
乳酸诱导巨噬细胞M2型极化对黑色素瘤细胞增殖、凋亡、迁移和侵袭的影响
2023-10-17 01:59杨颖颖邱炜
安徽医药 2023年11期
杨颖颖,邱炜
作者单位:1贵州医科大学生物与工程学院,贵州 贵阳550000;2遵义市第一人民医院肿瘤科,贵州 遵义563000
恶性黑色素瘤是最具侵袭性和抗治疗性的皮肤癌形式,其侵袭性主要是基于黑色素瘤细胞的高度转移潜力[1]。尽管靶向治疗和免疫检查点抑制剂治疗取得了一定的疗效,但转移性黑色素瘤仍然是一种无法治愈的疾病[2-3]。因此,探索黑色素瘤转移的机制对于提高其诊断和治疗水平极为必要。
肿瘤转移与肿瘤微环境(tumor microenvironment,TME)高度相关[4]。缺氧引发一系列生化反应引发局部酸化进而将TME转变为对抗肿瘤免疫细胞不利环境的驱动力之一[5]。乳酸(lactic acid,LA)是一种信号分子,在调节TME内的代谢途径、免疫反应和细胞内通信等方面具有重要作用[6]。此外,乳酸在厌氧肿瘤环境中浓度很高,并且已被证明可以改变巨噬细胞以获得促进肿瘤生长的特性[7]。肿瘤相关巨噬细胞(tumor-associated macrophages,TAM)参与调控恶性肿瘤发生和进展的稳态。此外,已有多项研究表明,在多种类型黑色素瘤中都观察到少量巨噬细胞[8]。然而,TAM在黑色素瘤转移中的功能仍然未知。本研究于2021年1月至2022年2月,首次描述了黑色素瘤的恶性生物学行为与乳酸诱导的TAM 极化之间的关系。
1 材料和方法
1.1 主要材料 乳酸(纯度≥98%,L6402)、脂多糖(LPS)(来源于大肠杆菌0111:B4,L5293)、白细胞介素(IL)-4(纯度≥98%,SRP3211)获取自美国Sigma-Aldrich公司;小鼠巨噬细胞Raw264.7(HTX1760)和小鼠黑色素瘤细胞系B16F10(AC-2661H)获取自上海雅吉生物科技有限公司;CCK-8细胞增殖与毒性检测试剂盒(CK04)获取自上海研卉生物科技有限公司;Annexin V-FITC/PI凋亡试剂盒获取自台湾亚诺法生技股份有限公司;小鼠IL-12 ELISA试剂盒(orb566228)、小鼠IL-10 ELISA试剂盒(orb565120)获取自英国Biorbyt公司;CD206(ab270647)抗体获取自英国Abcam公司;High Capacity cDNA反转录试剂盒(4368813)、Maxima SYBR Green qPCR(K0253)获取自美国Thermofisher。
1.2 方法
1.2.1 实验药物配制 乳酸通过首先溶解在少量的DMSO溶液中,而后慢慢添加PBS缓冲液溶解进而配制形成1 mol/L的母液,保存于-4 ℃冰箱备用。待使用时使用相应的培养基配制为合适的浓度。
1.2.2 细胞培养 Raw264.7和B16F10细胞使用DEME培养基培养(含10%胎牛血清),常规传代培养。实验中使用的细胞传代次数不超过30次。
1.2.3 乳酸的细胞毒性检测 将培养的Raw264.7和B16F10细胞以1×104个/孔铺至96孔板中继续培养,待细胞贴壁生长后,添加不同浓度的乳酸(0、5、10、20 mmol/L)[9]继续培养24 h。随后,添加10 μL的CCK-8溶液培养4 h后,在酶标仪中检测乳酸处理后的细胞活力。细胞活力(%)=(OD对照组-OD实验组)/(OD对照组-OD空白组)×100%。
1.2.4 细胞分组 将Raw264.7细胞以1×106个/孔铺至6孔板中,将其依次分为对照组、LPS组(诱导M1型巨噬细胞)、IL-4组(诱导M2型巨噬细胞)、乳酸 5 mmol/L组、乳酸 10 mmol/L组和乳酸 20 mmol/L组。对照组Raw264.7细胞常规培养,LPS组Raw264.7细胞使用终浓度为1 mg/L的LPS[10]处理,IL-4组Raw264.7细胞使用终浓度为20 μg/L的IL-4[10]处理;乳酸 5 mmol/L组、乳酸 10 mmol/L组和乳酸 20 mmol/L组分别使用终浓度为5、10、20 mmol/L的乳酸处理,所有细胞均培养24 h。
1.2.5 黑色素瘤细胞增殖实验 收集各组处理的Raw264.7细胞的上清液用于培养B16F10细胞培养24 h。随后,添加10 μL的CCK-8溶液培养4 h后,在酶标仪中检测乳酸处理后的细胞活力。细胞活力(%)=(OD对照组-OD实验组)/(OD对照组-OD空白组)×100%。
1.2.6 黑色素瘤细胞凋亡实验 各组处理的Raw264.7细胞的上清液培养B16F10细胞24 h后,将细胞悬浮于200 μL的缓冲液中,分别添加10 μL的Annexin V-FITC和5 μL的PI溶液避光孵育20 min,在1 h内通过流式细胞仪分析各组细胞凋亡率。
1.2.7 黑色素瘤细胞迁移实验 B16F10细胞按照1×106个/孔铺至6孔板中,待其铺满后,使用200 μL移液器枪头进行划痕,使用各组处理的Raw264.7细胞的上清液培养24 h,分别对0 h和24 h后的划痕宽度进行拍照记录。划痕愈合率(%)=(0 h划痕宽度-24 h划痕宽度)/0 h划痕宽度×100%。
1.2.8 黑色素瘤细胞侵袭实验 50 μL的基质胶包被在Transwell小室(8 μm)上室中,将B16F10细胞使用无血清的DEME培养基稀释后接种在Transwell小室上室中,下室中添加正常培养基。常规培养24 h后,使用4%多聚甲醛将小室下室中细胞固定并通过0.1%结晶紫染色20 min,在显微镜下对侵袭到下室中的细胞进行拍照并统计。
1.2.9 巨噬细胞形态观测 培养Raw264.7细胞,按照“1.2.4”中方法分组处理后于光学显微镜下观察各组Raw264.7细胞形态学变化。
1.2.10 巨噬细胞极化相关因子表达检测 严格按照试剂盒说明,收集每组Raw264.7细胞上清液,每孔中添加50 μL的样品,使用试剂盒中的胶带覆盖并孵育。加入对应的IL-12或IL-10结合物,再次孵育。加入底物避光孵育30 min后加入终止液终止反应。检测各组细胞的吸光度,根据绘制的标准曲线得出各因子浓度。
1.2.11 qRT-PCR检测肿瘤蛋白翻译调节因子1(Tpt1)和肺腺癌转移相关转录本1(Malat1)表达水平 Trizol试剂从各组处理的Raw264.7细胞中分离总RNA。使用反转录试剂盒制备cDNA。接下来使用SYBR Green(2×)和特异性引物(均由上海生工设计并合成)进行qRT-PCR分析,使用2-ΔΔCt方法计算倍数变化,然后进行统计分析。引物序列见表1。

表1 引物序列
1.2.12 CD206表达水平检测 Raw264.7细胞接种在6孔板中(1×106个/孔),按照1.2.4中方法分组处理后收集细胞,加入CD206抗体在室温下避光孵育20 min,添加PBS清洗后,在流式细胞仪中检测CD206表达水平。
1.3 统计学方法 SPSS 22.0对本文中的实验数据进行分析。Shapiro-Wilk检验进行正态性检测,符合正态分布的研究结果使用来表示。采用单因素方差分析进行多组间的比较,多组间比较具有差异的数据进一步两两比较通过SNK-q检验进行。P<0.05为差异有统计学意义。
2 结果
2.1 乳酸对巨噬细胞和和黑色素瘤细胞活性的影响 不同浓度乳酸对Raw264.7细胞和B16F10细胞均无明显细胞毒性(P>0.05)。见表2。
表2 乳酸对Raw264.7细胞和B16F10细胞的毒性作用检测/
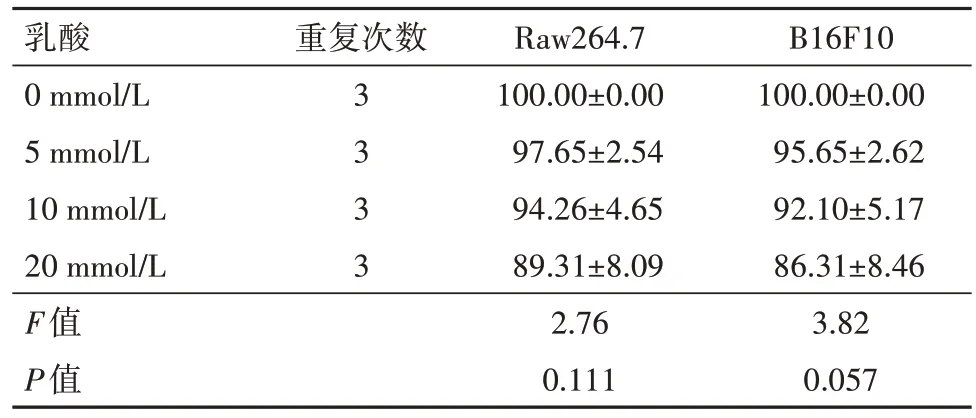
表2 乳酸对Raw264.7细胞和B16F10细胞的毒性作用检测/
乳酸0 mmol/L 5 mmol/L 10 mmol/L 20 mmol/L F值P值重复次数3 3 3 3 Raw264.7 100.00±0.00 97.65±2.54 94.26±4.65 89.31±8.09 2.76 0.111 B16F10 100.00±0.00 95.65±2.62 92.10±5.17 86.31±8.46 3.82 0.057
2.2 乳酸调控巨噬细胞对黑色素瘤细胞恶性生物学行为的影响 如表3结果所示,使用不同浓度乳酸干预Raw264.7细胞后的上清液培养B16F10细胞,结果显示,与对照组相比,乳酸组和IL-4组细胞活力、划痕愈合率以及侵袭细胞数量均明显升高,细胞凋亡率显著降低(P<0.05),LPS组细胞活力、划痕愈合率、侵袭细胞数量以及凋亡率均无显著变化(P>0.05)。而不同浓度乳酸组随着乳酸浓度升高,细胞活力、划痕愈合率以及侵袭细胞数量逐渐升高,细胞凋亡率逐渐降低(P<0.05)。由此推测,乳酸可能诱导巨噬细胞M2型极化促进黑色素瘤恶性生物学行为。
表3 乳酸对B16F10细胞恶性生物学行为的影响/
注:LPS为脂多糖,IL为白细胞介素。①与对照组比较,P<0.05。②与LPS组比较,P<0.05。③与IL-4组比较,P<0.05。④与乳酸5 mmol/L组比较,P<0.05。⑤与乳酸10 mmol/L组比较,P<0.05。
侵袭细胞数/个45.36±4.21 50.13±5.23 167.26±12.36①②89.36±8.12①②③113.05±10.03①②③④131.25±11.14①②③④⑤126.32<0.001组别对照组LPS IL-4乳酸 5 mmol/L乳酸 10 mmol/L乳酸 20 mmol/L F值P值重复次数3 3 3 3 3 3细胞活力/%100.00±0.00 92.23±8.76 188.37±9.25①②125.36±7.88①②③144.36±8.65①②③④167.28±10.12①②③④⑤63.78<0.001凋亡率/%57.26±4.25 55.14±3.45 8.20±0.87①②41.23±4.02①②③29.26±3.14①②③④18.64±2.09①②③④⑤115.39<0.001划痕愈合率/%25.26±2.23 28.23±2.02 78.26±6.25①②39.21±3.15①②③52.65±3.87①②③④64.24±6.12①②③④⑤71.25<0.001
2.3 乳酸诱导巨噬细胞Raw264.7向M2型极化设置LPS诱导的M1型巨噬细胞和IL-4诱导的M2型巨噬细胞为对照,使用上述验证的无毒性的乳酸处理巨噬细胞24 h。观察细胞形态得出,乳酸组巨噬细胞形态学变化与IL-4组相似,均呈现M2样变化。进一步结果显示(表4),与对照组比较,LPS组细胞中肿瘤坏死因子-α(TNF-α)、IL-12均升高(P<0.05),血管内皮生长因子(VEGF)、IL-10、CD163水平无显著变化(P>0.05);IL-4组和乳酸 5 mmol/L组、乳酸10 mmol/L组和乳酸 20 mmol/L组细胞中TNF-α、IL-12水平差异无统计学意义,VEGF、IL-10、CD163水平显著升高(P<0.05)。ELISA实验检测的IL-12与IL-10表达水平以及流式细胞仪检测的CD206的表达水平则具有相似的结果(P<0.05)。经过对巨噬细胞的形态学观察和相关分子水平的检测得出,乳酸可诱导巨噬细胞向M2型极化。
表4 qRT-PCR检测乳酸对B16F10细胞M1和M2极化相关分子水平比较/

表4 qRT-PCR检测乳酸对B16F10细胞M1和M2极化相关分子水平比较/
注:LPS为脂多糖,IL为白细胞介素,TNF-α为肿瘤坏死因子-α,VEGF为血管内皮生长因子。①与对照组比较,P<0.05。②与LPS组比较,P<0.05。③与IL-4组比较,P<0.05。④与乳酸 5 mmol/L组比较,P<0.05。⑤与乳酸 10 mmol/L组比较,P<0.05。
CD163/%8.15±0.78 8.87±0.64 58.36±5.12①②18.39±1.54①②③25.69±2.87①②③④37.16±3.15①②③④⑤138.81<0.001组别对照组LPS IL-4乳酸 5 mmol/L乳酸 10 mmol/L乳酸 20 mmol/L F值P值重复次数3 3 3 3 3 3 TNF-α/(ng/L)22.39±3.36 45.63±4.58①23.15±1.37 22.36±2.15 24.36±2.02 20.19±2.18 34.45<0.001 IL-12/(ng/L)45.17±4.16 87.29±7.15①40.13±5.01 41.39±4.27 44.26±4.13 48.26±3.15 41.89<0.001 VEGF/(ng/L)78.15±7.56 82.56±8.36 245.36±20.15①②124.36±11.03①②③164.39±14.35①②③④198.65±15.39①②③④⑤72.89<0.001 IL-10/(ng/L)27.25±2.21 32.36±3.14 97.54±7.25①②47.26±4.02①②③65.32±5.48①②③④79.36±7.12①②③④⑤83.13<0.001
2.4 乳酸对巨噬细胞中Tpt1、Malat1表达的影响表5结果显示,与对照组比较,不同浓度乳酸可明显提高Tpt1 mRNA和Malat1 mRNA表达水平(P<0.05),结果与IL-4组相一致,而LPS组Tpt1 mRNA和Malat1 mRNA表达水平无明显变化。
表5 乳酸对Raw264.7细胞中Tpt1、Malat1表达水平的影响/
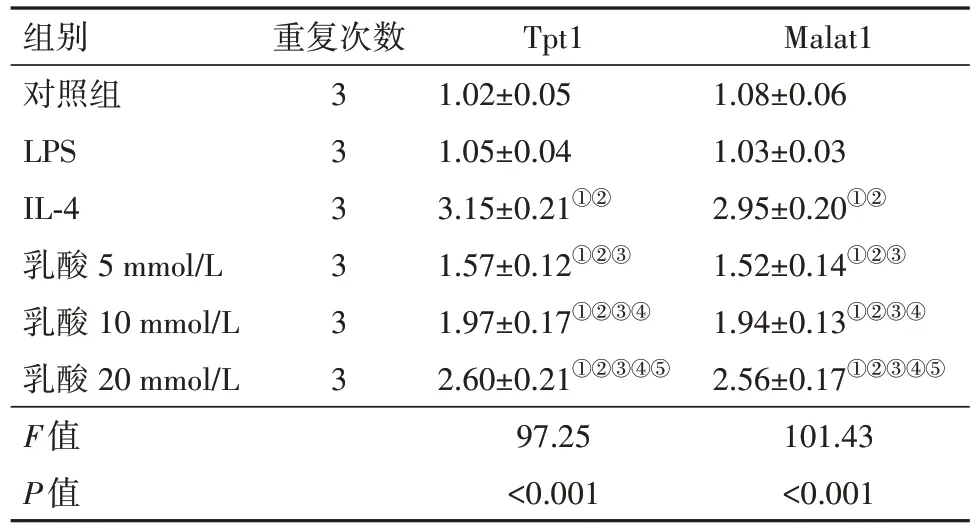
表5 乳酸对Raw264.7细胞中Tpt1、Malat1表达水平的影响/
注:LPS为脂多糖,IL-4为白细胞介素-4,Tpt1为肿瘤蛋白翻译调节因子1,Malat1为肺腺癌转移相关转录本1。①与对照组比较,P<0.05。②与LPS组比较,P<0.05。③与IL-4组比较,P<0.05。④与乳酸 5 mmol/L组比较,P<0.05。⑤与乳酸 10 mmol/L组比较,P<0.05。
Malat1 1.08±0.06 1.03±0.03 2.95±0.20①②1.52±0.14①②③1.94±0.13①②③④2.56±0.17①②③④⑤101.43<0.001组别对照组LPS IL-4乳酸 5 mmol/L乳酸 10 mmol/L乳酸 20 mmol/L F值P值重复次数3 3 3 3 3 3 Tpt1 1.02±0.05 1.05±0.04 3.15±0.21①②1.57±0.12①②③1.97±0.17①②③④2.60±0.21①②③④⑤97.25<0.001
3 讨论
以往研究大多都集中在基因组本身与肿瘤本身,以说明肿瘤恶性行为的机制。肿瘤细胞与基质细胞之间的相互调节塑造了TME的状态,这可能决定肿瘤的结果,并且可能是肿瘤发展的一种新机制[11]。乳酸已被证明可以改变多个关键癌基因的转录活性,并且研究证实乳酸产生诱发的致癌信号可以作为Warburg效应的一个重要目的[12]。乳酸代谢不仅与缺氧(产生乳酸)和常氧(输入乳酸)癌细胞之间的代谢平衡相关,而且还与缺氧癌细胞将TAM极化为糖酵解不良的M2型巨噬细胞有关[13]。据报道,癌细胞中糖酵解增加引起的而TME的酸化,而黑色素瘤则通过感知TME的酸化来促进TAM的M2型极化[14]。已证明,乳酸特别地将TAM偏向于“肿瘤友好”的M2样表型进而帮助肿瘤细胞逃避免疫监视[15]。本研究发现,乳酸对巨噬细胞Raw264.7和黑色素瘤细胞B16F10均无明显毒性,但乳酸作用Raw264.7细胞后的上清液可明显促进B16F10细胞的恶性生物学行为。提示乳酸可能通过调控TAM,进而对黑色素瘤具有促癌作用。
TAM是组织和TME内稳态的关键调节剂。原发性肿瘤中的TAM促进肿瘤的迁移和侵袭,被认为是癌症联合治疗中极具吸引力的一部分[16]。既往研究表明,肿瘤组织中巨噬细胞常为M2型,往往具有促进肿瘤增殖和转移的作用[17]。另外,研究证实,在小鼠模型体内调节巨噬细胞向M1型激活可有效抑制黑色素瘤细胞转移[18]。本研究结果发现,乳酸干预Raw264.7细胞后,Raw264.7细胞呈现M2型变化,且可明显提高M2型巨噬细胞极化相关分子VEGF、CD163和IL-10的表达,而对M1型巨噬细胞极化相关分子IL-12和TNF-α的表达无显著影响。以上数据表明,乳酸可促进巨噬细胞M2型极化。然而,对于乳酸诱导巨噬细胞M2型极化后如何调控黑色素瘤的发展尚不清楚。因此,有必要对巨噬细胞M2型极化的关键分子靶点进行探究。Tpt1是一种高度保守的蛋白质,据报道它在多种恶性肿瘤中强烈表达,参与调控细胞增殖、侵袭、细胞周期和细胞凋亡等过程[19]。Wang等[20]研究发现,Tpt1在胃癌组织和细胞中高表达,抑制Tpt1可显著抑制胃癌细胞增殖和迁移。另外,Malat1是长链非编码RNA中的一种,首次在肺癌中发现,在多种癌症中均具有预后和诊断价值[21]。已有多项研究表明,Malat1在黑色素瘤中发挥促癌作用,下调Malat1可通过抑制肿瘤细胞的增殖、迁移和侵袭进而抑制黑色素瘤的发展[22]。且研究表明,细胞外囊泡穿梭的Malat1可促进巨噬细胞极化[23]。令人感兴趣的是,本研究发现,乳酸作用Raw264.7细胞可明显提高Tpt1和Malat1的表达水平,提示Tpt1和Malat1极有可能是乳酸诱导巨噬细胞M2极化的分子靶点,暗示乳酸可能通过上调Tpt1和Malat1表达进而诱导巨噬细胞M2极化,在黑色素瘤中发挥促癌作用。然而,其中的具体作用机制仍需进一步探究。
综上,乳酸诱导的TAM M2型极化可促进黑色素瘤的发展。这一发现为黑色素瘤的免疫治疗提供新的参考依据。然而,本研究并未对乳酸是否以及如何通过TAM极化影响黑色素瘤细胞活性进行初步验证,而是直接进行了TAM M2型极化分析,缺乏中间的过度实验。其次,本研究并未在动物模型中进行同步验证。后续仍需进一步探索以补充完善乳酸在黑色素瘤发展过程中作用机制。
猜你喜欢
现代财经-天津财经大学学报(2022年5期)2022-06-01
皮肤病与性病(2021年3期)2021-07-30
学生天地·小学低年级版(2017年12期)2018-04-16
电子测试(2017年15期)2017-12-18
西南国防医药(2016年6期)2016-12-01
初中生学习·高(2016年10期)2016-05-30
癌症进展(2016年10期)2016-03-20
实用手外科杂志(2015年4期)2015-08-27
电源技术(2015年1期)2015-08-22
物理实验(2015年10期)2015-02-28
