
异源过表达蒺藜苜蓿MtATG7 基因促进拟南芥抵抗碳氮饥饿胁迫
2023-10-12 04:02朱晓洁王贤杰杨明康
华南农业大学学报 2023年5期
朱晓洁 ,王贤杰,杨明康,黄 巍,2,陈 亮,2
(1 华南农业大学 生命科学学院, 广东 广州 510642; 2 岭南现代农业科学与技术广东省实验室, 广东广州 510642)
自噬是真核生物体内一种重要的分解代谢过程。错误折叠的蛋白或受损的细胞器被自噬体包裹运输至液泡,在液泡腔中被水解酶降解[1]。分解产物会被储存于液泡,或被运输回细胞质中重复利用[2]。在模式植物拟南芥Arabidopsisthaliana中,现已鉴定到大约40 个自噬相关基因ATGs(Autophagyrelated genes),且大部分ATGs对自噬体的形成是必需的[3-4]。
研究发现,过表达ATGs可以增强植物的环境适应性。自噬作为物质循环途径之一,对氮素的再利用和营养饥饿的抵抗至关重要。过表达AtATG8a、AtATG8e、AtATG8f和AtATG8g的拟南芥植株氮素再活化能力增强,转基因植株中有更多的氮素从衰老叶片中活化并运输到种子中,从而使种子的蛋白质含量显著提高[5]。拟南芥中异源过表达水稻OsATG8b或苹果MdATG18a均可以增强拟南芥对氮饥饿或碳饥饿的耐受性[6-8]。自噬过程也是植物抗旱、抗盐的关键途径之一。在干旱条件下,过表达MdATG18a和MdATG8i的苹果植株,以及异源过表达香蕉MaATG8f的拟南芥植株,都表现出更强的耐干旱能力,而且在这些过表达植株中,光合效率提高,可溶性糖和氨基酸的积累量明显增加[9-11]。在高盐环境下,过表达MdATG10苹果植株的根系具有更强的水分吸收能力,植株能够保持较高的光合效率从而维持生长[12]。除此之外,过表达MdATG18a的苹果植株不仅对碱性和高温等不良环境的耐受力增强,而且对斑褐病的免疫力也明显提高[13-15]。
自噬相关蛋白A T G 7 通过催化A T G 8-ATG3 偶联物的形成激活ATG8 的脂化作用,被认为是自噬过程中的关键限速因子之一[16]。atg7缺失突变体不能形成自噬体,在碳/氮饥饿条件下生长受阻,叶片早衰,繁殖能力下降[2,17]。过表达AtATG7的拟南芥植株自噬活性显著提高,转基因植株不仅具有衰老延缓、生物量和产量提高、种子含油量增加等优势,而且对病原菌的抗性也明显增强[16]。
苜蓿Medicagospp.是一种广泛种植的饲料作物,同时也是豆科模式植物之一[18]。通过过表达苜蓿ATGs使自噬活性水平提高,是改善苜蓿农艺性状的一种潜在手段。然而苜蓿ATGs的生物学功能是否保守,以及其对植物的抗逆能力是否有促进作用在很大程度上仍然未知。本文以蒺藜苜蓿Medicago truncatula中关键的自噬相关基因MtATG7为研究对象,构建拟南芥异源过表达MtATG7的稳定遗传植株,并在碳/氮饥饿条件下进行表型分析。本研究旨在为利用MtATG7基因改良苜蓿和其他植物的农艺性状提供理论依据。
1 材料与方法
1.1 植物材料与培养条件
拟南芥生态型为Columbia,拟南芥T-DNA 插入的自噬功能缺陷突变体atg7-3(SAIL_11_H07)和GFP-ATG8e植株为Chen 等保存[19],通过杂交得到atg7-3×GFP-ATG8e纯和植株,构建35S::MtATG7-NOS 过表达载体,通过农杆菌Agrobacterium tumefaciens介导侵染Col、atg7-3、atg7-3×GFPATG8e的过表达株系获得MtATG7-ox、atg7/MtATG7、MtATG7/atg7-3/GFP-ATG8e转基因植株。
蒺藜苜蓿生态型为A17。
培养条件:光暗周期为16 h 光照/8 h 黑暗,温度为22 ℃,光照强度为120 μmol·m-2·s-1。
1.2 植物表达载体构建及拟南芥遗传转化
以蒺藜苜蓿cDNA 为模板,利用常规PCR 方法扩增MtATG7基因的编码序列(2 097 bp)。分别在上、下游引物中引入BamHI 和PstI 的酶切位点,通过BamHI/PstI 双酶切插入到双元表达载体pCambia1300,使用无缝克隆试剂盒(金沙生物,SC612)进行连接,完成携带35S::MtATG7的植物表达载体构建。使用农杆菌GV3101 介导的花序浸泡法[20]转化拟南芥Col-0、atg7-3和atg7-3×GFP-ATG8e植株。将转基因拟南芥T0 代种子在含有25 mg/L 潮霉素的筛选培养基上经过3 代筛选获得6 个纯合株系,选取2 个代表株系用于后续试验。所有构建均经测序验证正确,构建引物信息见表1。
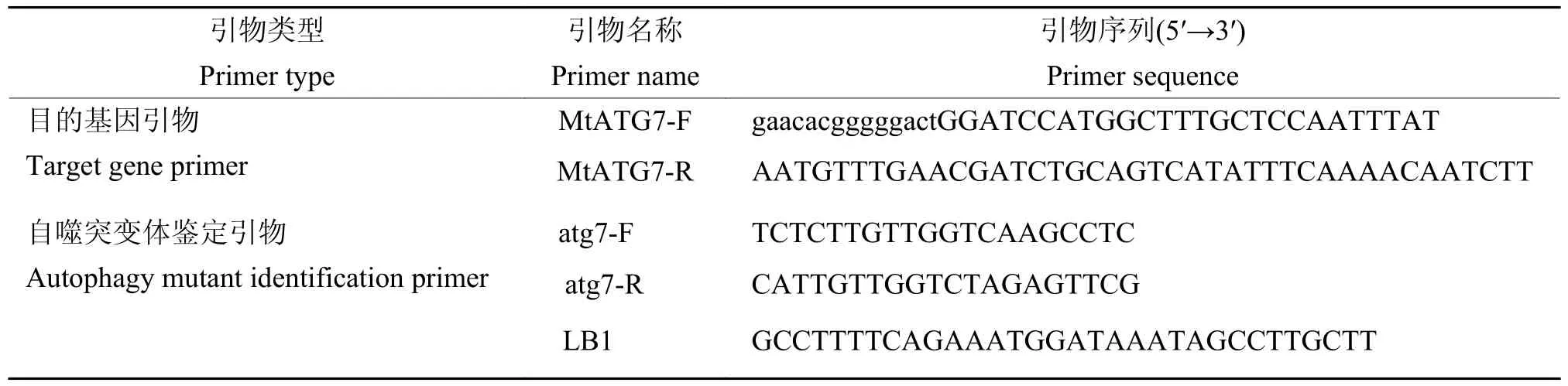
表1 PCR 特异性引物序列Table 1 Specific primers used for PCR
1.3 MtATG7 基因的生物信息学分析
通过Phytozomev13 网站(https://phytozome-next.jgi.doe.gov/)查询不同物种中ATG7基因的氨基酸序列,利用Pfam 数据库(http://pfam.xfam.org/search/sequence)分析不同物种ATG7基因的保守基序;利用MEGA11 软件构建系统发育树。
1.4 缺碳胁迫处理和存活率测定
缺碳处理:将在1/2MS 培养基平板上生长11 d的拟南芥幼苗,转移至无糖1/2MS 培养基平板上,并放置于黑暗环境下培养使植物不能进行光合作用,继续培养9 d 后进行表型观察和拍照。缺碳存活率的测定:将土中生长3 周的拟南芥植株转移至黑暗条件下培养7 d,随后转移到正常生长条件下恢复8 d,统计植株存活率。试验经3 次生物学重复。
1.5 缺氮胁迫处理
拟南芥幼苗在正常1/2MS 培养基平板上生长11 d 后,转移至缺氮1/2MS 培养基平板上,继续培养7 d 后进行表型观察和拍照。
1.6 叶绿素含量的测定
取适量拟南芥幼苗称量鲜质量,加入2 mL N,N-二甲基甲酰胺(N,N-Dimethylformamide,DMF),4 ℃遮光提取2 d,每个样品设3 个生物学重复。按以下公式计算叶绿素含量:叶绿素浓度=20.21×D645nm+8.02×D663nm,叶绿素含量=叶绿素浓度×提取液体积×稀释倍数/样品鲜质量。
1.7 蛋白提取及Western blot 检测自噬活性
取适量2 周龄的拟南芥幼苗,整株取样至2 mL 离心管中,经液氮速冻后,用均质破碎仪研磨样品。加入200 μL RIPA Buffer 并充分振荡混匀,4 ℃放置10 min;4 ℃、12 000 r·min-1离心 30 min,吸取上清。蛋白溶液经95 ℃加热5 min 后,使用变性聚丙烯酰胺凝胶(SDS-PAGE)进行电泳分离。使用PVDF 膜,在4 ℃条件下恒电流200 mA 转膜120 min。将膜置于质量分数为5%的脱脂奶粉中封闭1 h,然后使用Anti-GFP(Invitrogen A11122)抗体4 ℃孵育过夜。1×TBST 洗膜3 次后孵育二抗2 h。1×TBST 溶液重复洗膜3 次,将PVDF 膜放至干净的培养皿中,用显色液试剂盒进行显色后,使用化学发光成像仪拍照保存。
1.8 数据统计分析
试验所得数据使用统计软件SPSS 19.0 进行分析,其中叶绿素含量测定数据n=8,存活率数据n=3,样品之间差异采用单因素方差分析和LSD 多重比较方法进行统计检验,P<0.05 视为差异显著,数据用平均值±标准差表示。
2 结果与分析
2.1 ATG7 蛋白结构在植物中高度保守
为了解蒺藜苜蓿MtATG7的进化特征和生物学功能,本研究对MtATG7的氨基酸序列与拟南芥等其他植物的同源基因构建了系统发育树,并分析了MtATG7及其同源基因的序列相似性。结果表明,在分析的15 种植物中,蒺藜苜蓿的ATG7与红车轴草Trifoliumpratense、鹰嘴豆Cicer arietinum的ATG7亲缘关系最近,与模式植物拟南芥的亲缘关系相对较远。利用Pfam 数据库中对ATG7 的氨基酸序列进行结构域分析,发现在所有15 种植物中都具有典型的ATG7 特征结构域,说明ATG7 的功能很可能是高度保守的(图1)。
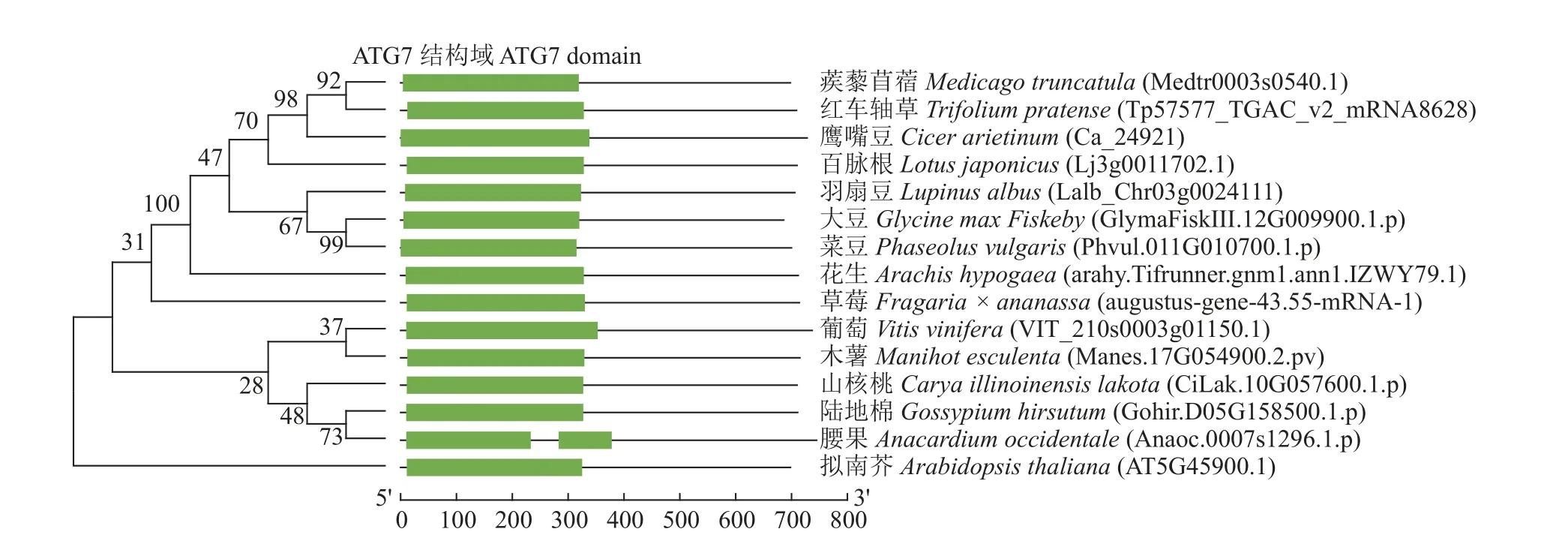
图1 不同植物ATG7 基因的系统发育树分析和保守结构域预测Fig. 1 Phylogenetic tree analysis and domain prediction of ATG7 gene from different species
2.2 MtATG7 基因的克隆及异源过表达植株的遗传转化
根据MtATG7的DNA 序列设计PCR 引物,以蒺藜苜蓿的cDNA 为模板进行PCR 扩增(表1)。PCR 产物经琼脂糖凝胶电泳后得到约2 000 bp 的特异条带,与2 097 bp 的MtATG7编码序列长度吻合,说明MtATG7基因扩增成功(图2A)。进一步使用无缝克隆方法,将MtATG7序列连接到表达载体上,构建了35S::MtATG7双元过表达载体(图2B)。在农杆菌介导下,通过花序浸泡法分别侵染拟南芥野生型(Col)、atg7-3和atg7-3/GFP-ATG8e植株,获得了多个独立的过表达株系,包括MtATG7-OX、atg7/MtATG7和atg7-3/MtATG7/GFP-ATG8e植株。每个基因型植株挑选两个纯合的株系进行后续试验。
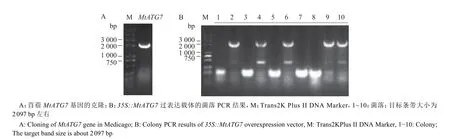
图2 MtATG7 过表达载体的构建Fig. 2 Construction of MtATG7 overexpression vector
2.3 过表达MtATG7 减缓拟南芥叶片在碳饥饿胁迫下的衰老速率
将苗龄11 d 的转基因植株转移至无糖培养基上,并放置在黑暗环境下培养,使植物光合作用停止并产生碳饥饿。在缺碳胁迫条件下,atg7幼苗的叶片明显变黄并伴随白化现象,但野生型Col 与互补植株atg7/MtATG7#1,atg7/MtATG7#2 的叶片仍呈现绿色,说明MtATG7能够挽救atg7突变体的生长缺陷表型(图3A、3B)。过表达植株MtATG7-OX#1 和MtATG7-OX#2 和野生型的叶片一样都保持相对嫩绿。对缺碳胁迫前后的幼苗进行叶绿素含量的测定,结果显示缺碳胁迫后,atg7突变体的叶绿素含量和野生型相比显著下降,但异源转基因的atg7/MtATG7植株和atg7突变体相比叶绿素含量明显提高(图4)。该结果表明MtATG7能够互补AtATG7抵抗碳饥饿胁迫的生物学功能。

图3 MtATG7 促进转基因拟南芥抵抗碳胁迫Fig. 3 MtATG7 promotes resistance to carbon starvation in transgenic Arabidopsis
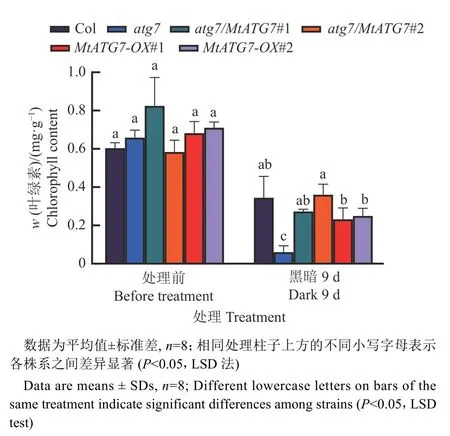
图4 碳胁迫前后的拟南芥幼苗叶绿素含量Fig. 4 Chlorophyll content of Arabidopsis seedling before and after carbon stress
2.4 过表达MtATG7 基因提高拟南芥在碳饥饿胁迫下的存活率
将苗龄3 周的拟南芥转移到黑暗条件下处理7 d,探究过表达MtATG7对拟南芥碳饥饿胁迫存活率的影响。研究发现,在碳饥饿胁迫下,atg7因自噬功能缺陷导致生长受到抑制,即使恢复光照也无法存活,但自噬功能正常的植株都有一定比例植株能够存活下来(图5)。值得注意的是,多个过表达MtATG7的植株,包括atg7/MtATG7#1、MtATG7-OX#1 和MtATG7-OX#2,存活率和野生型Col 相比都明显提高(图6)。上述结果表明过表达MtATG7基因可以提高拟南芥对碳饥饿胁迫的耐受能力。
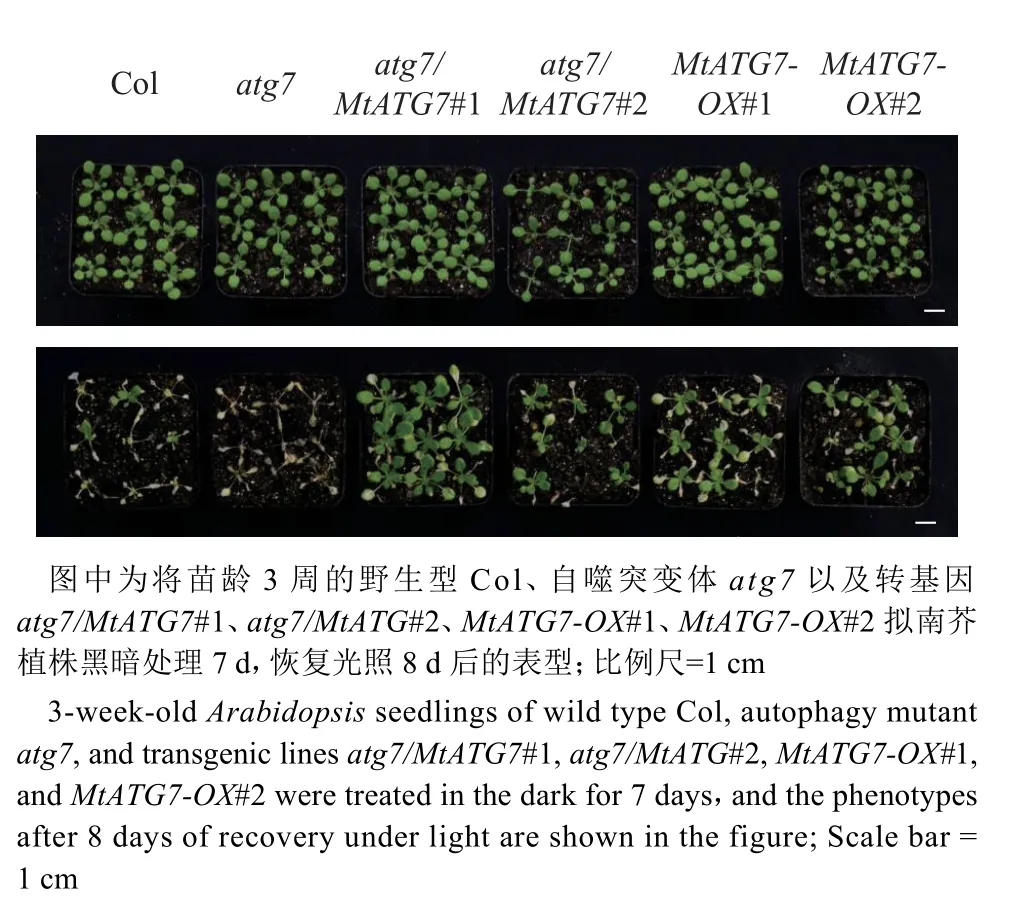
图5 MtATG7 促进转基因拟南芥在碳胁迫下的生存Fig. 5 MtATG7 promotes survival of transgenic Arabidopsis under carbon starvation
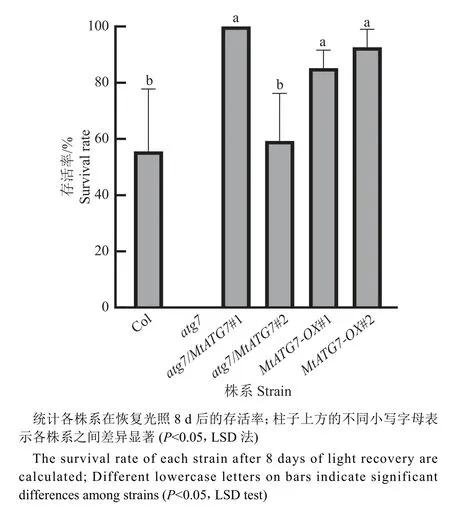
图6 碳胁迫后的拟南芥幼苗的存活率Fig. 6 Survival of Arabidopsis seedlings after carbon stress
2.5 MtATG7 基因能够恢复拟南芥atg7 突变体的自噬活性
GFP-ATG8e 重组蛋白能够被自噬体降解,并释放游离的GFP,因此GFP/GFP-ATG8e 比值是植物自噬活性的重要指标之一。为了探究MtATG7过表达植株是否通过自噬途径增强碳饥饿胁迫耐受性,将GFP-ATG8e、atg7/GFP-ATG8e以及转基因的atg7/MtATG7/GFP-ATG8e置于碳饥饿胁迫下,并检测GFP-ATG8e 蛋白的剪切活性。结果显示atg7突变体在碳饥饿胁迫下,不能产生正常的游离的GFP 条带,而转基因的atg7/MtATG7/GFPATG8e植株的GFP-ATG8 剪切活性能够恢复到接近Col 的水平(图7)。以上结果表明,MtATG7能够互补拟南芥内源AtATG7的功能,自噬活性的恢复很可能是碳饥饿胁迫耐受性提高的原因。

图7 MtATG7 使拟南芥atg7 突变体的自噬活性恢复Fig. 7 MtATG7 restores autophagy activity of atg7 mutant in Arabidopsis
2.6 过表达MtATG7 抑制氮饥饿导致的叶片变黄
将苗龄11 d 的MtATG7过表达植株转移至缺氮培养基上培养,结果表明,在缺氮处理后atg7和野生型相比叶片明显变黄,叶绿素含量下降(图8、9)。而互补植株atg7/MtATG7的叶绿素含量和野生型相近,说明MtATG7能够挽救atg7突变体的缺陷表型(图9)。其中atg7/MtATG7#1 和MtATG7-OX#1 两个株系在缺氮处理后,几乎所有植株的叶片仍然保持绿色,而野生型已有少量植株开始发紫(图8A、8B)。叶绿素测量结果也表明,atg7/MtATG7#1和MtATG7-OX#1 两个株系的叶绿素含量和野生型相比有轻微增加(图9)。

图8 MtATG7 促进转基因拟南芥抵抗氮胁迫Fig. 8 MtATG7 promotes resistance to nitrogen starvation in transgenic Arabidopsis
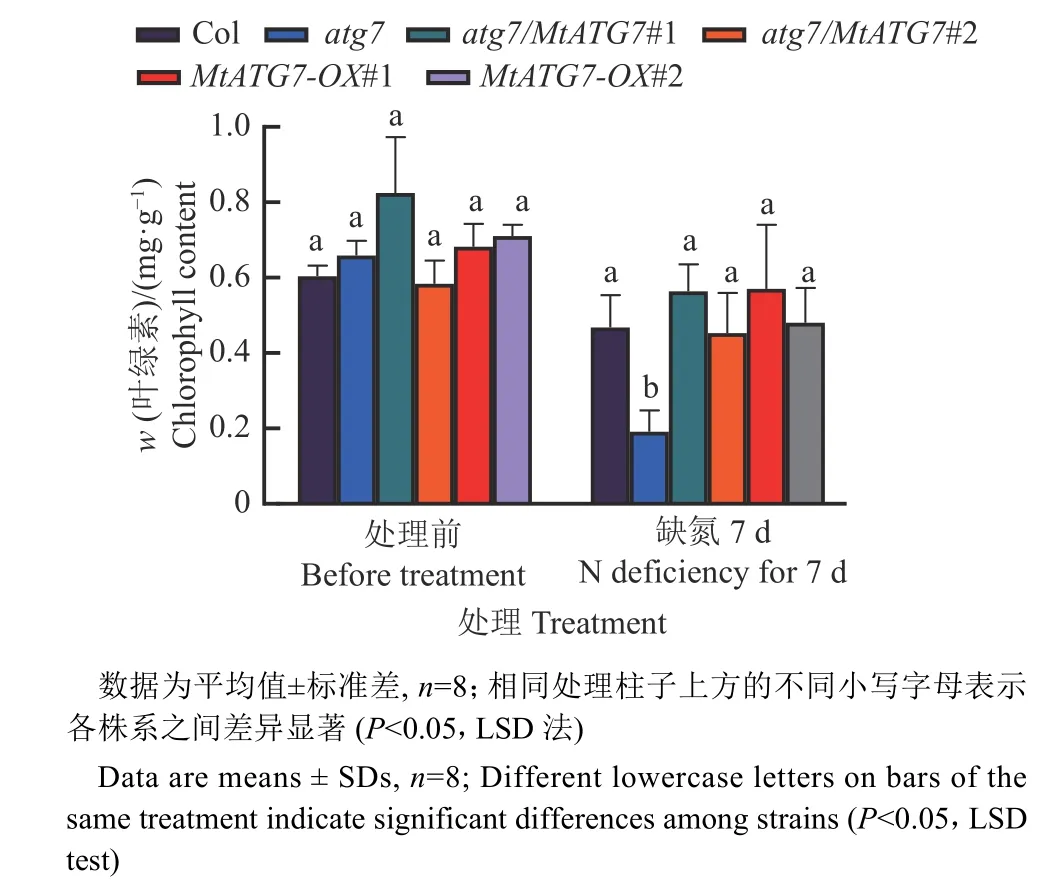
图9 氮胁迫前后的拟南芥幼苗叶绿素含量Fig. 9 Chlorophyll content of Arabidopsis seedlings before and after N stress
3 讨论与结论
碳元素是糖类物质和蛋白质分子的基础,对植物体内能量和物质的稳态至关重要。一方面,植物遭受短期碳饥饿时,细胞自噬能够降解储存于叶片中的瞬时淀粉,为细胞提供额外的能量以渡过逆境[21];另一方面,在长期碳饥饿时,细胞自噬能够降解叶绿体及其中的蛋白质,维持重要组织的氨基酸供应,从而提高植物的生存能力[22]。氮元素是氨基酸、核苷酸、叶绿素的重要组成成分,也是植物生存所必需的元素之一。在低氮条件下,细胞自噬能够将衰老叶片中的氮素分解再活化,使之能够被运输至新生叶片,实现生长发育效率的最大化;同时氮素的再活化也是种子成熟的关键步骤之一[23]。
本研究发现M t A T G 7 蛋白具有保守的ATG7 特征结构域,异源过表达MtATG7能够改善atg7突变体在碳/氮饥饿条件下的叶片早衰性状,而且在缺碳胁迫下MtATG7过表达植株的存活率显著提高,从野生型的55.5%提高至85.2%~92.6%。缺碳条件下,MtATG7能够使atg7突变体的自噬剪切活性恢复,说明MtATG7能够参与拟南芥自噬途径的调控。ATG7 是植物自噬过程的限速酶之一,在自噬过程中分别和ATG12、ATG8 结合,激活ATG8 的脂化过程[24],因此MtATG7很可能通过提高植物自噬水平,使碳/氮饥饿的耐受能力提高。在拟南芥中,ATG7和许多叶片衰老标记基因,例如MYB2、NAP、SAG12、NYE1的表达存在明显关联性,表明ATG7也是植物叶片衰老调控的枢纽之一[25]。在碳/氮饥饿条件下,MtATG7过表达植株叶绿素含量下降的速度减慢,该结果暗示MtATG7还可能通过抑制衰老相关基因的表达,使植物叶片的衰老延迟。
虽然MtATG7基因在参与自噬过程当中具有保守性,但是在不同植物中,自噬途径所影响的生理过程仍然存在特殊性。例如在正常生长条件下,自噬途径的缺失对拟南芥生长和繁殖几乎没有影响;但是在水稻和烟草中,自噬途径受阻会导致其育性明显降低,结实率下降,同时水稻的抽穗时间明显延迟[26-27]。因此过表达MtATG7基因在苜蓿和拟南芥中对抗逆性状的影响是否存在差异,是今后值得探究的课题。
猜你喜欢
金桥(2021年10期)2021-11-05
现代畜牧科技(2021年9期)2021-10-13
现代畜牧科技(2021年4期)2021-07-21
中国三峡(2017年4期)2017-06-06
安徽医科大学学报(2016年12期)2017-01-15
山东农业工程学院学报(2016年6期)2016-12-01
为了孩子(3~7岁)(2016年6期)2016-05-14
文学少年(原创儿童文学)(2016年19期)2016-02-28
文史春秋(2016年8期)2016-02-28
天津医科大学学报(2015年2期)2015-12-22
