
褐飞虱对环氧虫啶的抗性选育和遗传方式
2023-10-16 03:38:28杜祖仪宋璐丹马康生李建洪
植物保护 2023年5期
覃 耀, 杜祖仪, 宋璐丹, 马康生, 万 虎, 李建洪
(华中农业大学植物科学技术学院,昆虫资源利用与害虫可持续治理湖北省重点实验室,武汉 430070)
褐飞虱Nilaparvatalugens(Stål)(brown planthopper,BPH)属半翅目Hemiptera飞虱科Delphacidae,是当前水稻上防控压力最大的害虫之一[1],亦是全球范围内抗药性事件排名第一的粮食作物害虫[2],因其防控意义重大而被农业农村部列入《一类农作物病虫害名录》[3]。目前,褐飞虱田间种群防控主要依赖化学防治,但随着用药历史不断延长,褐飞虱田间种群已经对众多常用杀虫剂产生了明显的抗性[4-7]。新烟碱类杀虫剂因其独特的作用机制及优异的杀虫活性被广泛应用于褐飞虱田间防治[8]。尽管我国已经停止使用吡虫啉防治稻田褐飞虱,但近年来,褐飞虱田间种群对吡虫啉仍具有极高水平抗性,对噻虫嗪和呋虫胺的抗性已达高水平,对噻虫胺的抗性处于中等至高水平抗性,对氟啶虫胺腈、烯啶虫胺和啶虫脒产生了中等水平抗性,并且抗药性水平有进一步上升的趋势[6, 9-10]。褐飞虱对杀虫剂抗性的持续升高可能会导致防治失败和水稻生产成本的大幅度增加[11],给新型杀虫剂的创制及害虫抗药性治理带来严峻的挑战。
环氧虫啶(cycloxaprid)是由华东理工大学自主研发的一种新烟碱类杀虫剂,具有独特的顺式硝基构型和七元氧桥杂环结构,而吡虫啉、呋虫胺和噻虫胺等传统新烟碱类杀虫剂中的-NO2或者-CN均为反式结构[12]。作为烟碱型乙酰胆碱受体(nAChR)拮抗剂,环氧虫啶对稻飞虱、蚜虫和粉虱等刺吸式口器害虫具有优异的杀虫活性[12-14],不易与吡虫啉等新烟碱类杀虫剂产生交互抗性,能够有效防治对吡虫啉具有抗性的褐飞虱[15-16]、棉蚜Aphisgossypii[17]和烟粉虱Bemisiatabaci[14]。目前,我国褐飞虱田间种群对环氧虫啶处于敏感至低水平抗性阶段[18]。
抗药性的发展受多种因素的影响,其中遗传因子影响抗性发展速度,研究害虫抗药性遗传方式有助于了解抗性动态及抗性治理策略的选择,也有助于抗性机理的研究。目前,关于昆虫和螨类抗性遗传方式的研究已有大量报道,例如,家蝇Muscadomestica对残杀威的抗性为单基因控制的常染色体、不完全隐性遗传[19];棉籽尖长蝽Oxycarenushyalinipennis对氟啶虫胺腈的抗性为常染色体、多基因控制的不完全显性遗传[20];小菜蛾Plutellaxylostella对氰氟虫腙、朱砂叶螨Tetranychuscinnabarinus对丁氟螨酯的抗性均为多基因控制的常染色体、不完全隐性遗传[21-22]。已有研究表明,通常多基因控制的抗性比单基因控制的抗性发展更缓慢,但是多基因控制的抗性形成后治理难度较大;隐性或不完全隐性基因控制的抗性有利于药剂防治和抗性治理[23-24]。目前褐飞虱对许多常用杀虫剂的抗性遗传方式已被广泛报道,但是褐飞虱对环氧虫啶的抗性遗传方式却鲜有报道。因此,明确褐飞虱对环氧虫啶的抗性遗传方式对于保护和延长环氧虫啶的使用寿命具有非常重要的意义,可为褐飞虱的高质量防控和抗性治理提供理论依据。
1 材料与方法
1.1 供试昆虫
褐飞虱室内敏感品系(CYC-S):2009年采自湖南省长沙市水稻田,在实验室连续饲养多年且未接触任何药剂;褐飞虱环氧虫啶室内抗性品系(CYC-R):由CYC-S品系经环氧虫啶连续34代的室内抗性筛选后获得,两个品系具有相同的遗传背景。
试验用褐飞虱均在人工气候培养箱中,采用未经过任何药剂处理的‘Taichung Native 1’(‘TN1’)水稻稻苗饲养,饲养条件为:温度(27±1)℃,相对湿度70%~80%,光周期L∥D=16 h∥8 h。
1.2 供试药剂
97.5%环氧虫啶(cycloxaprid)原药由华东理工大学馈赠。
1.3 试验方法
1.3.1生物活性测定
参照《水稻褐飞虱抗药性监测技术规程》(NY/T 1708-2009)中的稻苗浸渍法进行褐飞虱毒力测定。选取生长健康、长势一致的10 cm高稻苗,15根一组,洗净,于荫凉处晾干。将供试药剂的母液用0.1% Triton X-100水溶液稀释成6~9个系列浓度。将供试稻苗置于药液中浸30 s后取出晾干,并用浸水的脱脂棉包裹住稻苗根部后放入一次性塑料杯中,每杯接入15头健康一致的褐飞虱3龄中期若虫后用尼龙细纱布封口,以0.1% Triton X-100水溶液处理作为空白对照,每处理3个重复。将处理后的稻苗放入温度(27±1)℃,相对湿度70%~80%,光周期为L∥D=16 h∥8 h的人工气候培养箱中。处理后96 h统计各处理的若虫死亡数,以小毛笔轻触虫体,不能协调运动的个体视为死亡。利用POLO-Plus软件处理数据计算毒力回归方程的斜率(slope)、标准误、卡方值(χ2)、自由度(df)、LC50及其95%置信限。抗性倍数(resistance ratio, RR)=抗性品系褐飞虱的LC50/敏感种品系褐飞虱的LC50。抗性水平分级标准:RR ≤ 5为敏感;5
1.3.2抗性品系选育
由于褐飞虱田间种群的用药背景较为复杂,不宜作为抗性遗传方式研究的材料,故采用室内选育的抗环氧虫啶褐飞虱品系作为试验材料。将室内敏感品系(CYC-S)作为抗性选育的出发种群,一部分试虫进行抗性筛选,获得抗环氧虫啶褐飞虱品系(CYC-R),另一部分试虫(CYC-S)不接触任何药剂,于相同条件下继续饲养。在抗性筛选过程中,每一代都采用稻苗浸渍法进行毒力测定。根据上一代的毒力测定结果,选用环氧虫啶LC70浓度浸泡新鲜稻苗(每30根一组)30 s后取出晾干,以浸水的脱脂棉包住稻苗根部后放入一次性塑料杯中,每杯接入30头生长一致的3龄中期若虫后用尼龙纱布封口,放入人工气候培养箱中。每代筛选约3 000头试虫,处理后96 h,将存活试虫转移至未接触药剂的新鲜水稻苗上继续饲养、繁殖,供下一代筛选。
1.3.3杂交、回交试验
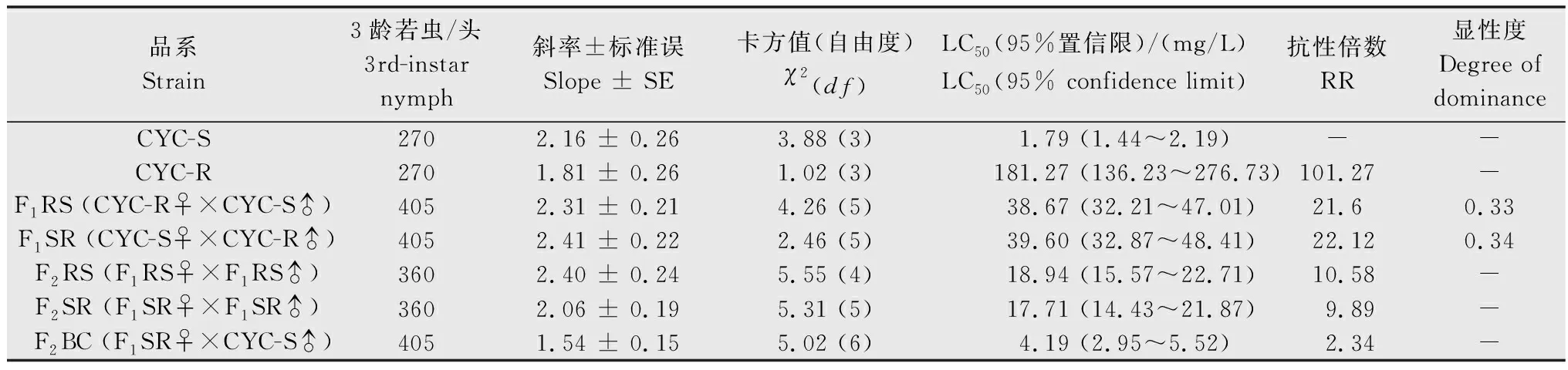
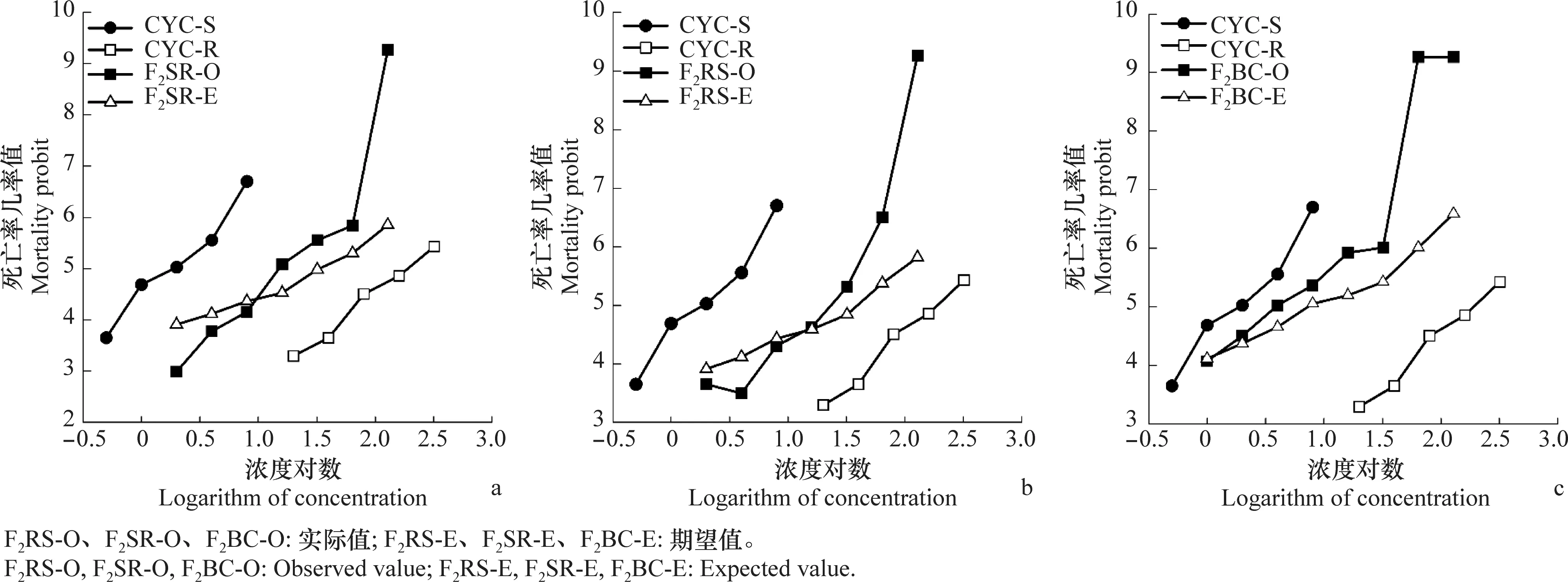
分别选取120头CYC-S和CYC-R品系5龄若虫,于玻璃试管中单头饲养。待若虫发育为成虫后,抗、敏品系进行正反交(CYC-R♀ × CYC-S♂;CYC-S♀ × CYC-R♂),后代为F1代(F1RS;F1SR)。在抗性不存在性连锁遗传的情况下,F1代与敏感父本CYC-S回交,后代为F2BC。F1代自交产生F2代(F2RS;F2SR)。每个杂交组合确保雌雄比例1∶1,雌雄虫各50头。
采用稻苗浸渍法分别测定环氧虫啶对亲本(CYC-R和CYC-S)、F1RS、F1SR、F2RS、F2SR、F2BC代褐飞虱3龄若虫的毒力,并绘制浓度对数-死亡率几率值线(LC-P线)。
1.3.4抗性遗传方式分析
根据Stone[25]的Falconer公式计算抗性显性度(D):
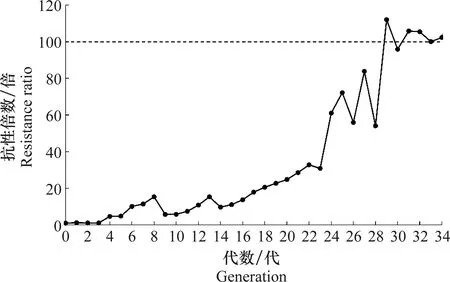
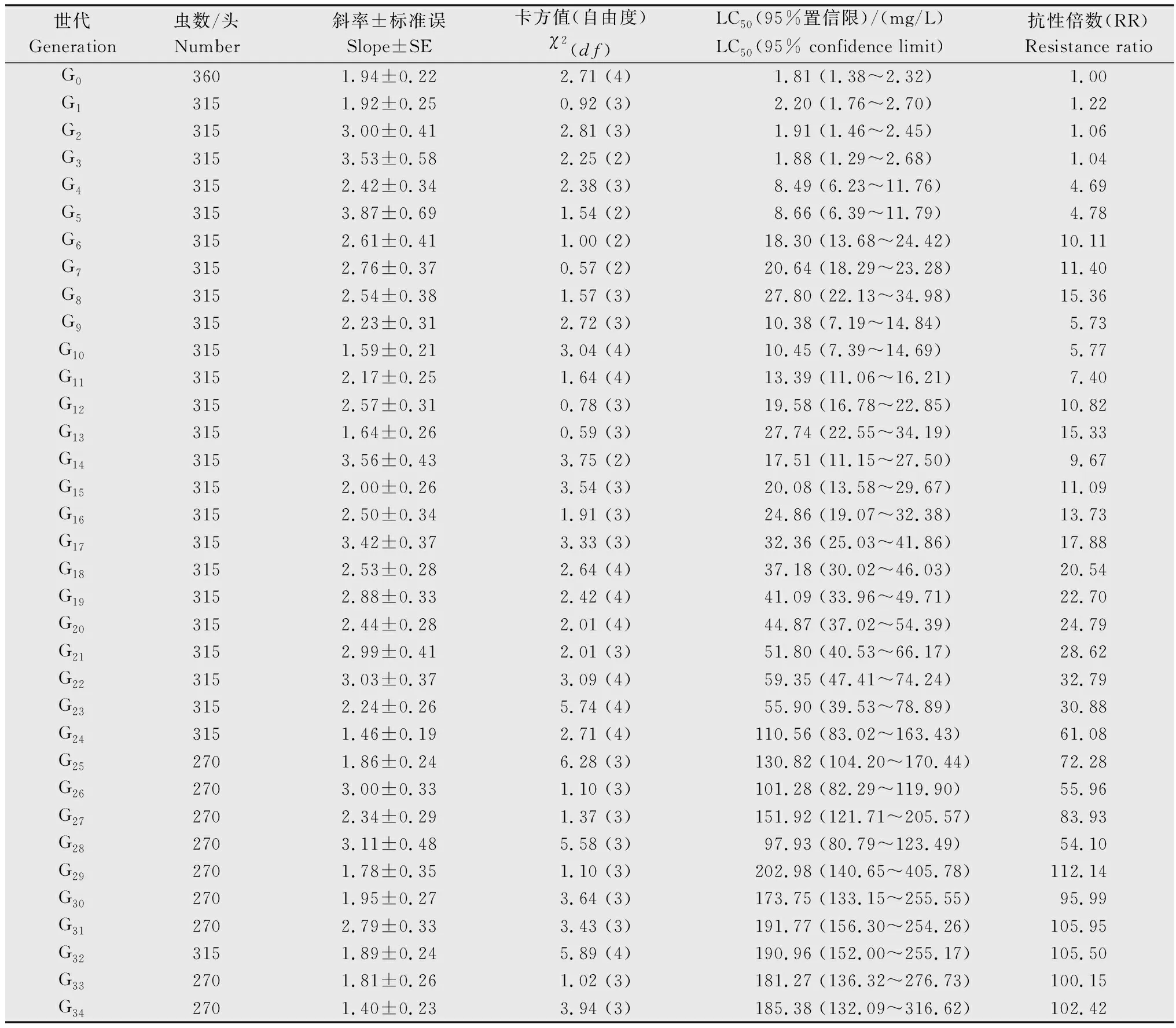
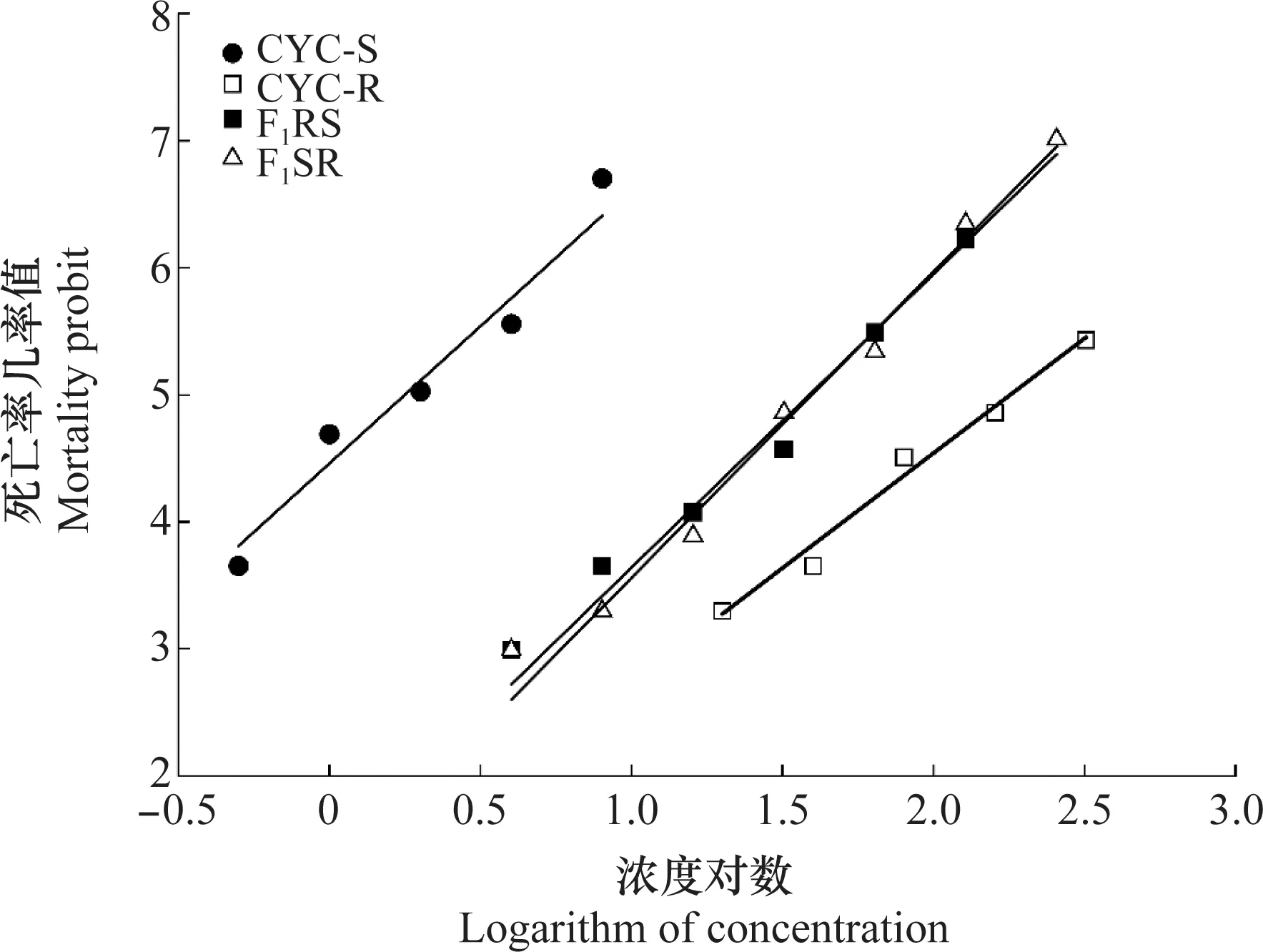
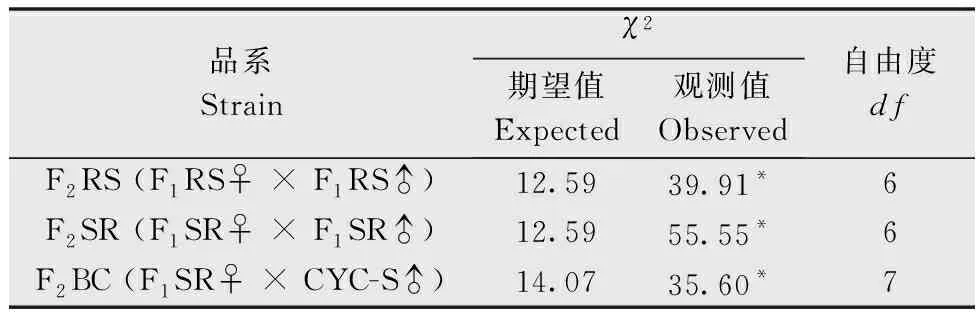
公式中X1、X2、X3分别为亲本抗性品系(CYC-R)、正反交F1(F1RS和F1SR)和亲本敏感品系(CYC-S)的LC50的对数值。当D=-1时,抗性为完全隐性遗传;当-1 根据Tsukamoto[26]提出的抗性遗传方式分析方法,假设褐飞虱对环氧虫啶的抗性遗传方式为单基因控制的遗传,则回交后代F2BC的浓度对数-死亡率几率值线在死亡率50%处、自交后代F2RS和F2SR在死亡率为25%和75%处会出现明显平坡。若以上相应位置没有出现明显平坡,表明褐飞虱对环氧虫啶的抗性由2个或2个以上基因控制。根据Georghiou等[27]提出的公式计算单因子遗传假定的自交后代F2RS、F2SR和回交后代F2BC在一系列药剂浓度下的期望死亡率: E(F2RS或F2SR)=W1× 0.25 +W2× 0.5 +W3× 0.25; E(F2BC)=(W1或W3)× 0.5+W2× 0.5; 其中E(F2RS)、E(F2SR)和E(F2BC)分别代表F2RS、F2SR和F2BC在相应浓度下的期望死亡率;W1、W2、W3分别为CYC-S、F1RS(或F1SR)、CYC-R在相应浓度下的实际死亡率。 为进一步确定控制抗性基因的数目,根据Sokal等[28]提出的单基因遗传假设的适合性测验,对F2RS、F2SR和F2BC的实测值和理论值进行适合性检验,公式如下: 采用稻苗浸渍法以褐飞虱敏感品系为出发种群进行室内抗性选育,以获得对环氧虫啶高抗的褐飞虱品系。通过连续34代的药剂筛选,获得了一个抗性倍数为102.42倍的抗性品系(CYC-R)。如表1和图1所示,前5代抗性上升较慢,LC50从2.20 mg/L上升为8.66 mg/L,处于敏感至低水平抗性阶段;随后G6至G23代抗性水平逐渐上升,抗性倍数从10.11倍上升为30.88倍,处于中等水平抗性;从G23代到G29代抗性上升较快,在G29代达到高水平抗性(RR=112.14倍),在此过程中,抗性水平存在较大的波动,G28代抗性急剧下降,但仍处于中等水平抗性,之后在G29代快速上升至高抗水平;第29代后抗性维持在高抗水平。 图1 褐飞虱对环氧虫啶抗性室内选育动态Fig.1 Dynamics of cycloxaprid-resistance in Nilaparvata lugens during resistance selection 表1 褐飞虱对环氧虫啶抗性选育1)Table 1 Selection for cycloxaprid-resistance in Nilaparvata lugens in laboratory 2.2.1抗性显性度分析 敏感品系CYC-S与抗环氧虫啶褐飞虱品系CYC-R经正反交后,得到F1RS和F1SR两个子代,由表2可知,环氧虫啶对F1RS和F1SR的LC50分别为38.67(32.21~47.01)mg/L和39.60(32.87~48.41)mg/L,二者数值较为接近,且95%置信区间基本重叠,即环氧虫啶对F1RS和F1SR的LC50无显著差异,表明褐飞虱对环氧虫啶的抗性基因位于常染色体上。通过Stone公式计算两个F1代的抗性显性度,显性度D值分别为0.33和0.34,二者均大于0小于1,表明褐飞虱对环氧虫啶的抗性为不完全显性遗传。为进一步验证上述结果,根据4个种群的毒力测定结果绘制了浓度对数-死亡率几率值线(LC-P线),由图2可知,杂交后代F1RS和F1SR的LC-P线处于两个亲本的中间位置,基本重叠,且更靠近抗性品系的LC-P线,这也进一步证实了褐飞虱对环氧虫啶的抗性为不完全显性遗传。 图2 环氧虫啶对CYC-S、CYC-R、正反交F1RS和F1SR代褐飞虱的毒力回归线Fig.2 Toxicity regression lines of cycloxaprid for CYC-S, CYC-R, and their reciprocal cross offspring of F1RS (CYC-R♀ × CYC-S♂) and F1SR (CYC-S♀ × CYC-R♂) 表2 环氧虫啶对抗性品系、敏感品系、杂交F1代、自交及回交F2代褐飞虱3龄若虫的毒力1)Table 2 Responses of parental, reciprocal crosses and backcross populations of Nilaparvata lugens to cycloxaprid 2.2.2遗传基因控制数量分析 图3 环氧虫啶对CYC-S、CYC-R、F2SR(a)、F2RS(b)和F2BC(c)代褐飞虱的浓度对数-死亡率几率值线Fig.3 Logarithm concentration-mortality probit lines of cycloxaprid for susceptible CYC-S, resistant CYC-R, F2SR (a), F2RS (b) and F2BC (c) 表3 褐飞虱对环氧虫啶抗性单基因遗传适合性检验1)Table 3 Chi-square (χ2) analysis for monogenic or polygenic inheritance of cycloxaprid-resistance in Nilaparvata lugens 近年来,褐飞虱田间种群已经对吡虫啉、烯啶虫胺、氟啶虫胺腈、噻虫胺、毒死蜱、吡蚜酮和噻嗪酮等多种杀虫剂产生了不同程度的抗性[4, 6-7],抗性形势日益严峻,但关于褐飞虱对环氧虫啶的抗性鲜有报道。在靶标害虫还未对新型杀虫剂产生抗性前,室内抗性选育及抗性遗传方式研究可为该药剂的合理使用和高质量抗性治理策略的制定及实施提供重要的理论依据。 本研究使用环氧虫啶对室内敏感品系褐飞虱(CYC-S)进行抗性选育,经过29代筛选后,获得对环氧虫啶产生112.14倍的高水平抗性的褐飞虱抗性品系(CYC-R),之后继续对其进行抗性筛选,使其抗性稳定在高抗水平。由此可知,即使环氧虫啶具有独特的作用机制,但连续、单一的药剂选择同样会迫使褐飞虱产生较高的抗性,表明褐飞虱对环氧虫啶存在产生高水平抗性的风险。类似地,室内连续筛选导致灰飞虱Laodelphaxstriatellus对环氧虫啶产生了10.24倍的中等水平抗性[29];棉蚜Aphisgossypii经环氧虫啶室内筛选20代后,其对环氧虫啶的抗性达到了69.50倍[30]。因此,为预防或延缓褐飞虱田间种群对环氧虫啶抗性的产生,亟须明确其对环氧虫啶抗性的遗传方式,用以指导田间合理用药并延长环氧虫啶的使用寿命。 遗传杂交试验表明,抗环氧虫啶褐飞虱品系(101.27倍)对环氧虫啶的抗性为常染色体、不完全显性遗传,该遗传方式理论上更有利于抗性的发展。本文的研究结果与褐飞虱对吡虫啉、噻虫胺、烯啶虫胺和呋虫胺的抗性遗传方式的研究结果类似,均为常染色体控制的不完全显性遗传[31-34]。然而,褐飞虱对氟啶虫胺腈抗性为常染色体、不完全隐性遗传[35],对三氟苯嘧啶抗性为常染色体、共显性遗传[36]。抗性遗传方式的差异可能是不同的药剂选择压、杀虫剂类型和遗传背景等所导致。另外,浓度对数-死亡率几率值线分析及适合性检验表明,褐飞虱对环氧虫啶的抗性由多个等位基因决定,可能与褐飞虱代谢酶以及靶标受体烟碱型乙酰胆碱受体基因的变化有关。在室内筛选的褐飞虱抗性种群中,褐飞虱对吡虫啉、噻虫胺、烯啶虫胺和呋虫胺等新烟碱类杀虫剂的抗性均受多个基因控制[31-34]。已报道的研究表明,虽然多基因遗传抗性的发展速度比单基因遗传抗性慢,但产生的抗性谱较宽,一旦抗性形成后通常比单基因遗传更难控制,且抗性治理难度较大[24, 32, 35, 37]。因此,针对褐飞虱对环氧虫啶的抗性问题,可采用预防性抗性治理策略,在抗性未明显产生前就应限制环氧虫啶的使用次数,与具有不同作用机制的药剂混用或轮用,避免单一药剂的大量连续使用,以延缓褐飞虱对环氧虫啶的抗性演化,保持环氧虫啶在褐飞虱防治过程中的良好防效。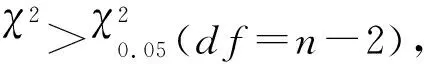
2 结果与分析
2.1 抗环氧虫啶褐飞虱品系选育
2.2 褐飞虱对环氧虫啶抗性遗传方式
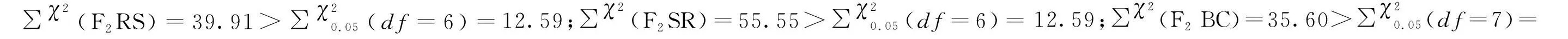
3 结论与讨论
猜你喜欢
云南农业(2024年3期)2024-03-11 10:58:46
贵州畜牧兽医(2023年3期)2023-06-29 07:07:28
中国水稻科学(2023年3期)2023-05-18 02:17:50
农技服务(2023年2期)2023-03-15 00:43:08
昆虫学报(2021年8期)2021-09-06 05:42:40
东坡赤壁诗词(2021年4期)2021-09-05 18:02:41
昆虫学报(2020年8期)2020-10-15 10:50:34
青年文学家(2020年19期)2020-07-26 14:17:36
园林科技(2020年2期)2020-01-18 03:28:18
小学生作文选刊(2017年12期)2017-06-26 12:54:56
