵林,刘 鑫*
(1.广西国有钦廉林场,广西钦州 535000;2.广西大学生命科学与技术学院,广西南宁 530000;3.广西大学林学院,广西南宁 530000)
白木香[Aquilariasinensis(Lour.) Gilg.]又称土沉香,主要分布在我国南部地区,如海南、广西、广东、云南、香港、台湾等[1]。白木香所产生的树脂是我国和东南亚部分国家的天然香料和传统名贵药物,具有降气温中、暖身纳气的效果[2],同时能够用于熏香、香水、药物、心脏补品等[3];从沉香中提取的精油也具有抗菌特性[4]。沉香的高价值引发的商业开发和不受控制的砍伐正在导致这些树木数量的减少,目前8种沉香属的植物被列于《濒危物种国际贸易公约》附录Ⅱ(1994),并被国际自然保护联盟(IUCN,2009)列为易受伤害物种。
沉香为沉香属植物中的树脂,是沉香属植物在受到外来刺激或者损伤之后所产生的次生代谢物[5],其成分以倍半萜类化合物(如沉香螺旋醇、白木香酸、白木香醛)和色酮类化合物[2-(2-苯乙基)色酮衍生物]为主。国产沉香化学成分的研究表明白木香酸和白木香醛的产生是为了应对某些外来因素刺激的防御[6],许多学者针对沉香结香原理进行了研究,Wang等[7]揭示了沉香中2-(2-苯基乙基)色酮的生物合成途径。沉香作为一种名贵的药材,其自然发育需要25~30年,产量较低,无法满足市场需求[5]。该研究对广西境内一株老龄野生白木香的内生菌进行分离鉴定,探究其内生菌结构,了解其内生菌的菌群多样性,分析白木香不同部位的内生菌群结构差异,为功能菌株的发掘、人工诱导结香及研究内生菌群与宿主的相互作用关系奠定基础。
1 材料与方法
1.1 材料采集和处理于2021年9月在广西钦州浦北县某村进行白木香[Aquilariasinensis(Lour.) Gilg.]样本采集。该树树龄约130年、树高15 m、胸径64.5 cm。在不伤害白木香的前提下,采集根部、叶片、树干表皮有明显病斑部位、表皮以下1~2 cm内层植物组织进行表面消毒后,放入灭菌袋中,编号,保存于4 ℃,并在24 h内对样品进行处理。
1.2 菌株分离纯化白木香内生真菌的分离与纯化分别使用马铃薯葡萄糖琼脂培养基和沙氏琼脂培养基;细菌分别使用Luria-Bertani琼脂培养基和牛肉膏蛋白胨琼脂培养基。
样品分离前,按下列程序进行表面消毒: 分别使用30%过氧化氢和75%乙醇进行表面消毒,消毒结束后使用无菌水漂洗2~3次,漂洗后使用无菌滤纸吸干表面水分。
分离纯化方法:①插块培养法。将消毒后的样品使用无菌手术刀切去表面后,裁成约5 mm的均匀小块,分别插入相应的培养基中,使截面与培养基充分接触,真菌培养基分别添加50 mg/L氨苄西林和50 mg/L卡那霉素,置于(28±1)℃恒温培养箱培养2~10 d。②液培涂布法。将消毒后的样品使用无菌手术刀切去表面后,裁成约5 mm的均匀小块后,用无菌研钵稍加研磨,将样品内部暴露后分别接入相应的液体培养基中,(28±1)℃摇床培养1~3 d。待培养液浑浊后将培养液稀释至10-4和10-5在对应的培养基中进行涂布,涂布后置于(28±1)℃恒温培养箱培养2~10 d。③纯化。待插块样品的培养基和进行液培稀释涂布的培养基上长出菌落后,通过对不同菌落形态特征进行区分,使用无菌接种环分别挑取不同群落接入对应固体培养基中,在(28±1)℃恒温培养箱培养3~5 d,进行纯化分离。
1.3 内生菌的鉴定
1.3.1真菌的分子生物学鉴定。将培养基上无法产孢的内生真菌培养一段时间后,从平板上刮取约 1 g 菌丝体,置于1.5 mL 离心管中,使用上海生工DNA提取试剂盒进行内生真菌DNA的提取,并以此为模板,采用真菌通用引物ITS-4F (TCCTCCGCTTATTGATATGC)和ITS-5R(GCAAGTAAAAGTCGTAACAAGG)扩增菌株的ITS序列。PCR 反应体系(50 μL体系):TaqDNA Polymerase 0.5 μL,ITS-4F和ITS-5R各 4.0 μL,模板 DNA 9.0 μL,双蒸水补足 50 μL。扩增程序:95 ℃ 5 min;94 ℃ 30 s, 50 ℃ 1 min,72 ℃ 1 min,35个循环;72 ℃ 10 min。 扩增产物用 1.2%琼脂糖凝胶核酸染料电泳检测,将符合要求的扩增产物送至上海生工生物技术有限公司进行测序。
1.3.2细菌的分子生物学鉴定。将在培养基上的细菌菌落挑至液体培养基中,37 ℃、200 r/min摇床培养12 h,用枪头吸取10 μL菌液于0.2 mL PCR管中,采用细菌16S通用引物27F(AGAGTTTGATCMTGGCTCAG)和1492R(GGTTACCTTGTTACGACTT )扩增菌株16S序列。PCR反应体系(25 μL体系):2× RapidTaqPCR Master Mix 12.5 μL,16S通用引物27F和1492R各1.0 μL,双蒸水补足25 μL。扩增程序:95 ℃ 5 min;95 ℃ 30 s, 54 ℃ 30 s,72 ℃ 30 s,35个循环;72 ℃ 5 min。扩增产物用1.5%琼脂糖凝胶核酸染料电泳检测,将符合要求的扩增产物送至上海生工生物技术有限公司进行测序。
1.3.3内生菌的分子生物学分析。测得的序列在NCBI(https://www.ncbi.nlm.nih.gov/)上使用Blast工具进行检索比对,选择相似度大于98%以上的序列作为参考序列,并采用ClustalX 2.0 进行同源性比较和匹配排序。
1.4 内生菌多样性分析该研究采用分离频率、多样性指数等方面分析白木香内生菌的多样性以及分布特性[8]。
2 结果与分析
2.1 白木香内生菌的组成
2.1.1白木香内生细菌的组成。从所取样品中分离得到内生细菌111株,经形态学和16S rRNA分析,得到的菌株分属于13个属。由表1可知,在门分类水平上分离得到的菌株多为厚壁菌门和变形菌门,在目的分类水平上,芽孢杆菌目和肠杆菌目分离频率较高,分别为45.94%和39.64%;在科的分类水平上,芽孢杆菌科和肠杆菌科分离频率较高,分别为45.94%和33.33%;在属的水平上以芽孢杆菌属(Bacillus)、肠杆菌属(Enterobacter)和假单孢菌属(Pseudomonas)分离频率较高,分别为36.03%、10.83%和9.92%。部分细菌菌落形态如图1所示。
图1 部分细菌菌落形态Fig.1 Morphology of partial bacterial colonies
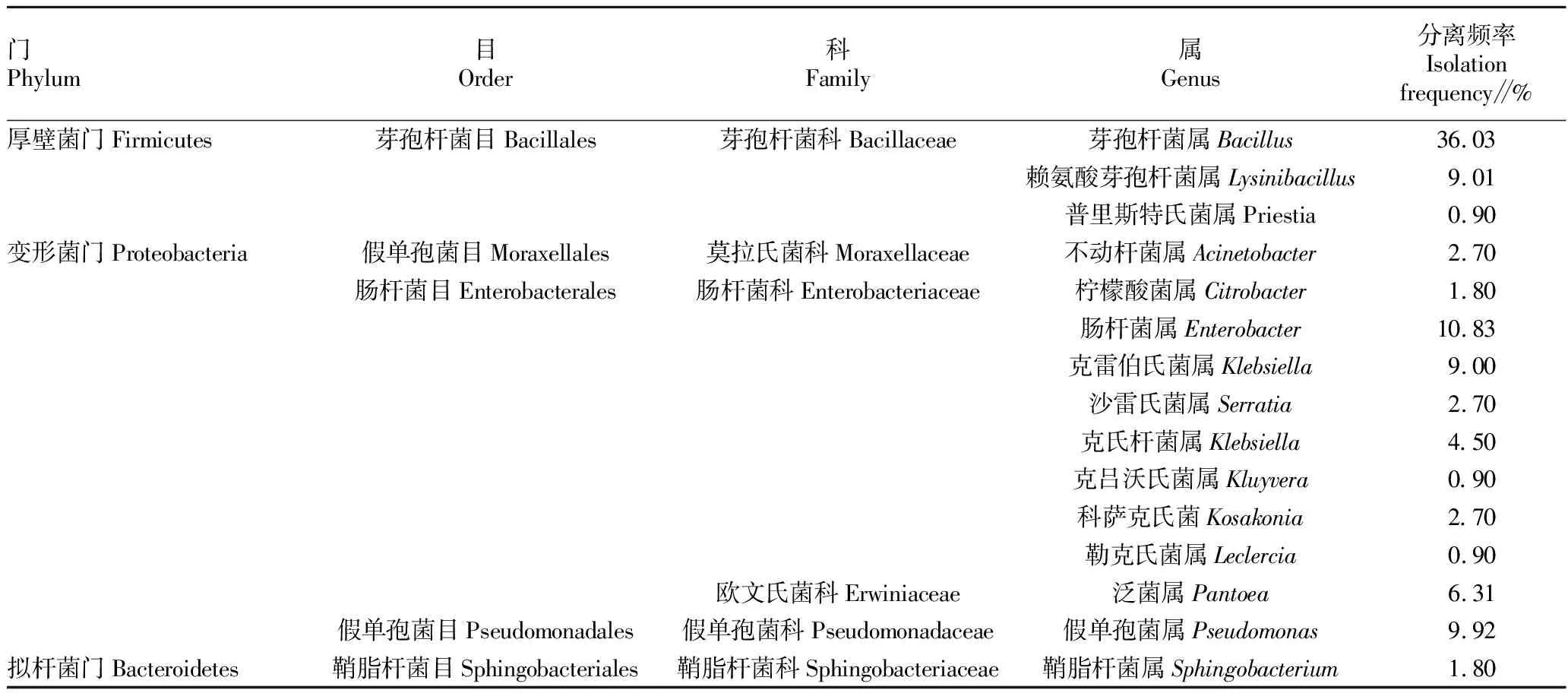
表1 白木香内生细菌的菌群组成
2.1.2白木香内生真菌的组成。从所取样品中分离得到内生真菌49株,经形态学和ITS序列分析,得到的菌株分属于10个属。由表2可知,分离菌株均为子囊菌门,在目的分类水平上,肉座菌目、葡萄座腔菌目分离频率较高,分别为36.74%、22.45%;在科的分类水平上,赤壳菌科、葡萄座腔菌科和小丛壳科分离频率较高,分别为24.49%、22.45%和20.41%;在属的水平上,镰刀菌属、可可毛色二孢菌属和炭疽菌属分离频率较高,分别为24.49%、22.45%和20.41%。部分真菌菌落形态如图2所示。

表2 白木香内生真菌的菌群组成
2.2 白木香内生菌多样性分析
2.2.1不同部位内生细菌多样性分析。从表3可以看出,白木香不同部位的细菌分离频率差别较大,其中从干茎中分离得到的菌株最多,占分离菌株的86.48%,其次为干茎内层(5.41%)、根部(5.41%),从叶中得到的菌株最少(2.70%)。不同部位的细菌分布差异较大,其中,干茎中分离得到96株,在属的分类水平上有13个属,其中占比较高的有芽孢杆菌属和肠杆菌属;干茎内层中分离得到6株菌株,在属的分类水平上分别为芽孢杆菌属、赖氨酸芽孢杆菌属、肠杆菌属、假单孢菌属;根部分离得到6株菌株,在属的分类水平上分别为芽孢杆菌属、柠檬酸菌属、肠杆菌属和假单孢菌属;叶片分离得到3株菌株,在属的分类水平上有2个属,分别为芽孢杆菌属和泛菌属。
2.2.2不同部位内生真菌多样性分析。从表4可以看出,不同部位的真菌分离频率差别较大,其中从干茎中分离得到的菌株最多,占分离总菌株数的71.43%,其次为根部(12.25%)、叶(10.20%)、干茎内层(6.12%)。不同部位的真菌分布差异较大,其中,干茎分离得到35株,在属的分类水平上有9个属,其中,主导菌株为炭疽菌属、镰刀菌属和可可毛色二孢菌属;从叶片分离得到5株,属分类水平上分为4个属,分别为木霉属、可可毛色二孢菌属、镰刀菌属和炭疽菌属;从根部分离得到6株,属分类水平上有5个属,分别为炭疽菌属、附球霉属、镰刀菌属、可可毛色二孢菌属、木霉属;从干茎内层分离得到3株,在分类水平上分属3个属,分别为附球霉属、镰刀菌属、平脐蠕孢属。

表4 白木香不同部位内生真菌类群组成、分布、多样性指数和分离频率
2.3 白木香内生菌相似性
2.3.1内生细菌相似性分析。各部位的细菌菌种相似性如图3所示。各部分分离得到的菌株在属水平分类差别较大;根部分离出的内生细菌与其他部分有较大重合度;干茎中分分离得到最多菌株,但所分离得到的菌株在属水平分类上与其他部位分离出的菌株重合度不高。

图3 不同部位细菌属水平上的分布韦恩图Fig.3 Venn diagram of the distribution of bacteria genera in different parts
2.3.2内生真菌相似性分析。各部位的真菌菌种相似性如图4所示。与分离得到的细菌在属水平分类上较为相似,各部分离得到的菌株在属水平分类差别较大;根部分离出的内生真菌在属水平分类上与其他部分有较大重合度;干茎中分分离得到最多菌株,但所分离得到的菌株在属水平分类上与其他部位分离出的菌株重合度不高。
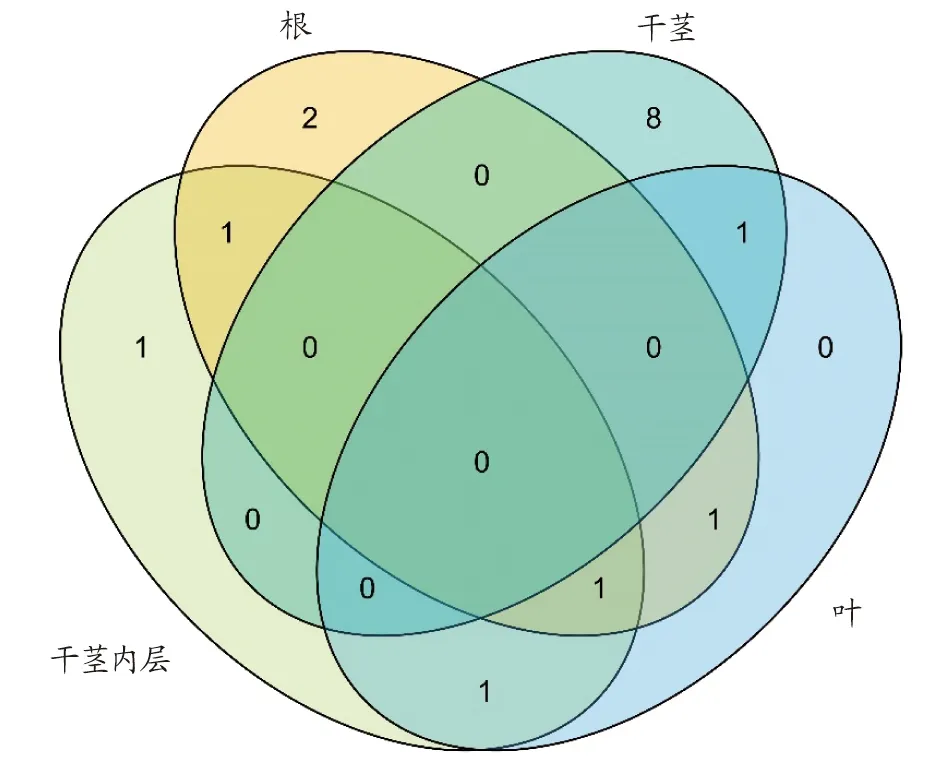
图4 不同部位真菌属水平上的分布韦恩图Fig.4 Venn diagram of the distribution of fungi genera in different parts
3 小结与讨论
从20世纪以来,国内外研究者从沉香属植物树体分离了一些内生真菌,主要为镰刀菌、毛色二孢菌、可可毛色二孢菌、曲霉、木霉、毛霉、青霉等。研究认为沉香的形成可能是由于树体被一种或者多种真菌侵染寄生,在真菌体内酶的作用下,导致木薄壁细胞贮藏的淀粉发生变化,形成香脂,多年沉积形成沉香。戚树源等[9]针对黄绿墨耳菌与沉香结香机制进行了研究,发现沉香组织中色酮类成分含量与真菌侵染时间呈正相关,且沉香主要成分的形成很可能是在逆境条件下白木香和内生真菌相互作用所产生的代谢产物。Bhore等[10]对沉香中可培养的77种细菌进行分析,发现大部分(36.4%)为短小芽孢杆菌(Bacilluspumilus)。近年来大量的研究表明镰刀菌、毛色二孢菌、可可毛色二孢菌、曲霉、木霉、毛霉、青霉等真菌均具有促进沉香树结香的效果[6],也有研究证明,内生细菌对沉香的产生也有促进作用,Chhipa 等[11]利用泛菌(Pantoea)成功诱导了沉香螺醇的产生;该研究从白木香的叶、茎、根和周边干茎内层等不同部位分离出111株细菌和49株真菌。其中细菌中厚壁菌门和变形菌门的占比较大,主要为芽孢杆菌属、肠杆菌属和假单孢菌属。真菌均属子囊菌门,主要为镰刀菌、炭疽菌和可可毛色二孢菌。陈瑶[12]从齐楠沉香[Aquilariasinensis(Lour.) Gilg.]分离得到的真菌有担子菌门和子囊菌门,在属水平上的优势菌为镰刀菌属、枝孢属和帚枝霉属,此结果与该试验有较大区别,该试验分离得到的真菌在门分类水平上专一性高,可能与试验树种有关系,该试验样品采自多年树龄的结香白木香。
分离得到的根部中的细菌有芽孢杆菌属、柠檬酸菌属、肠杆菌属和假单孢菌属,分离得到的干茎内层的细菌有芽孢杆菌属、赖氨酸芽孢杆菌属、肠杆菌属和假单孢菌属。其中肠杆菌属和芽孢杆菌属是常见的植物根际促生细菌;分离得到的干茎内层真菌主要有镰刀菌属、附球霉属和平脐蠕孢属,存在于根部的真菌主要有炭疽菌属、附球霉属、镰刀菌属、可可毛色二孢菌属和木霉属,二者也有高度的相似性。存在于干茎内层和根部中的真菌多为植物致病菌,或与真菌从树干入侵后,菌丝在树干内分布,或者直接从土壤入侵,进而渗透到根部有关[13]。
从叶片中分离得到的细菌只有芽孢杆菌属和泛菌属,分离得到的真菌有炭疽菌属、可可毛色二孢菌属、镰刀菌属和木霉属,均为常见的植物致病菌。可分离的真菌在叶片的分布与其在根部的分布高度相似,由于叶片和根茎不是结香部位,并且上述细菌和真菌常见于各种植物,所以其部位上分离得到的内生菌应该与结香没有直接关系。
干茎上有丰富的结香,于干茎部位分离得到大量的真菌和细菌。分离的细菌有芽孢杆菌属、赖氨酸芽孢杆菌属、不动杆菌属、柠檬酸菌属、肠杆菌属、克氏杆菌属、克吕沃氏杆菌属、科萨克氏菌属、泛菌属、假单孢菌属、鞘脂杆菌属、普里斯特氏菌属和勒克氏菌属。分离得到的真菌有节菱孢属、炭疽菌属、镰刀菌属、可可毛色二孢菌属、脉孢菌属、拟盘多毛孢属、青霉属、木霉属、平脐蠕孢属。干茎的细菌和真菌的群落组成与叶片和根部的差异较大。Zheng等[14]研究发现内生真菌在茎中的定殖率显著高于叶片,叶片和根的组织内的真菌群落差异显著。这与该试验的结果一致。
分离得到的真菌主要为镰刀菌属、炭疽菌属和可可毛色二孢菌属,均为常见的植物致病菌。目前已有研究证明镰刀菌属对沉香的产生有重要影响,Faizal等[15]在对Gyrinopsversteegii(Gilg.)接种茄病镰孢菌90 d后对比未接种菌株的穿孔对照组,发现接种镰刀菌的树枝产生了明显的产香区域;GC-MS分析表明接种区域含有别香橙烯、β-真烯醇、β-斯利烯以及色酮衍生物2-(2-苯乙基)色酮-4-1,6-甲氧基-2-(2-苯乙基)铬酮、6,7-二甲氧基-2-(2-苯乙基)铬酮等沉香的特征倍半萜物质 ;继而又使用茉莉酸甲酯和茄病镰孢菌培养液提取物进行AquilariamalaccensisLamk组织培养,发现茄病镰孢菌能够导致沉香化合物生成[16]。毛色二孢菌属也是常见的植物致病菌,广泛分布于热带的亚热带地区。毛色二孢菌会促进产生茉莉酸甲酯,该物质与植物防御有密切关联,可以从化学分子的角度上开启植物防御机制,进而促进结香[17]。Chen等[18]在利用菌株发酵液诱导沉香形成的研究发现可可毛色二孢菌(L.theobromae)和茄病镰孢菌(Fusariumsolani)能够有效导致白木香结香,但茄病镰孢菌无法长期保持结香效果,而可可毛色二孢菌能在一年内持续结香。Zhang等[19]研究发现由可可毛色二孢菌诱导产生沉香后提取的精油与野生沉香的精油无论是在化学成分还是抗菌活性方面都具有高度相似性,进一步证实可可毛色二孢菌促进沉香结香。分离得到的细菌中,占比最大为的芽孢杆菌属,有研究证实,腐生芽孢杆菌通过分解纤维素促进沉香的有效成分释放[20];此外,芽孢杆菌属可以通过产生几丁质酶和挥发物质表现出对镰刀菌属等植物病原菌的抑制作用[21]。
该研究通过对具有百年树龄的结香白木香进行内生菌的分离鉴定,得到了大量的沉香内生菌,丰富了沉香内生菌的多样性,并为人工结香提供了菌种资源。下一步拟使用分离得到的菌株对白木香进行结香试验,进一步探究各类微生物的结香功能和人工结香条件。
猜你喜欢
小小说月刊(2022年14期)2022-07-18
趣味(作文与阅读)(2021年3期)2021-07-19
东北农业大学学报(2020年3期)2020-05-14
生物技术通报(2019年2期)2019-03-15
四川农业大学学报(2018年5期)2018-11-09
中成药(2018年1期)2018-02-02
中国现代中药(2017年8期)2017-09-21
现代装饰(2017年11期)2017-05-25
金色少年(奇趣科普)(2017年1期)2017-03-03
中南民族大学学报(自然科学版)(2015年2期)2015-12-16
