
犀金龟亚科昆虫(鞘翅目∶金龟科)的分类研究历史及中国研究进展
2023-10-20 12:07:24路园园佟一杰王新谱
环境昆虫学报 2023年4期
王 涛 ,路园园,李 升,佟一杰,王新谱*,白 明,3,4*
(1. 宁夏大学农学院,银川 750021;2. 中国科学院动物研究所,动物进化与系统学院重点实验室,北京 100101;3. 中国科学院大学,北京 100049;4. 海南省崖州湾种子实验室,海南三亚 572000)
1 研究简介
犀金龟亚科Dynastinae隶属于昆虫纲Insecta鞘翅目Coleoptera多食亚目Polyphaga金龟总科Scarabaeoidea金龟科Scarabaeidae(Bouchardetal., 2011)。因其头部和前胸背板多数具突出的角突、其它突起或凹坑,酷似“犀牛角”,因此得名犀金龟,英文名Rhinoceros beetle。犀金龟成虫体粗短,背表面近圆形且明显拱起,雌雄二型明显;多数种类体型巨大,体长30~65 mm,上颚多少外露于头部背面可见;触角8~10节不等,10节最常见,末三节鳃叶状;头和前胸背板具角状突起,雄虫尤为显著;中胸小盾片可见,中足基节靠近,后足胫节具2枚端距;鞘翅表面光滑、网状或具刻点;雄虫腹部末端略内凹,雌虫常圆弧形。幼虫蛴螬型;下口式;触角约与头等长;肛门呈“一”或“V”字形(章有为, 1997; 路园园, 2020)。
犀金龟成虫以木本植物为食,幼虫以腐木形成的腐殖质为食,部分幼虫以作物、林木根部为食(张芝利, 1984)。根据活动规律,分为夜出和日夜均活动2种类型。夜间活动的占多数,如双叉犀金龟Trypoxylusdichotoma;日夜均活动的较少,如阔胸禾犀金龟Pentodonmongolicus,该种白天仅见少数个体爬行,且不飞翔(路园园, 2020)。
犀金龟分布广泛,多集中于热带和新热带地区,尤以南美洲种类最为丰富,古北区较为贫乏。犀金龟因其独特外形,吸引众多学者对其进行了研究。截止2021年,世界犀金龟亚科已知8族225属1 860余种(Endrödi, 1985; Ratcliffeetal., 2021),其中扁犀金龟族Pentodontini种类最多,有100属550余种,六齿犀金龟族Hexodontini种类最少,仅有3属10余种(Endrödi, 1985; Ratcliffe and Cave, 2015; Krell and Theuerkauf, 2015)。我国地域辽阔,横跨古北区和东洋区,在世界动物地理区划中占据重要位置。但我国犀金龟亚科类群研究时间较短且相对滞后,仅限于系统分类、生物学习性等相关研究,我国已知的犀金龟亚科种类较少,包括5族17属67种(亚种),仅占世界已知种类的4%左右,主要分布在云南(29种)、广西(23种)、台湾(15种)等地,而在内陆地区记录较少(Krell and Bezděk, 2016; Schoolmeesters, 2021)。
因此,对世界和中国的犀金龟分类研究历史进行整理,系统地开展中国犀金龟亚科的物种多样性与地理分布研究,厘清该亚科各族、各属之间系统发育关系和地理分布,进而提高该类群的分类研究水平,为探究该类群中益虫的开发利用与害虫的防治提供理论基础。本文简要回顾了世界和我国犀金龟亚科的研究历史以及对经济的影响,给出了中国犀金龟亚科物种列表,总结了目前研究现状及存在的问题,并对未来研究进行展望。
2 世界研究历史及问题分析
2.1 分类系统研究
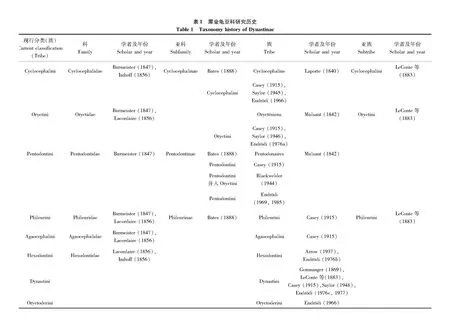
Linnaues(1758)首次在《自然系统》第10版中建立蜣螂属Scarabaeus,该属成为金龟总科雏形,其中犀金龟类群包括8个物种。Lamarck(1807)指定蜣螂属模式种为Scarabaeushercules,该模式种后被Kirby(1825)修订为犀金龟属Dynastes模式种长戟犀金龟Dynasteshercules。MacLeay(1819)提出的20科系统中,犀金龟类群定义为科级阶元Dynastidae,指定模式属为犀金龟属。往后多年,Latreille(1829)、Burmeister(1847)、Lacordaire(1856)、Gemminger(1869)、Bates(1888)等人都沿用这个分类系统。Bates(1888)提出了犀金龟科的6亚科系统,Casey(1915)将犀金龟定义为亚科阶元,Endrödi(1969)等人均采用此分类系统,一直沿用至今。
针对犀金龟亚科下的低级阶元研究,Latreille(1817)首次提出族级分类阶元,然而这时期族级分类系统比较混乱,亚科下的类群分类地位一直变动(表1)。直至1985年,Endrödi通过对全球犀金龟种类研究提出8族分类系统,并利用主要分布地对族的划分提出了建议(表2):六齿犀金龟族只分布在马达加斯加,类蛀犀金龟族Oryctoderini只分布在东洋区、巴布亚区、澳大利亚地区,因这两个族分布地局限,美洲的犀金龟亚科分类没有将其列入其中(Ratcliffe and Cave, 2006)。到目前为止,犀金龟亚科内族级阶元的系统分类仍存在问题,如通过后足胫节顶端边缘截状或圆齿状来区分蛀犀金龟族Oryctini和扁犀金龟族,但其单系尚未得到证实(Gascaetal., 2008)。后续有学者认可该分类特征,但仍存疑,还有待进一步研究(Sanabria-Garcíaetal., 2012; Ratcliffe and Cave, 2015; López-Garcíaetal., 2016)。

表2 世界犀金龟亚科分布情况
尽管Endrödi提出的8族分类系统沿用至今,但族下属级阶元的分类地位经常发生改变。Morón和Ratcliffe(1996)依据幼虫和成虫特征将Coscinocephalus从圆头犀金龟族Cyclocephalini转移到禾犀金龟族Phileurini。Jameson和Wada(2004)提到Neohypus和Melanhyphus的分类地位在圆头犀金龟族或类蛀犀金龟族之间还存在争议。Endrödi(1968, 1985)将Pucaya划分到扁犀金龟族,Paucar-Cabrera和Moore(2018)根据形态学和分子数据将Pucaya从扁犀金龟族转移到圆头犀金龟族。Costa等(2021)研究Gibboryctes模式种的正模标本和其他物种标本后,认为蛀犀金龟族下Gibboryctes为犀金龟族Dynastini下Megasoma的属级异名。
犀金龟亚科和丽金龟亚科Rutelinae之间一些属的归属问题长期存疑,如Oryctomorphus兼有丽金龟亚科(Machatschke, 1972)和犀金龟亚科的特征(Endrödi, 1985),其分类地位一直在两个亚科之间来回变化。此外,Acrobotbia的分类位置存在争议,有时属于丽金龟亚科丽金龟族Rutelini,有时属于犀金龟亚科扁犀金龟族(Ohaus, 1912; Jameson, 1997),Jameson等(2002)对其进行修订,依据触角特征将其转移到犀金龟亚科圆头犀金龟族,但未有分子数据研究佐证。
世界犀金龟亚科的分类学研究已有200多年历史,主要分为传统分类学研究、生物地理区系研究以及近些年的分子生物学研究三个方向。针对该类群的分类学和生物学研究,至今超过180名学者发表过900余篇论文和专著。1847年出版的HandbuchderEntomologie第一卷中,Burmeister详细描述了该亚科世界范围内记载350个种的外部形态特征、分布地等信息,Gemminger和Harold(1868-1876)在首次出版的世界鞘翅目名录里记载518种犀金龟,1910-1937年间出版的鞘翅目名录数量增长到1 335种(Kohlmann and Morón, 2003),1985年Endrödi在TheDynastinaeoftheWorld记录1 422种。随着分类特征的确定,众多学者进行了物种描述工作。LeConte(1848-1866)、Fairmaire(1856-1901)、Kolba(1883-1914)、Blackburn(1888-1909)、Prell(1909-1937)、Lea(1918-1926)、Endrödi(1938-1983)等分别对世界各地区的犀金龟进行物种描述和修订,记录很多新属、新种甚至新族,编写地方性专著和名录等,进一步完善了犀金龟亚科区系分布记录。
分类学家多以地理区系为研究单位,并对部分类群进行了世界范围的系统修订。犀金龟布及全球,以热带区和新热带区种类最为丰富,古北区分布较少(Endrödi, 1985)。Arrow(1910)对东洋区中的印度、斯里兰卡及缅甸地区的犀金龟进行了全面考察,共描述46种,并对形态、性二型、生物学等进行了深入的研究。Kolba和Paulian自1914年开展了非洲不同地区的分类研究,但因生境较差,此后调查较少。Carne(1957)将Blackburn和Lea的研究总结于Australian Dynastinae中。1985年Endrödi对澳洲区物种进行了整理,发现该地区种类较多,此后相关修订研究缺乏。新热带区主要有Restrepo-Giraldo和Heyller(2003)、Gasca-Alvarez和Amat-Garcia(2010)、López-García等(2015)先后整理哥伦比亚犀金龟物种列表、成虫检索表和地理分布信息等;Cave和Ratcliffe(2009)研究和记录南美洲北部犀金龟物种多样性和时空分布等信息,并厘清了该地区犀金龟亚科下的属种。新北区主要有Miguel等(2021)整理墨西哥瓦哈卡地区6族22属 98种列表。近年来,美国学者Ratcliffe(1977-2021)在犀金龟的系统分类工作及生物地理分布工作的贡献尤为突出,整理巴拿马、墨西哥、秘鲁等地区物种列表,统计地理分布等信息,并对Endrödi等人的工作进行了总结和修订;巴西学者Grossi在2009-2020年期间,对Altitatiayus、Philoscaptus和Megasoma物种列表进行整理。因其在美洲有丰富的种类,一些学者已经对该亚科进行了较详细的系统分类学研究,尽管在过去20年中对新热带区犀金龟亚科的了解有所提高,但仍需对南美洲地区进行物种列表整理和分类研究(Cave and Ratcliffe, 2015)。
近年来,分子生物学技术研究不断发展,在探究犀金龟亚科系统分类和系统发育等方面已有相关应用。Smith等(2006)基于18S、28S核糖体DNA序列系统发育关系分析,结果表明犀金龟亚科与丽金龟亚科亲缘关系更近。犀金龟和蜣螂的头、前胸背板具角状突起,而目前基于对此性状的研究仅限于蜣螂,Ohde等(2018)通过RNA干扰技术分析发现犀金龟与蜣螂角状突起的发育有相似之处。Filipovic' 等(2021)报道了犀金龟亚科第一个有关椰蛀犀金龟Oryctesrhinoceros完整线粒体基因组,为调控椰蛀犀金龟生长和发育等方面提供理论支撑。线粒体基因组的引入,为解决该亚科分类学、系统发育和有效防治提供分子证据。Rowland和Miller(2012)基于形态学、生物地理学和分子特征分析开展了犀金龟族系统发育研究,获得一棵稳定拓扑结构的系统发育树,为该族的分类和进化研究提供证据。Yang等(2021)运用特异性位点扩增片段测序技术分析叉犀金龟属Trypoxylus地理分布时,发现地理因素影响种群内同质性和种群间差异,该研究为解决叉犀金龟属分类和种群遗传分化问题提供了理论支撑。Dutrillaux 和Dutrillaux (2013)通过研究犀金龟属及近缘类群的精细胞、中肠细胞和卵细胞发现,犀金龟属物种的核型与其他同亚科物种不同,且该属南美洲与北美洲物种在第9对染色体和性染色体上有差别,表明犀金龟属起源于南美洲及向北迁徙并在中美洲地区形成物种(亚种)。分子生物学技术的应用,为犀金龟亚科与近缘亚科系统发育关系等问题的解析提供了新方法,但族级、属级系统关系等方面研究涉及较少。
2.2 幼虫及蛹的研究
随着犀金龟物种的发现,幼虫及蛹的特征描述也逐渐增多。单独使用幼虫特征或与成虫特征结合使用,为建立自然类群以及研究不同分类学水平的系统发育关系方面提供了比成虫形态学更可靠的依据(Ritcher, 1966; Verdú,etal., 2004; Micó,etal., 2008;ípeketal., 2009)。早期学者基于Ritcher(1944, 1966)提出的犀金龟亚科幼虫术语,描述了椰树和棕榈树上的幼虫(Endrödi, 1974; Bedford, 1979)。1990年以来,圆头犀金龟族80%种类的幼虫和蛹被描述(Mooreetal., 2018)。Ocampo和Moron(2004)、Neita-Moreno和Ratcliffe(2011)、Ratcliffe和Curoe(2011)、Ibarra-Polesel等(2017)先后描述了部分禾犀金龟族蛹期的关键特征,López-Colón(2003)、Neita-Moreno和Orozco(2009)、Pardo-Locarno等(2009)对部分蛀犀金龟族幼虫和蛹特征进行了描述。直到2006年,Pardo-Locarno和Morón才描述了彩犀金龟族Agaocephalini第一个幼虫Lycomedeshirtipes。虽然已有一些关于幼虫及蛹的研究,但一些学者对于幼虫鉴定特征在族级阶元中的使用意义持怀疑态度。Ibarra-Polesel等(2020)提到鉴定圆头犀金龟族Stenocrates成虫时,常使用(或仅使用)扁犀金龟族的分类鉴定特征,但其幼虫的族级特征则更符合圆头犀金龟族。还有一些属依据幼虫特征分类地位发生改变。如Platyphileurus从禾犀金龟族转移到蛀犀金龟族(Albertonietal., 2014),Diloboderus从扁犀金龟族转移到彩犀金龟族(Pardo-Locarno and Morón, 2006; Neita-Morenoetal., 2014;Morón and Grossi, 2015)。
2.3 存在主要问题
目前,在世界范围内,犀金龟亚科仍以分类学研究为主,虽然已具备一定基础,但仍存在问题。主要包括以下几方面:1)世界区系调查不均衡。美洲地区调查较多,而非洲地区很少。2)犀金龟亚科低级分类阶元不稳定。属级分类阶元不明确,主要原因是部分族级分类阶元单系性尚未明确。3)形态特征研究不足。犀金龟角状突起外形独特,与蜣螂等亚科相比,研究不足。4)研究方法运用少。早期以传统分类方法为主,较少对幼虫和蛹等特征进行描述,调查种类丰富地区尚缺乏运用现代研究方法(包括分子系统学、几何形态学和三维形态学等)开展的研究。
3 中国犀金龟亚科研究情况
3.1 成虫研究历史
我国早期的犀金龟研究主要是由国外学者开展的,如Guérin(1834),Hope(1845),Fairmaire(1878, 1887, 1898),Nonfried(1890),Frivaldszky(1890)等人描述了一些新种,为开展国内犀金龟亚科研究奠定基础。
中国地域辽阔,气候复杂,但犀金龟种类不多,仅占世界已知种的4%左右。1937年,胡经甫最早记录我国犀金龟7属8种。黄可训(1952)针对胡氏“中国昆虫目录”进行了补充,共5属12种。章有为先生对我国犀金龟亚科分类研究做出突出贡献,修订了我国分布的犀金龟2属:异爪犀金龟属Heteronychus和蔗犀金龟属Alissonotum,先后在《中国北方常见金龟子彩色图鉴》、《中国珍稀昆虫图鉴》、《海南森林昆虫》和《福建昆虫志》第六卷中均有对犀金龟的记述(章有为, 1983; 1984; 1997; 1999; 2002a; 2002b)。这些地方志或专著的记述,为丰富我国犀金龟亚科研究做出了重要贡献。基于以上研究,章有为先生对我国犀金龟亚科昆虫较系统而全面的总结,共记述4族13属33种,包括3个新种(章有为, 1991)。截止到2021年底,我国记录犀金龟亚科共67种(表3),隶属于5族17属,即:圆头犀金龟族:1属1种;犀金龟族:3属13种;蛀犀金龟族:5属12种;禾犀金龟族:7属35种;扁犀金龟族:1属6种。其中,双叉犀金龟、阔胸禾犀金龟和中华晓扁犀金龟Eophileuruschinensis为中国广布,其他均为狭域分布。此外,我国的东洋区处于热带和亚热带地区,植被丰富,犀金龟的食物充足,是中国犀金龟主要分布及引起农林灾害的区域,同时也是犀金龟害虫治理和益虫开发的热点地区(章有为, 1991)。

表3 中国已记录犀金龟亚科昆虫物种列表
3.2 幼虫和蛹的研究
中国犀金龟种类虽少,但其幼虫严重危害我国农林业的发展,了解幼虫和蛹的形态特征使有效防治害虫成为可能。沈阳农业大学地下害虫研究组先后在1974年和1982年描述4属4种犀金龟幼虫。张芝利(1984)在《中国经济昆虫志》记录6属8种幼虫,掌握幼虫和蛹期形态特征和生活史等信息,探索有效抑制犀金龟幼虫生长发育的生物防治方法,为促进农林发展提供新机遇(崔景岳等, 1981; 张中润等, 2004)。单立华等(1998)对双叉犀金龟幼虫营养成分进行分析,为后续开展化学成分、结构、利用方式等提供依据。现阶段规模化养殖已成为一种重要的商业模式,由于一些犀金龟幼虫生存环境条件较高,人工饲养难度较大,还有待进一步研究。
3.3 我国研究主要问题
我国犀金龟亚科研究在章有为等学者的推动下,取得了一定研究成果,主要以基础分类为主。目前我国已记载犀金龟67种(亚种),但有些物种的分类地位尚有争议,该亚科的系统发育工作进展缓慢,现阶段我国对犀金龟亚科昆虫的地理分布调查也相对薄弱。针对目前的研究情况,我国犀金龟亚科的研究所面临的主要问题包括:1)缺少本底资源调查:以前针对犀金龟调查集中于我国热带和亚热带地区,新疆、西藏等地区较国内其他内陆地区相比,发现物种较多,但调查较少,且犀金龟亚科物种名录尚未完成。2)资源开发研究少:之前研究多集中于生物学和形态学特性上,而药用价值(抗氧化、抗糖等)仅有少数报道,还缺乏具体研究。3)应用和经济价值研究较少:例如目前有研究显示双叉犀金龟幼虫可用于处理废弃物等,但目前还缺少更加深入、详细、完整的研究。
4 经济意义
犀金龟经济昆虫资源丰富,观赏、药用、食用、仿生、生态系统平衡等方面成为经济开发利用的主导向,极具商业化前景。全球经济一体化和国际贸易频繁往来,促使农林产品大量流动,为外来物种提供了传播途径,生物入侵概率大大增加,检疫及进出口管理成为防范生物入侵的重要手段(鞠瑞亭等, 2012)。此外,部分犀金龟也是农业上的重要害虫,科学防治变得尤为关键。近年来,自然灾害和人类活动加剧,犀金龟物种多样性遭到破坏,究其原因主要是对昆虫多样性保护的认识不足(New, 2005)。
4.1 观赏价值
犀金龟成虫多数体型巨大,是鞘翅目乃至昆虫纲内“巨虫”家族之一(章有为, 1991)。头部和前胸背板多数具角突,外形独特,极具观赏价值,激起昆虫爱好者收集、饲养、销售、装饰服装的热潮。在马来西亚和菲律宾等地,橡胶木犀金龟Xylotrupesgideon被用于赌博竞技。南非地区将犀金龟颜色鲜艳的部位(鞘翅、前胸背板、足)做成饰品或装饰服装(Ratcliffe, 2006)。借助改良幼虫饲料和温度诱导缩短化蛹时间的方法,大规模饲养椰蛀犀金龟成为可能(Schipper, 1976)。犀金龟的观赏价值给经济发展注入了新活力,但伴随非法贸易的盛行,给当地的生态环境造成了一定威胁(Ichiishietal., 2019)。
4.2 药用和食用价值
犀金龟营养成分丰富,在药用和食用上被广泛应用。双叉犀金龟指名亚种Trypoxylusdichotomusdichotomus内含独角仙素,对白血病的治疗具有重要作用。使用抗生素常出现耐药现象,椰蛀犀金龟含有的抗菌肽Ac-AMP2具易降解、广谱抗菌等作用,拓宽了抗菌抑菌研究新思路(Hemmietal., 2003)。有相关研究表明,部分犀金龟的蛋白质、脂肪酸、微量元素等含量远高于猪肉和鸡蛋等食物,韩国已将双叉犀金龟列入食用昆虫目录(Ghoshetal., 2017)。很多植食性犀金龟亚科幼虫在3龄时个体较大,且数量巨多,成为重要的食物资源。如墨西哥恰帕斯州拉坎顿人食用Cyclocephalafasciolata幼虫、蛹和成虫(Ramos-Elorduy and Pino-Moreno, 2002),在南非人们食用Oryctesmonoceras的幼虫和成虫(Bodenheimer, 1951)。随着犀金龟药用和食用价值的开发,商业需求量增多,同时面临物种数量减少的情况下,规模化饲养成为现阶段的可行性方案。
4.3 仿生应用
仿生材料基于高强度、轻量型等特性快速发展,已成为现阶段研究热点,加强犀金龟在仿生材料应用的探索,推动仿生材料在经济发展中发挥更大作用。目前犀金龟亚科在力学、生物学特性和昆虫表皮等领域已有相关报道:利用双叉犀金龟前翅小柱分布及其前翅表面微细构造特点为航空航天的构造设计提供新思路(Chenetal., 2002);Lee等(2017)采用聚焦离子束原位操作的方法测试双叉犀金龟头部角突的机械性能,研究表明头部角突具有弹性模量高及抗断裂拉伸的特点,为研制该特点的仿生材料提供新思路。此外,双叉犀金龟表皮基因与几丁质结合的特性,为新型蛋白质替代物和昆虫表皮仿生材料提供优质蛋白质资源(叶长青等, 2021; 刘晶等, 2022)。
4.4 维持生态系统平衡
以犀金龟为代表的昆虫群落是生态系统重要组成部分,在维持系统结构和功能方面发挥重要作用。双叉犀金龟幼虫利用多糖消化酶等在森林环境中发挥分解作用,也可通过腐烂沉积物和肠道细菌调节土壤菌落组成和多样性,改善土壤环境,促进生态系统循坏(Wadaetal., 2014, Eoetal., 2017)。圆头犀金龟族是棕榈类植物重要的传粉者,其幼虫在土壤中发育,以有机物为食,有助于养分的循环(Mooreetal., 2018)。近年来,经济快速发展促使昆虫栖息地被开发,生境被破坏,部分犀金龟物种数量下降,出现生态系统失衡现象(Pathaniaetal., 2015),日本过度进口犀金龟已严重破坏当地生态环境和物种多样性(New, 2005)。
4.5 保护价值
经济全球化带来了机遇,同时给昆虫多样性带来了危机,有效协调经济发展与物种多样性之间的关系,成为当今多样性保护的主要矛盾。在自然灾害和人类活动等因素的影响下,物种多样性急剧下降,增强保护意识迫在眉睫(叶水送等, 2013)。近年来,气候干旱、洪涝等自然灾害频发,昆虫资源受损严重(尤民生, 1997)。犀金龟作为观赏昆虫深受爱好者的喜爱,收藏、销售等贸易活动盛行,导致昆虫数量急剧减少(Ichiishietal., 2019)。人类受利益驱使,过度放牧,采摘野菜、中药材等植物资源,此类行为严重破坏了昆虫栖息地环境(曹巍等, 2019)。值得注意的是,梳理国内近年昆虫群落多样性调查发现,犀金龟记载种类逐年减少,已将部分种类列入《国家重点保护野生动物名录》、《国家保护的有重要生态、科学、社会价值的陆生野生动物名录》和地方重点保护野生动物名录。针对上述情况,此后应重点保护犀金龟昆虫的生存环境,制定相关法律法规,积极宣传昆虫保护知识,开展犀金龟生物学习性等科学研究,为保护昆虫资源、维持物种多样性等提供理论和制度支撑(张茂林和王戎疆, 2011)。
4.6 危害
犀金龟在创造巨大经济价值的同时,部分类群对生态系统平衡、生物多样性和进出口检疫等方面造成了一定的破坏,并对人类经济利益造成损失(New, 2005; Ratcliffe, 2006; Ruzzier, 2020)。椰蛀犀金龟是世界范围内椰子树上的重要害虫(Schipper, 1976)。双叉犀金龟常表现出啃食树皮的行为,是日本的重要检疫害虫(Ichiishietal., 2019)。外来生物入侵破坏本地植物和昆虫群落,影响本地昆虫的群落结构和食性,破坏当地生物多样性分布格局。Cyclocephla和Adoryphorus是作物或牧草害虫(Bergetal., 2014; Fuhrmannetal., 2019),Golofa在竹子生长和觅食(Cespedes and Ratcliffe, 2010),影响竹子生长,Heteronychuslicas损害山药和甘蔗(Jerath, 1966),Temnorhynchusbaal幼虫严重危害埃及草莓等(Shehataetal., 2020)。这些害虫的为害,给农业发展造成了重大的经济损失,有效防治这些害虫,加大监测监管和进出口检疫力度,基础在于做好分类工作,提供研究基础。
5 研究意义及展望
通过前文整理的犀金龟亚科分类研究历史,分析现阶段存在的主要问题,本研究认为,要推动犀金龟亚科系统分类研究,需要结合几何形态学、分子生物学等新技术方法,重点探究犀金龟亚科与近缘亚科、族级和属级系统发育关系,并解决部分种间因形态差异小而无法厘定以及亚族植食性关系等问题,进一步对犀金龟亚科分类系统进行修订。其次,世界犀金龟种类繁多,应加强对非洲区、澳洲区、东洋区等地区的调查,进一步丰富世界物种记录,重点研究我国的犀金龟种类。第三,积极开展犀金龟外形独特、药用价值和仿生等资源研究,推动农林经济发展和物种多样性保护,同时应加强对外来生物入侵的监管和检测。第四,重视幼虫及蛹形态特征和生物学习性等相关研究,为农林业害虫防治、物种分类地位确定提供更多证据。
针对我国犀金龟亚科研究状况,在加强我国热带和亚热带地区调查的同时,应关注我国内陆地区的种类,分析我国犀金龟物种多样性及地理分布格局,为进一步开展系统发育、生物地理学、生物防治、仿生学和开发利用等方面研究提供基础资料。
猜你喜欢
生物技术通报(2023年2期)2023-03-07 12:56:26
玩具世界(2022年3期)2022-09-20 01:48:20
甘肃教育(2020年2期)2020-09-11 08:01:48
四川动物(2020年3期)2020-06-11 11:08:28
吉林林业科技(2020年2期)2020-04-15 07:25:22
武夷科学(2019年2期)2019-12-20 08:39:16
小学生优秀作文(低年级)(2018年9期)2018-09-10 05:47:38
小学生必读(低年级版)(2018年11期)2018-03-13 00:54:24
武夷科学(2017年0期)2017-03-10 10:03:31
新疆农垦科技(2014年9期)2014-02-28 19:20:55
