
3种对虾病毒囊膜蛋白对假型昆虫杆状病毒的包装与对虾组织亲嗜性的影响❋
2023-10-18 04:04:04马振龙陶奕文赵亚琦郭华荣
中国海洋大学学报(自然科学版) 2023年11期
马振龙, 陶奕文, 赵亚琦, 宋 柳, 郭华荣,2❋❋
(1. 中国海洋大学海洋生命学院, 海洋生物遗传与育种教育部重点实验室, 山东 青岛 266003;2. 中国海洋大学海洋生物进化与多样性研究所, 山东 青岛 266003)
病毒介导的基因转移技术具有毒性小、致死率低、操作简单和转移效率高等优点[1]。其中,哺乳动物病毒介导的有逆转录病毒、慢病毒、腺病毒和腺相关病毒表达系统,昆虫病毒介导的有昆虫杆状病毒表达系统。根据病毒是否具囊膜,上述病毒又可分为两类:腺病毒和腺相关病毒为不具囊膜病毒,病毒的衣壳是裸露的,衣壳蛋白决定了病毒的宿主专一性;而逆转录病毒、慢病毒和昆虫杆状病毒的最外层由囊膜包被,为具囊膜病毒,其囊膜蛋白在病毒侵染宿主细胞的过程中发挥识别和结合宿主受体的作用,决定着病毒侵染的宿主范围及组织和细胞的特异性[2]。有研究表明,一种病毒的囊膜中可以插入其他病毒的囊膜蛋白,这种插入了另一种病毒囊膜蛋白的病毒被称为假型病毒,假型病毒往往因为引入了其他病毒的囊膜蛋白而改变了其本身的宿主亲嗜性[3]。本实验室Pu等[4]最早通过共转染的方法,将对虾白斑综合征病毒(WSSV)的囊膜蛋白VP19和VP28引入到逆转录病毒的囊膜中,发现所构建的假型逆转录病毒具有了对虾细胞亲嗜性,可以感染对虾原代培养血淋巴细胞,但是感染效率不高。随后,本实验室Chen等[5]采用类似的方法将VP19和VP28引入到慢病毒的囊膜中,发现所构建的假型慢病毒可以感染对虾原代培养血淋巴细胞和淋巴(Oka)器官原代培养细胞,感染和表达效率明显优于逆转录病毒。但上述两种假型病毒在对虾细胞中的感染和表达效率仍远低于哺乳动物细胞。与哺乳动物相比,昆虫和对虾的亲缘关系更近,本实验室Wu等[6]采用类似的方法,将对虾WSSV病毒囊膜蛋白VP28引入昆虫重组杆状病毒(Bacmid-GUS)的囊膜中,发现VP28可以显著提高上述杆状病毒的包装效率及其在对虾中的亲嗜性,尤其是对虾成体组织,在几乎所有染毒对虾的组织中均可检测到β-葡萄糖苷酸酶基因(GUS)的表达,但是具有明显的组织特异性。本文作者推测,VP28-假型昆虫杆状病毒所表现的对虾组织特异性可能与VP28的引入有关,但是VP28对于上述假型病毒的对虾组织特异性的贡献有多大以及对虾WSSV病毒其他囊膜蛋白是否有相似的贡献尚不清楚。
VP19和VP24是对虾WSSV病毒另外两种重要的囊膜蛋白。VP19蛋白由121个氨基酸组成,存在两个跨膜结构域,可锚定在WSSV囊膜中发挥作用[7]。在病毒中和实验中,抗rVP19血清可显著降低WSSV感染对虾的死亡率,表明VP19可能在对虾的感染中起重要作用[8-9]。Pu等[4]和Chen等[5]将VP19分别引入到逆转录病毒和慢病毒表达系统中发现,两种方式均明显提高了假型病毒对对虾细胞的亲嗜性。因此,本文作者关注到VP19对于假型昆虫杆状病毒(Bacmid-GUS/VP19)的对虾组织特异性的贡献如何这一问题。VP24蛋白含有208个氨基酸,其N端存在一段跨膜结构[10]。Niu等[11]发现对虾MarsupenaeusjaponicuspIgR是一种WSSV受体,与囊膜蛋白VP24相互作用,WSSV可通过pIgR-CaM-网格蛋白的内吞途径进入对虾细胞。有研究表明,斑节对虾细胞表面的几丁质结合蛋白(PmCBP)可以与VP24结合,促使WSSV进入对虾细胞内[12]。因此,VP24成为本研究第二个关注的对虾WSSV囊膜蛋白。Syed Musthaq等[13]构建了对虾WSSV囊膜蛋白VP28的重组杆状病毒表达载体,发现过表达的VP28蛋白定位于昆虫包装细胞的细胞膜上,为VP28可被引入重组杆状病毒的囊膜上提供了直接的实验证据。而将VP19和VP24引入假型昆虫杆状病毒时是否也能定位于昆虫包装细胞的细胞膜上,是否也可以提高假型病毒的包装效率以及其对于假型病毒的对虾组织特异性的贡献如何,尚不清楚。
因此,为了探究VP28以及另外两种囊膜蛋白VP19和VP24对该假型重组杆状病毒的对虾组织特异性的贡献,本研究拟通过共转染的方法分别将上述3种对虾病毒囊膜蛋白引入到昆虫杆状病毒(Bacmid-GUS)的囊膜中,然后比较上述3种假型病毒的包装效率及其在对虾中的组织特异性,并进一步分析上述3种囊膜蛋白在包装细胞(Sf9)中的亚细胞定位。本研究成果将拓展人们对于对虾WSSV病毒囊膜蛋白的生物学功能的认知,并为假型昆虫杆状病毒介导的基因转移与表达系统的构建及其在对虾活体基因转移中的应用提供指导。
1 材料与方法
1.1 对虾和小龙虾
健康的刀额新对虾(Metapenaeusensis),体质量为15~20 g,体长为10~15 cm,购自山东省青岛市南山水产品市场,于过滤的充气海水中暂养,每日换水1次,禁食,用于病毒感染实验。
克氏原螯虾(Procambarusclarkii)又称小龙虾,体质量为20~25 g,体长为13~20 cm,购自山东省青岛市南山水产品市场,于浅层的曝气自来水中暂养。取感染WSSV致死的冻存小龙虾的肌肉组织,制备病毒粗提液,以肌肉注射的方式感染小龙虾,制备WSSV染毒小龙虾,用于提取WSSV全基因组。
1.2 Sf9细胞
Sf9细胞系来源于草地贪夜蛾(Spodopterafrugiperda)的卵巢组织,复苏后置于含10%胎牛血清(FBS)的SIM培养基(Sino biological公司)中,于28 ℃培养箱中培养,每隔2~3 d传代一次,用于杆状病毒的包装和滴度测定。
1.3 表达质粒
基础质粒pcDNA3.1/V5-His购自Invitrogen公司,克隆质粒pMD19-T购自TaKaRa公司,表达质粒pcDNA3.1-VP19、pcDNA3.1-VP28、pcDNA3.1-VP28-△TAA/mCherry和昆虫杆状病毒GUS报告基因重组表达质粒Bacmid-GUS为实验室前期构建[4,6]。
表达质粒pcDNA3.1-VP24的构建方法如下:根据对虾WSSV病毒囊膜蛋白VP24的开放阅读框(ORF)序列设计特异性引物,并在引物两端加上酶切位点EcoRⅠ和BamHⅠ,引物序列见表1。WSSV染毒小龙虾的肌肉组织总DNA的提取采用TIANamp Genomic DNA Kit试剂盒(TianGen公司),利用超微量分光光度计和琼脂糖凝胶电泳检测DNA的质量和浓度。以WSSV染毒小龙虾的肌肉组织总DNA基因组为模板,使用Premix TaqTM试剂盒(TaKaRa公司)扩增VP24基因的编码框,PCR产物凝胶电泳,切胶回收并与克隆载体pMD19-T连接,测序验证后,利用EcoR Ⅰ和BamHⅠ双酶切,将VP24基因片段定向插入表达质粒pcDNA3.1/V5-His,构建pcDNA3.1-VP24重组表达质粒。

表1 PCR引物Table 1 PCR primers used in this study
为验证对虾病毒囊膜蛋白(VP19和VP24)在包装细胞(Sf9)中的亚细胞定位,本文进一步构建了红色荧光蛋白(mCherry)标记的VP19囊膜蛋白的融合表达重组质粒:pcDNA3.1-VP19-△TAA/mCherry,以及绿色荧光蛋白(eGFP)标记的VP24囊膜蛋白融合表达重组质粒:pcDNA3.1-VP24-△TAA/eGFP,所用引物序列见表1。其中,mCherry和eGFP均标记在上述2个囊膜蛋白的羧基端(C端),并分别删除了其前面的囊膜蛋白的终止密码子(△TAA)。将无终止密码子的VP19和VP24基因片段分别插入到BamHⅠ和EcoRⅠ 之间,将mCherry和eGFP报告基因片段分别插入到其后的EcoRⅠ和NotⅠ之间。
上述以pcDNA3.1/V5-His为基础质粒的重组表达质粒的纯化采用FastPure EndoFree Plasmid Maxi Kit试剂盒(诺唯赞生物公司)。Bacmid-GUS重组质粒的纯化采用PureLinkTMHiPure Plasmid DNA Purification Kits试剂盒(Invitrogen公司),直接从Bacmid-GUS重组菌株DH10BacTM中提取[6]。所使用的溶菌肉汤(LB)液体培养基中含50 μg/mL卡那霉素、7 μg/mL庆大霉素和10 μg/mL四环素。各种表达质粒冻存于-20 ℃备用。
1.4 3种假型昆虫杆状病毒的包装、浓缩和纯化
3种假型昆虫杆状病毒的包装、浓缩和纯化参照Wu等[6]的方法进行,采用了Wu等[6]所报道的VP28-假型杆状病毒包装时的最佳共转染条件,包括囊膜蛋白表达质粒与昆虫杆状病毒表达质粒(Bacmid-GUS)的共转染比例为3∶1,转染试剂(L)∶质粒(g)=5∶1,以及转染后第5天收集病毒等。其中,3种对虾WSSV囊膜蛋白(VP19、VP24和VP28)对于假型杆状病毒的包装效率的影响实验在48孔细胞培养板中进行,分别比较了其在最佳转染条件和非最佳转染条件下的包装效率。具体方法如下:将Sf9细胞均匀接种到48孔板中,待细胞铺板率达到80%~90%时,采用Cellfectin Ⅱ Reagent(Gibco公司)进行转染。其中,最佳转染条件为:于1.5 mL离心管中,在终体积为200 μL的无血清无抗生素的Grace昆虫培养基(供应商:Gibco公司)中溶解0.375 μg Bacmid-GUS重组质粒、0.125 μg囊膜蛋白表达质粒pcDNA3.1-VP19(或pcDNA3.1-VP24,或pc-DNA3.1-VP28)和2.5 μL Cellfectin Ⅱ转染试剂,混匀,室温孵育30 min;弃除旧培养基,将200 μL/孔上述脂质体-DNA复合物加入到48孔板中,于28 ℃培养4~5 h后,更换培养液为含10% FBS的SIM培养基;继续孵育5 d后,进行X-gluc染色,检测GUS基因的表达。与此不同的是,非最佳转染条件为:每孔0.15 μg Bacmid-GUS重组质粒、0.05 μg囊膜蛋白表达质粒和1 μL CellfectinⅡ转染试剂,其他条件同上。
GUS基因的表达产物为β-葡萄糖苷酸酶,是一个水解酶,X-gluc(5-溴-4-氯-3-吲哚-β-葡萄糖苷酸)是该酶的水解底物,可被该酶水解产生无色的吲哚氧基中间产物,吲哚氧基中间产物可被进一步氧化成为蓝色沉淀,沉积于组织和细胞的原位,因而可根据X-gluc染色的结果来检测GUS基因的表达。X-gluc染色方法:首先弃除旧培养基,每孔加200 μL PBS溶液(8 mg/mL NaCl、0.2 mg/mL KCl、3 mg/mL Na2HPO4(12H2O和0.2 mg/mL KH2PO4,pH=6.8)洗涤1次;其次加125 μL固定液,固定5 min,用PBS洗涤2次;然后加入125 μL X-gluc细胞染液(含1 mg/mL X-gluc、5 mmol/L K3Fe(CN)6、5 mmol/L K4Fe(CN)6和2 mmol/L MgCl2的PBS溶液)[6],孵育过夜;最后第二天弃染液,用PBS洗涤1次,加500 μL PBS后,观察拍照。
3种假型病毒:Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28的包装实验在直径10 cm的细胞培养皿内进行,采用前面提及的最佳转染条件,不同的是,在终体积为500 μL的无血清无抗生素的Grace培养基中溶解15 μg Bacmid-GUS质粒、5 μg pcDNA3.1-VP19(或pcDNA3.1-VP24,或pcDNA3.1-VP28)表达质粒以及100 μL Cellfectin Ⅱ转染试剂。转染后继续孵育5 d后,收集细胞培养基上清,离心去除细胞碎片,获得P1代病毒上清;将6个直径10 cm细胞培养皿中分别接种上400 μL P1代病毒上清,感染5 d后,收集所有培养基上清并离心,获得P2代病毒上清。
病毒的浓缩方法如下:用0.45 μm滤膜过滤收集到的P2代病毒上清,并转移到超速离心管中;在离心管底部加入25%的蔗糖溶液,于4 ℃下80 000g离心75 min,进行病毒的浓缩和纯化,加入100 μL的无菌PBS溶液重悬病毒颗粒,分装储存于-80 ℃备用。
浓缩后的假型病毒的滴度测定方法如下:将Sf9细胞接种到96孔板,待细胞密度达到80%~90%时,分别接种经无血清无抗生素的SIM培养基梯度稀释(10-1~10-8)的假型昆虫杆状病毒:Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28,每孔添加100 μL病毒稀释液,于28 ℃培养箱中培养5 d后,进行X-gluc染色,检测GUS基因的表达。转导单位(TU)指可以感染并进入到靶细胞中的病毒基因组数。病毒滴度(TU/mL)=阳性细胞百分数×细胞总数×稀释倍数/病毒稀释液体积(mL)。
1.5 囊膜蛋白VP19、VP24和VP28在包装细胞Sf9中的细胞膜定位
将Sf9细胞接种到12孔培养板中的细胞爬片上,待细胞铺板率达到70%~80%时,将Bacmid-GUS分别同pcDNA3.1-VP19-△TAA/mCherry、pcDNA3.1-VP24-△TAA/eGFP和pcDNA3.1-VP28-△TAA/mCherry按照最优转染条件共转染到Sf9细胞中。转染96 h后在激光共聚焦显微镜下观察上述3种融合表达的囊膜蛋白在细胞中的表达并拍照。
1.6 3种假型昆虫杆状病毒感染对虾成体
根据3种假型昆虫杆状病毒的滴度来确定其在对虾成体组织中的最高和最低注射剂量,如表2所示。对照组对虾注射PBS溶液,实验组分别注射三种假型病毒。采用肌肉注射的感染方式,注射部位为第一、二腹足之间。注射完毕后,摁压针眼10 s左右以防止病毒溶液漏出。感染5 d后,分别取染毒对虾的心脏、Oka器官、肠、鳃和肌肉组织,置于1.5 mL EP管中;然后加入X-gluc组织染液(1 mol/L磷酸钠缓冲液、0.5 mol/L Na2EDTA、10% Triton X-100、50 mmol/L K3Fe(CN)6和50 mg/mL X-gluc)[6],浸没组织;置于37 ℃培养箱孵育12 h,弃除染液,加PBS洗涤后,于体视显微镜下观察并拍照。

表2 3种假型病毒在对虾成体组织中的注射剂量Table 2 The injection doses of three pseudotyped viruses in adult shrimps
为了检测假型病毒在对虾组织中是否具有复制和增殖能力,选择上述3种假型昆虫杆状病毒的最大剂量感染对虾,取GUS表达阳性的组织进行透射电镜分析。其中,假型病毒Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28的对单个对虾最大剂量分别是1×109、1×109和1×1011个转导单位,前2种假型病毒的GUS表达阳性组织为心脏和肌肉,另外一种假型病毒的GUS表达阳性组织为心脏、鳃和肌肉。分别取大小为1~2 mm3的组织块,于4 ℃预冷的2.5%戊二醛固定液中4 ℃固定12 h,然后送至青岛大学医学院进行透射电子显微镜分析和观察。
1.7 统计学分析
所有实验至少重复3次。应用软件Image J计算3种假型昆虫杆状病毒在Sf9细胞中的感染与表达效率。阳性GUS信号的百分比通过以下公式计算:
(具有GUS信号的阳性细胞数/细胞总数)×100%。
采用SPSS软件对数据进行统计学分析。
2 实验结果
2.1 真核表达载体pcDNA3.1-VP24的构建
如图1所示,成功扩增得到对虾WSSV囊膜蛋白VP24基因的编码框,大小位于500~750 bp之间,符合预期(见图1A)。测序结果如图1C所示,VP24扩增片段与WSSV印度株的VP24基因序列一致(见图1D)。这表明含VP24的表达质粒:pcDNA3.1-VP24已构建成功(见图1B)。
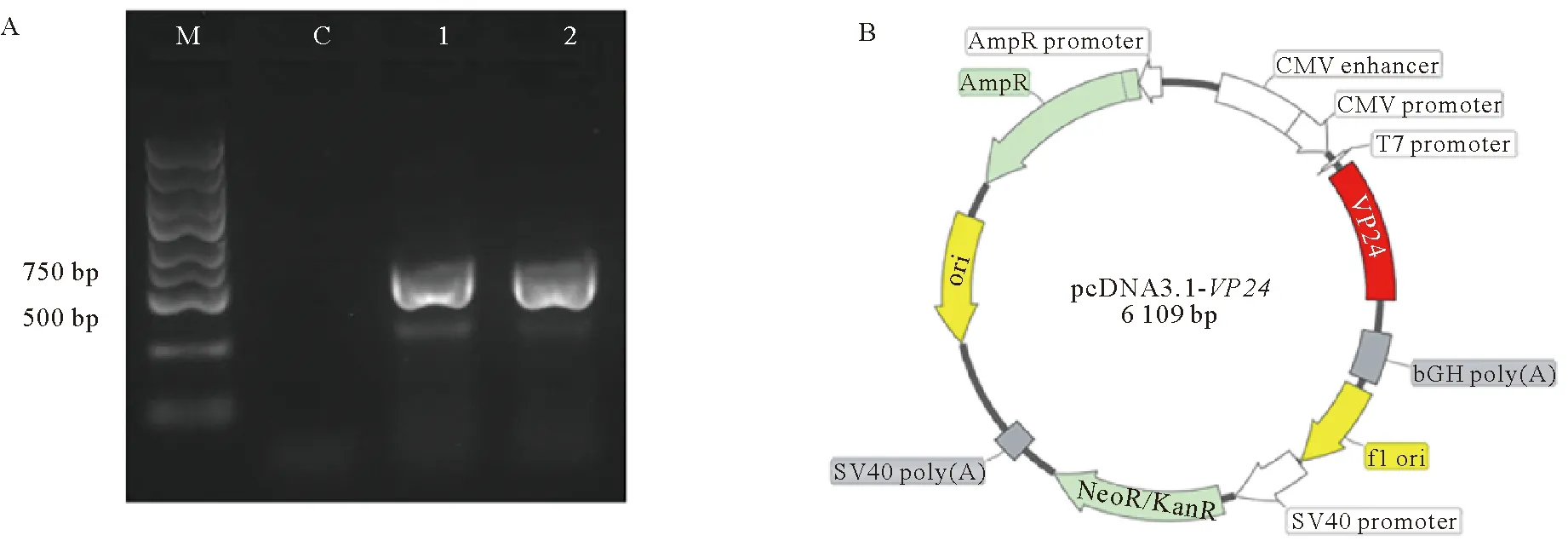
(A. VP24基因的PCR扩增产物的凝胶电泳结果。其中:泳道M为Trans2K Plus DNA maker;泳道1和2均为VP24扩增产物。B. pcDNA3.1-VP24的质粒图谱。C. VP24基因的测序结果。其中下划线分别代表酶切位点BamHⅠ和EcoRⅠ。方框标出起始密码子(ATG)和终止密码子(TAA)。D. 2种VP24基因的同源比对结果。VP24:PCR扩增片段。VP24-Ⅰ:WSSV印度株(GenBank No. DQ681068.1)。A. The agarose gel electrophoresis analysis of PCR products of VP24 gene. Lane M: Trans2K Plus DNA maker. Lanes 1 and 2: PCR products of VP24 gene. B. The plasmid map of pcDNA3.1-VP24. C. sequence analysis of VP24 gene. The endonuclease cleavage sites of BamHⅠ and EcoRⅠ are underlined. The initiation codon (ATG) and termination codon (TAA) are boxed. D. the alignment result of two nucleic acid sequences of VP24 gene. VP24, the sequence of amplified VP24 gene in this study. VP24-Ⅰ, the sequence of VP24 gene of the Indian strain (GenBank No. DQ681068.1).)
2.2 3种对虾病毒囊膜蛋白对假型昆虫杆状病毒包装效率的影响
如图2A1—F1所示,在最佳转染条件下,Bacmid-GUS重组表达质粒单独转染Sf9细胞的转染效率为(85.7±3.4)%,而Bacmid-GUS分别同3种对虾病毒囊膜蛋白表达质粒pcDNA3.1-VP19、pcDNA3.1-VP24和pcDNA3.1-VP28共转染Sf9细胞的转染效率分别为(90.7±0.3)%、(87.3±1)%和(85±1)%。由于GUS报告基因的表达效率同病毒的包装效率正相关,因此,上述结果提示,同非假型病毒Bacmid-GUS相比,上述3种假型病毒的包装效率没有得到明显提高。换言之,在最佳转染条件下对虾病毒3种囊膜蛋白(VP19、VP24和VP28)对其假型病毒(Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28)的包装效率没有明显的促进作用。

(A1—F1为最优转染条件下的转染结果,即Bacmid-GUS质粒为0.375 μg/孔。A2—F2为非最佳转染条件下的转染结果,即Bacmid-GUS质粒为0.15 μg/孔。A1和A2分别为不加Bacmid-GUS质粒的阴性对照。B1和B2为Bacmid-GUS单独转染结果。C1和C2为Bacmid-GUS与pcDNA3.1-VP19共转染结果。D1和D2为Bacmid-GUS与pcDNA3.1-VP24共转染结果。E1和E2为Bacmid-GUS与pcDNA3.1-VP28共转染结果。F1为B1—E1的转染效率的柱状图。F2为B2—E2的转染效率柱状图。转染效率为平均值(标准误(n=9)。表示差异显著,单因素方差分析,P<0.05。标尺,100 μm。Panels A1 to F1 are the results obtained under the optimal co-transfection condition, the amount of Bacmid-GUS plasmid used is 0.375 μg/well. Panels A2 to F2 are the results obtained under non-optimal transfection condition, the amount of Bacmid-GUS plasmid used is 0.15 μg/well. Panels A1 and A2, the un-transfected control Sf9 cells. Panels B1 and B2: The Sf9 cells transfected by Bacmid-GUS. Panels C1 and C2, the Sf9 cells co-transfected by Bacmid-GUS and pcDNA3.1-VP19. Panels D1 and D2: The Sf9 cells co-transfected by Bacmid-GUS and pcDNA3.1-VP24. Panels E1 and E2: The Sf9 cells co-transfected by Bacmid-GUS and pcDNA3.1-VP28. Panels F1 shows the corresponding packaging efficiencies of Panels B1 to E1. F2 shows the corresponding packaging efficiencies of panels B2 to E2. Data for transfection efficiency is expressed as average(standard error (n=9). represents significant difference, One-way ANOVA, P<0.05. Scale bar, 100 μm.)
考虑到在最优转染条件下GUS转染效率过高,可能会掩盖上述3种对虾病毒囊膜蛋白对假型病毒包装效率的促进作用。因此,本文又进一步比较了非最佳转染条件下GUS转染效率,即将Bacmid-GUS质粒的量降低为0.15 μg/孔。结果发现,如图2A2—F2所示,Bacmid-GUS的单独转染效率为(43.3±3.1)%,而Bacmid-GUS同pcDNA3.1-VP19、pcDNA3.1-VP24和pcDNA3.1-VP28的共转染效率分别为(57.4±6.2)%、(54.8±3.4)%和(55.0±4.0)%。可见,同Bacmid-GUS昆虫杆状病毒相比,上述3种假型病毒Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28的包装效率分别提高了14.1%、11.5%和11.7%,差异具有统计学意义(P<0.05)。这表明,对虾病毒3种囊膜蛋白(VP19、VP24和VP28)对3种假型病毒的包装效率具有明显的促进作用。但是其促进包装的具体机制尚不清楚。
2.3 过表达囊膜蛋白(VP19、VP24和VP28)在包装细胞Sf9中的亚细胞定位
如图3所示,将红色荧光蛋白(mCherry)或绿色荧光蛋白(eGFP)标记的VP19、VP24和VP28融合蛋白表达质粒(pcDNA3.1-VP19-△TAA/mCherry、pcDNA3.1-VP24-△TAA/eGFP和pcDNA3.1-VP28-△TAA/mCherry)分别同Bacmid-GUS质粒共转染至包装细胞Sf9中,均可在感染后细胞中检测到明亮的红色或绿色荧光信号,这表明本研究新构建的VP19-mCherry融合蛋白表达质粒(pcDNA3.1-VP19-△TAA/mCherry)和VP24-eGFP融合蛋白表达质粒(pcDNA3.1-VP24-△TAA/eGFP)以及实验室前期构建的VP28-mCherry融合蛋白表达质粒(pcDNA3.1-VP28-△TAA/mCherry)均是成功的。激光共聚焦显微镜观察结果表明,红色荧光信号(VP19和VP28)与绿色荧光信号(VP24)的分布呈明显的环状,位于细胞核之外。可见,3种融合表达囊膜蛋白主要定位于Sf9细胞的细胞膜上。
2.4 3种假型昆虫杆状病毒的滴度测定结果
将浓缩纯化后的Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28三种假型病毒分别感染Sf9细胞,然后X-gluc染色检测GUS基因的表达,结果如图4所示,上述3种假型病毒的单位体积(mL)滴度分别为1.3×1013、9.5×1010和1.2×1011个转导单位。

(A、B和C为未感染病毒的Sf9细胞(对照),A′、B′和C′分别为假型病毒Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28感染Sf9细胞的GUS表达结果。标尺:100 μm。Panels A, B and C, the uninfected Sf9 cells (control). Panels A′, B′ and C′, GUS expression results of the Sf9 cells infected by pseudotyped baculoviruses of Bacmid-GUS/VP19, Bacmid-GUS/VP24 and Bacmid-GUS/VP28, respectively. Scale bar: 100 μm.)
2.5 3种假型昆虫杆状病毒感染对虾成体组织的结果
将3种假型病毒(Bacmid-GUS/VP19、Bacmid-GUS/VP24和Bacmid-GUS/VP28)以不同的注射剂量感染成体对虾,5 d后分别取染毒对虾的心脏、Oka器官、肠、鳃和肌肉组织于1.5 mL EP管中,以进行X-gluc染色以检测GUS报告基因的表达,结果如图5—7所示。结果表明,携带不同囊膜蛋白的假型病毒在感染对虾成体时具有不同的最低感染剂量和不同的组织特异性。其中,心脏组织中有弱的GUS本底表达,而所检测的其他组织中则没有GUS本底表达。
假型病毒Bacmid-GUS/VP19的包装效率和病毒滴度是最高的,因而对单个对虾的最高染毒剂量可达到1×1011个转导单位。其感染结果如图5所示,该假型病毒只能感染对虾的心脏和肌肉组织,不能感染Oka器官、肠和鳃组织,具有明显的组织特异性。但是其单个对虾感染剂量在1×108个转导单位时,染毒对虾心脏组织的GUS阳性信号才明显高于对照组织;而单个对虾肌肉组织的感染剂量则需要达到1×1011个转导单位时才能检测到GUS信号。
假型病毒Bacmid-GUS/VP24的滴度是最低的,因而其对单个对虾的最高染毒剂量只能达到1×109个转导单位,感染结果如图6所示,该假型病毒与Bacmid-GUS/VP19相似,也是只能感染对虾的心脏和肌肉组织,不感染Oka器官、肠和鳃组织,具有明显的组织特异性。但是其在单个对虾心脏中的最低感染剂量为1×107个转导单位,在单个对虾肌肉组织中的最低感染剂量为1×108个转导单位。
假型病毒Bacmid-GUS/VP28的滴度介于前述两种假型病毒之间,其个体最高染毒剂量也只能达到1×109个转导单位,感染结果如图7所示,与前述VP19和VP24两种假型病毒的组织特异性不同,该假型病毒不仅能够感染对虾的心脏和肌肉组织,还可以感染鳃组织,但是不能感染淋巴器官和肠组织。而且其对单个对虾的最低感染剂量,在心脏组织中为1×107个转导单位,在鳃组织中为1×108个转导单位,在肌肉组织中为1×109个转导单位。

(同图6图注)
综上所述,上述假型病毒在成体对虾中具有不同的组织特异性和不同的最低感染剂量。
2.6 假型昆虫杆状病毒在对虾成体组织中的感染是复制缺陷的
上述3种假型病毒感染阳性的对虾成体组织的透射电镜分析结果如图8所示。结果表明,在染毒和未染毒对虾的肌肉、心脏和鳃组织中均没有观察到假型杆状病毒颗粒。仅在感染了假型病毒Bacmid-GUS/VP28的鳃组织细胞内,观察到有异常的圆形结构,直径大约为90 nm(见图8B3),以及感染假型病毒Bacmid-GUS/VP24的心脏组织细胞内观察到有不规则的线性结构,长度为100~400 nm(见图8C2)。这说明同哺乳动物细胞中的感染结果相似,假型昆虫杆状病毒在对虾成体组织中的感染也是复制缺陷的。也就是说,假型病毒可以感染对虾组织和细胞,甚至可以在对虾宿主细胞内表达病毒的蛋白和核酸,但是在异源细胞内失去了包装为成熟病毒粒子的能力。具体机制尚不清楚。

(A1—A3:未感染病毒对虾的肌肉、心脏和鳃组织的电镜分析结果。B1—B3:感染假型病毒Bacmid-GUS/VP28的对虾肌肉、心脏和鳃组织的电镜分析结果,B3′为B3图中方框的放大结果,B3′图中箭头指示异常圆形结构。C1和C2分别为感染假型病毒Bacmid-GUS/VP24的对虾肌肉和心脏组织的电镜分析结果,C2′为C2图中方框的放大结果,C2′图中箭头指示不规则线性结构。D1和D2分别为感染假型病毒Bacmid-GUS/VP19的对虾肌肉和心脏组织的电镜分析结果标尺,2 μm。Panels A1 to A3 were the results of muscle, heart and gill tissue of uninfected shrimps. Panels B1 to B3 were the results of muscle, heart and gill tissue of shrimps infected by the pseudotyped baculovirus of Bacmid-GUS/VP28, and B3′ was the enlargement of the box in Panel B3. The arrow in the Panel B3′ indicates an abnormal circular structure. Panels C1 and C2 were the results of muscle and heart tissue of shrimps infected by pseudotyped baculovirus of Bacmid-GUS/VP24, respectively. C2′ is the enlargement of the box in Panel C2. The arrow in Panel C2′ indicated an irregular linear structure. Panels D1 and D2 were the results of muscle and heart tissue of shrimps infected by pseudotyped baculovirus of Bacmid-GUS/VP19, respectively.)
3 讨论
囊膜蛋白是位于具囊膜病毒囊膜上的一类病毒蛋白,一般具有至少一个跨膜结构域,其在病毒侵染过程中主要发挥识别和结合宿主细胞的作用,与病毒侵染的宿主范围和组织与细胞特异性有关[2]。假型病毒是指囊膜中插入了另一种病毒的囊膜蛋白的病毒,外源囊膜蛋白的引入,往往导致假型病毒的亲嗜性发生改变。共转染是制备假型病毒的常用方法,即在病毒包装时,将病毒载体质粒、包装表达质粒和囊膜蛋白表达质粒共转染至包装细胞中,从而获得假型病毒[3]。其中载体质粒携带外源基因,包装质粒携带病毒复制和包装的遗传信息,表达质粒携带其他病毒的囊膜蛋白基因。因此,过表达的囊膜蛋白能否定位于包装细胞的细胞膜上是决定假型病毒制备能否成功的前提条件。Syed Mustha等[13]已报道,过表达的对虾WSSV病毒囊膜蛋白VP28可定位于Sf9细胞的细胞膜上,并用免疫实验证明了包装出的昆虫杆状病毒的囊膜上携带了VP28。本研究采用红色(mCherry)和绿色(eGFP)荧光蛋白报告基因分别标记对虾WSSV病毒的VP19和VP24两种囊膜蛋白,进一步分析了VP19和VP24在Sf9细胞中过表达后的亚细胞定位,发现这两种囊膜蛋白同VP28类似,过表达后均可定位于包装细胞Sf9的细胞膜上,表明昆虫杆状病毒从Sf9细胞中出芽和释放时可携带WSSV病毒囊膜蛋白,这为VP19-和VP24-假型昆虫杆状病毒的成功制备奠定了基础。
本研究中还发现,VP19、VP24和VP28的表达均可不同程度地提高假型病毒的包装效率,其中,以VP19的促包装效果最明显,但是其促进包装的具体机制尚不清楚。Chen等[5]采用类似的方法将VP19和VP28引入到慢病毒表达系统,也明显提高了假型慢病毒的包装效率,分别比非假型慢病毒高出52.9%和129.4%。可见,VP19和VP28在哺乳动物包装细胞(293T)中的促包装效果远高于其在昆虫细胞中的结果(14.1%和11.7%)。本文作者认为,这可能同VP19和VP28的表达质粒在不同包装细胞中的表达效率有关。上述囊膜蛋白表达质粒中采用的启动子为CMV启动子,该启动子在哺乳动物细胞内为强启动子,活性高,但是其在昆虫细胞中为弱启动子[14]。此外,还发现报告质粒pcDNA3.1-VP28-△TAA/mCherry单独转染Sf9细胞时的CMV启动子活性更低,红色荧光信号很弱;但是,将其与Bacmid-GUS共转染Sf9细胞时,红色荧光信号明显变强,但是转染效率仍远低于其在哺乳动物细胞(HeLa)中的转染效率(结果未发表)。可见,昆虫杆状病毒的表达产物可以激活CMV启动子的活性。
Wu等[6]已报道了VP28-假型病毒感染对虾活体的研究结果,发现VP28-假型病毒仅感染对虾的心脏、鳃和肠组织,不能感染Oka器官和肌肉组织,具有组织特异性。其在单个对虾的心脏和鳃组织中的最低感染剂量为1.5×107个转导单位,在单个对虾肠组织中最低感染剂量为3×107个转导单位。本文在VP28-假型病毒感染对虾成体的组织特异性及其最低感染剂量上的研究结果同Wu等[6]的结果不完全相同。首先,在病毒感染的组织特异性上,本研究中没有在肠组织中检测到阳性结果,这可能与肠道中有食物残渣,尤其是大肠杆菌有关。因为大肠杆菌可表达GUS蛋白,导致肠道出现假阳性结果。因此,在对虾成体染毒前,一定要将对虾进行饥饿处理,以降低假阳性出现的概率。另外,本文发现当病毒对单个对虾的感染剂量为1×109个转导单位时,可在肌肉组织中检测到GUS信号,即VP28-假型病毒在单个对虾肌肉组织中的最低感染剂量为1×109个转导单位,而Wu等[6]的工作中所检测的对单个对虾的最大染毒剂量为6×108个转导单位,从而导致其在肌肉组织中出现假阴性的检测结果。最后,本文在单个对虾鳃组织中的最低感染剂量为1×108个转导单位,高于Wu等[6]结果,这说明在更低的感染剂量下假型病毒也能感染对虾的鳃组织。
本文首次将VP19和VP24引入到昆虫杆状病毒表达系统(Bacmid-GUS)中,探究了其对假型昆虫杆状病毒在对虾活体组织中的亲嗜性的影响。发现与VP28-假型病毒不同,VP19-和VP24-假型病毒仅能感染对虾的心脏和肌肉组织。尤其是3种假型病毒在肌肉中的最低感染剂量差异明显,VP19、VP24和VP28的假型病毒在单个对虾肌肉中的最低感染剂量分别为1×1011、1×108和1×109个转导单位。这说明上述3种囊膜蛋白在对虾WSSV病毒感染肌肉组织时的贡献可能也是不同的。
已有研究表明,昆虫杆状病毒可以感染哺乳动物细胞并表达外源基因,但是其在哺乳动物细胞内不能进行增殖,是复制缺陷的[15]。因此,昆虫杆状病毒在异源的对虾组织中是否能够复制和增殖也是本文作者想了解的一个科学问题。本文通过透射电镜观察了上述3种假型病毒感染阳性的对虾心脏、肌肉和鳃组织,但未在感染组织中观察到成熟的杆状病毒粒子。仅在感染VP28-假型病毒的对虾鳃组织细胞内观察到异常的圆形结构,以及感染VP24-假型病毒的对虾心脏组织细胞内观察到不规则的线性结构。其中,圆形结构直径为90 nm左右,线性结构长度为100~400 nm,与成熟的昆虫杆状病毒大小(30~60 nm×250~300 nm)不符合。在对虾成体中能检测到上述假型昆虫杆状病毒含GUS基因的表达产物,但未观察到成熟的病毒粒子,这表明,与昆虫杆状病毒感染哺乳动物细胞的结果类似,昆虫杆状病毒在对虾细胞内可能也无法进行复制或包装,是复制缺陷的。
综上所述,本研究成果拓展了人们对于对虾WSSV病毒囊膜蛋白在假型昆虫杆状病毒的包装及其侵染对虾的组织特异性上的作用的认知。
猜你喜欢
山东畜牧兽医(2022年10期)2022-12-06 03:52:34
当代水产(2022年8期)2022-09-20 06:46:22
当代水产(2022年5期)2022-06-05 07:55:46
当代水产(2021年8期)2021-11-04 08:49:04
当代水产(2021年7期)2021-11-04 08:17:54
当代水产(2021年4期)2021-07-20 08:10:54
哈尔滨医药(2016年3期)2016-12-01 03:58:34
中国兽医杂志(2016年5期)2016-06-27 01:44:58
广东海洋大学学报(2015年4期)2016-01-13 08:39:33
癌变·畸变·突变(2015年4期)2015-02-27 06:15:17
