
PCSK9介导ApoER2降解在血管平滑肌细胞多倍体化及衰老中的作用
2023-10-12 07:43魏小云刘恒亮姬劲锐郭亚男
医学研究杂志 2023年9期
魏小云 刘恒亮 姬劲锐 郭亚男
冠状动脉粥样硬化性心脏病是目前最重要的死因和疾病负担之一,并且其患病率仍处于持续上升阶段[1]。血管平滑肌细胞是血管壁的主要组分,在各种血管性疾病进程中发挥关键作用[2]。既往研究发现,斑块内血管平滑肌会出现DNA损伤、增殖不良及端粒缩短等早期衰老现象,但引起血管平滑肌细胞衰老的具体机制尚未完全阐明[3,4]。PCSK9(proprotein convertase subtilisin/kexin type-9)是前蛋白转化酶的第9个成员,其可通过PCSK9-低密度脂蛋白受体(low-density lipoprotein receptor, LDLR)轴在胆固醇代谢中发挥关键作用,现已成为降脂治疗的重要靶点[5]。研究表明,循环PCSK9水平与年龄和动脉粥样硬化程度呈正相关,提示其与人体衰老可能关联,但其与血管平滑肌细胞衰老的相关性尚不完全清楚[2,6,7]。
近年来,Komaravolu等[8]研究发现PCSK9的靶蛋白ApoER2参与小鼠主动脉血管平滑肌细胞的有丝分裂过程,ApoER2敲除可以显著抑制血管平滑肌细胞的胞质分裂,并引起小鼠血管平滑肌细胞多倍体的形成及衰老。然而,目前尚未有研究证明PCSK9可以通过介导ApoER2的降解,促进人血管平滑肌细胞多倍体的形成及衰老。因此,本研究采用腺病毒引起PCSK9在血管平滑肌细胞中的过表达,对血管平滑肌细胞增殖及多倍体化的情况进行扫描分析,观察其对人血管平滑肌细胞多倍体形成的影响,并探讨ApoER2在这一过程中发挥的作用。
材料与方法
1.细胞及主要仪器、试剂:(1)细胞:人血管平滑肌细胞购自美国ATCC(货号:CRL-199),用含体积分数10%胎牛血清的Ham′s F12培养液在37℃、体积分数5% CO2培养箱中培养。当细胞贴壁约90%后用0.25%胰蛋白酶消化并传代。(2)主要试剂、仪器:PCSK9腺病毒(Ad-PCSK9)购自美国SignaGen公司(货号:SL103186)。空载腺病毒(Ad-Null)购自美国Vector Biolabs公司(货号:1240)。人重组ApoER2蛋白(recombinant human ApoER2, rhApoER2)购自美国R&D Systems公司(货号:3520-AR)。细胞衰老β-半乳糖苷酶染色(senescence associated β-galactosidase, SAβG)试剂盒购自美国Cell Signaling Technology (货号:9860)。PCSK9、ApoER2及BrdU抗体购自美国Cell Signaling Technology公司。BrdU购自美国Thermo公司(货号:B9285)。倒置荧光显微镜购自德国Leica公司,pH调节仪购自美国HANNA公司,激光扫描细胞仪(laser scanning cytometer, LSC)购自美国CompuCyte公司。
2.实验分组及干预:(1)Ad-Null转染组(空载腺病毒组):细胞均匀传代后,采用空载腺病毒(Ad-Null)转染人血管平滑肌细胞4h,后换用含10%胎牛血清的Ham′s F12培养液继续培养0、6、12、24、36h。(2)Ad-PCSK9转染组(PCSK9腺病毒组):细胞均匀传代后,采用Ad-PCSK9腺病毒转染人血管平滑肌细胞4h,后换用含有10%胎牛血清的Ham′s F12培养液继续培养0、6、12、24、36h。在24h时进行光学细胞形态观察,并检测细胞增殖、衰老及相关蛋白的表达水平。(3)rhApoER2干预(PCSK9腺病毒+ rhApoER2组):用无菌PBS将其稀释成8mmol/L的母液。在Ad-PCSK9转染血管平滑肌细胞前,在细胞培养基中加入rhApoER2,使其终浓度达到80μmol/L,并继续培养24h,并检测细胞增殖和衰老水平。
3.细胞衰老β-半乳糖苷酶检测:采用SAβG试剂盒进行细胞衰老检测。将1×104个细胞接种于35mm2培养皿。分组处理后,弃去培养基,PBS漂洗数次并加入细胞固定液。细胞固定15min后,在培养皿中加入含SAβG染液1.5ml,于37℃恒温箱中孵育过夜。然后在光学显微镜下拍照观察。SAβG染色阳性比率(%)=β-半乳糖苷酶染色阳性细胞数/总细胞数×100%。
4.蛋白水平的检测:冰上裂解细胞30min,置于1.5ml离心管中,并在4℃ 12000r/min(离心半径6cm)的离心机中离心15min。将上清液移至离心管中,DC法定量检测蛋白浓度并将蛋白浓度调整一致后置于煮沸仪上将其变性。每组样本混匀后取65μg蛋白进行SDS-PAGE凝胶电泳,检测内参β-actin,根据β-actin的A值调整上样量,使其与β-actin的A值保持一致。将蛋白以恒压100V后转移至PVDF膜上,用含5%脱脂奶粉的TBST进行封闭,然后加一抗(PCSK9和ApoER2,抗体稀释度均为1∶1000)于4℃冰箱中孵育过夜,加入二抗室温孵育2h,再用TBST洗3次,放入化学发光检测仪中进行检测。应用Gel-Pro Analyzer 4.0图像分析软件检测目的蛋白的A值。
5.LSC扫描分析: 细胞培养基中加入终浓度为10μmol/L的BrdU,并在37℃培养箱中标记1h。弃去培养基,加入1~2ml无水乙醇固定20min后,加入0.1%的Triton X-100常温孵育20min。加入4mol/L HCl并在常温下孵育20min后用枸橼酸钠中和盐酸。加入封闭缓冲液孵育20min,并加入BrdU抗体(1∶50)。常温孵育1h后,加入F488绿色荧光二抗。常温孵育30~60min后,加入PI进行细胞核的复染。常温孵育20min,用75%的甘油进行封片,并在荧光显微镜下进行拍照观察后采用LSC进行血管平滑肌细胞增殖及多倍体化的扫描分析。
6.统计学方法: 应用SPSS 25.0统计学软件对数据进行统计分析,两组间的比较采用t检验,3组间各指标的比较采用单因素方差分析,两两比较采用LSD法(方差齐)或Dunnett′sT3(方差不齐)检验,以P<0.05为差异有统计学意义。
结 果
1.PCSK9过表达对血管平滑肌细胞增殖水平的影响:为探究PCSK9对人血管平滑肌细胞增殖的影响,采用PCSK9过表达腺病毒转染人血管平滑肌细胞。PCSK9在人血管平滑肌细胞中仅有少量表达,而在Ad-PCSK9转染12h后,人血管平滑肌细胞中PCSK9的表达水平显著上调(P<0.05,图1A)。光学显微镜下的细胞形态学观察所示,在转染腺病毒24h后,Ad-PCSK9转染组的血管平滑肌细胞密度显著低于Ad-Null转染组,且多核血管平滑肌细胞(≥4N)的数目显著增加(P<0.01,图1B)。此外,为进一步明确PCSK9过表达对细胞增殖的影响,采用MTT细胞增殖实验证明,Ad-PCSK9转染组血管平滑肌细胞的增殖活性明显低于Ad-Null转染组(0.39±0.03 vs 0.72±0.06,P<0.01)。上述研究的结果提示,过表达PCSK9不仅可以抑制血管平滑肌细胞的增殖,还可诱导多倍体血管平滑肌细胞的形成。

图1 PCSK9过表达对血管平滑肌细胞增殖的影响A.PCSK9的表达水平,与0h比较,*P<0.05,**P<0.01;B.光镜下的细胞形态学改变;C.MTT实验细胞增殖比较
2.PCSK9过表达对血管平滑肌细胞多倍体形成的影响:为明确PCSK9过表达对人血管平滑肌细胞多倍体化的影响,采用BrdU/PI染色法进行染色,并用荧光显微镜和LSC扫描仪进行形态观察和扫描分析。人血管平滑肌细胞在转染Ad-PCSK9 24h后,可以显著下调阳性BrdU细胞的比例(P<0.01,图2中A~D),提示PCSK9过表达可以显著抑制血管平滑肌细胞的增殖。此外,较空载腺病毒组,Ad-PCSK9转染组多倍体血管平滑肌细胞(≥4N)的比例显著增加(P<0.01,图2中A~C、E)。上述的研究结果进一步证明PCSK9过表达不仅可以显著抑制血管平滑肌细胞的增殖,还可以显著促进多倍体血管平滑肌细胞的形成。
3.PCSK9过表达对血管平滑肌细胞衰老的影响:采用SAβG染色法检测血管平滑肌细胞衰老水平。Ad-PCSK9转染组的SAβG阳性血管平滑肌细胞数目显著高于Ad-Null转染组(31.93±7.2 vs 5.46±2.84,P<0.01,图3)。在Ad-PCSK9转染组中,SAβG阳性血管平滑肌细胞多为多倍体血管平滑肌细胞。上述的结果表明,PCSK9过表达诱导的血管平滑肌细胞多倍体化与血管平滑肌细胞的衰老密切相关。

图3 PCSK9过表达对血管平滑肌细胞衰老的影响A.SAβG染色后光镜观察细胞衰老(×200),箭头所指为多倍体细胞;B.细胞衰老水平比较
4.PCSK9过表达对血管平滑肌细胞ApoER2表达水平的影响:既往研究表明,PCSK9可以降解多种LDLR相关膜蛋白,包括LDLR、VLDLR、LRP1和ApoER2[9]。在Ad-PCSK9转染24h后,人血管平滑肌细胞中PCSK9的表达水平显著上调(P<0.01)并显著下调ApoER2的表达水平(P<0.01,图4中A~C)。上述的结果表明,PCSK9过表达可以显著促进ApoER2的降解。
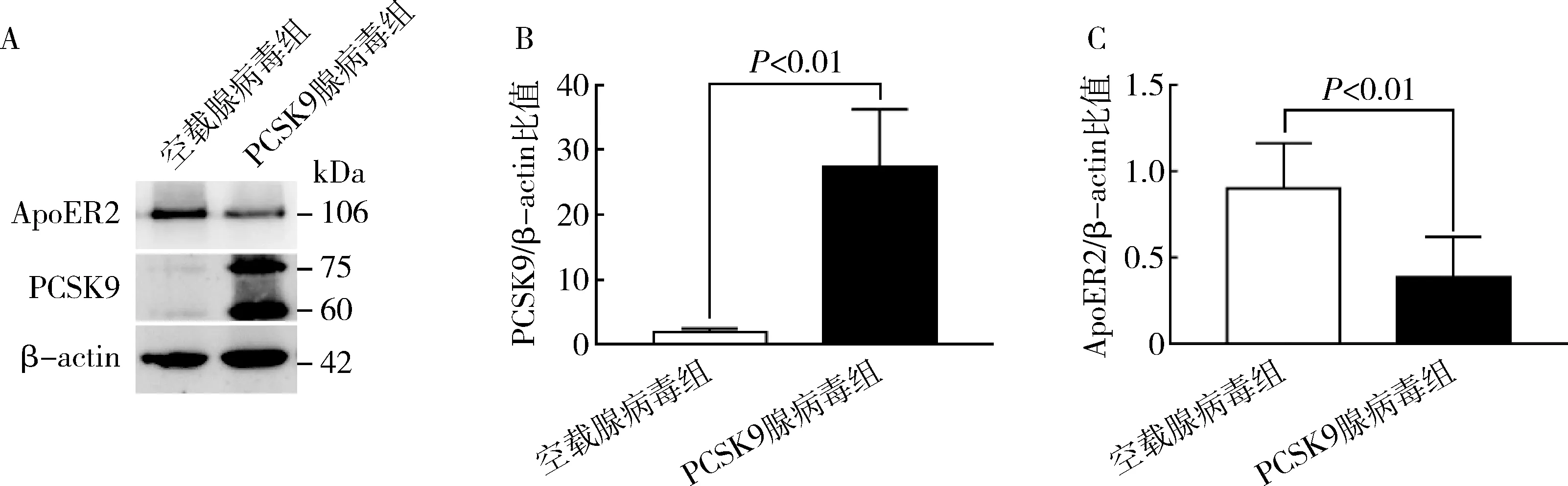
图4 PCSK9对血管平滑肌细胞ApoER2表达的影响A.蛋白表达情况;B.PCSK9的表达水平;C.ApoER2的表达水平
5.rhApoER2对PCSK9过表达诱导的血管平滑肌细胞多倍体化的影响:研究表明,ApoER2敲除可以抑制原代小鼠血管平滑肌细胞的胞质分离并促进血管平滑肌细胞多倍体的形成[8]。为探讨PCSK9介导的ApoER2降解对血管平滑肌细胞的多倍体化及衰老的作用,在Ad-PCSK9转染血管平滑肌细胞前加入可溶性rhApoER2,阻滞PCSK9对血管平滑肌细胞ApoER2的降解,并检测细胞多倍体及增殖情况。rhApoER2可以显著抑制PCSK9过表达诱导的血管平滑肌细胞多倍体化并促进血管平滑肌细胞的增殖(P<0.05,图5)。

图5 rhApoER2对 PCSK9过表达诱导的血管平滑肌细胞多倍体的影响A.LSC细胞扫描分析;B.多倍体细胞比例;C.增殖细胞比例
讨 论
血管平滑肌细胞是动脉粥样硬化斑块纤维帽的主要组分,血管平滑肌细胞的衰老可以减少纤维帽中血管平滑肌细胞组分,进而促进动脉粥样硬化斑块病变的进展[10,11]。近年来研究表明,PCSK9在人动脉粥样硬化斑块中的表达上调,而敲低PCSK9的表达可通过抑制TLR4/NF-κB信号通路的激活,从而减缓Apoe-/-小鼠动脉粥样硬化病变的进展,而血浆胆固醇水平却无显著改变[12]。上述研究结果表明,PCSK9可以通过非LDLR途径促进动脉粥样硬化病变的进展。本研究发现,PCSK9在血管平滑肌细胞中仅少量表达,而Ad-PCSK9转染人血管平滑肌细胞,使PCSK9过表达可通过抑制血管平滑肌细胞的增殖,并诱导血管平滑肌细胞多倍体的形成和衰老。此外,本研究还发现,PCSK9在血管平滑肌细胞中的过表达还可以显著抑制ApoER2的表达水平,而rhApoER2则可以抑制PCSK9诱导的多倍体血管平滑肌细胞形成。本研究结果提示,PCSK9可能通过诱导ApoER2的降解,促进血管平滑肌细胞多倍体的形成并诱导其发生衰老。
既往研究表明,斑块内血管平滑肌细胞会出现广泛的DNA损伤、端粒缩短及端粒酶的活性降低, 引起增殖不良,但其潜在机制尚不明确[4]。PCSK9是Kexin样前蛋白转化酶枯草杆菌蛋白酶家族的第9个成员,其在血脂代谢的过程中发挥重要作用,越来越多的证据表明PCSK9广泛参与多种心血管疾病的病理进程,如冠状动脉粥样硬化性心脏病、腹主动脉瘤、主动脉瓣钙化等[13~15]。此外,近年来研究表明,两种长寿相关蛋白SIRT1和SIRT6可以PCSK9依赖的方式调节血脂代谢[16,17]。然而,目前尚未有研究证实PCSK9与血管平滑肌细胞衰老之间的相关性。本研究发现,PCSK9可以通过诱导血管平滑肌细胞多倍体的形成,抑制其增殖,并促进其发生衰老,这为阐明PCSK9调控血管平滑肌细胞衰老的机制提供新的理论依据。
多倍体是指细胞整个基因组拷贝数增加(>2N)的状态,通常发生在植物细胞,在动物肝细胞、子宫平滑肌细胞、甲状腺细胞、高血压大鼠模型平滑肌细胞中也有发现[18,19]。有研究表明,细胞多倍体化参与血管平滑肌细胞凋亡和衰老的激活,但引起血管平滑肌细胞多倍体化的信号和潜在机制仍未完全阐明[3,8,20]。胞质分裂是细胞周期的最后一步,阻滞胞质分裂可以引起多倍体的形成,Komaravolu等[8]研究发现,ApoER2参与原代小鼠主动脉血管平滑肌细胞有丝分裂过程中后期促进复合物(anaphase-promoting complex, APC)/细胞分裂周期蛋白20(cell division cycle 20, CDC20)复合物的形成,ApoER2敲除可以显著抑制血管平滑肌细胞的胞质分裂,并引起血管平滑肌细胞的多倍体化及过早衰老。既往研究表明,ApoER2是LDLR家族成员,与LDLR的一级序列具有同源性,PCSK9催化结构域可以结合并促进LDLR家族成员的降解[9]。本研究结果显示,PCSK9过表达可以显著下调ApoER2在血管平滑肌细胞中的表达水平,加入的可溶性rhApoER2阻滞PCSK9对血管平滑肌细胞ApoER2的降解后可以逆转PCSK9过表达诱导的血管平滑肌细胞多倍体化。因此,推测PCSK9可能通过介导ApoER2的降解,抑制血管平滑肌细胞的胞质分裂,进而导致血管平滑肌细胞的多倍体化和衰老。
综上所述,本研究发现PCSK9过表达不仅显著抑制血管平滑肌细胞的增殖,还可引发血管平滑肌细胞的多倍体化和衰老。机制上,PCSK9可能通过促进ApoER2在血管平滑肌细胞中的降解,引发血管平滑肌细胞的多倍体化。这些发现进一步揭示了PCSK9在血管平滑肌细胞中的生理作用,并提示PCSK9单克隆抗体不仅可以降低患者血脂水平,还在改善血管老化方面发挥潜在作用。
猜你喜欢
传染病信息(2022年3期)2022-07-15
今日农业(2020年14期)2020-12-14
诊断学(理论与实践)(2020年6期)2020-12-09
中国临床医学影像杂志(2019年4期)2019-06-18
浙江中西医结合杂志(2018年12期)2018-12-27
灾害医学与救援(电子版)(2018年1期)2018-06-05
上海农业学报(2017年4期)2017-04-10
临床与实验病理学杂志(2017年3期)2017-03-07
罕少疾病杂志(2016年5期)2016-03-11
吉林大学学报(医学版)(2015年4期)2015-12-17
