
病原哈维氏弧菌(Vibrio harveyi)TS-628菌株基因组分析
2023-10-10 02:23李辰睿徐荣亮张小旭陈东萍邱于洋陈伟琴覃映雪
渔业研究 2023年5期
李辰睿,徐荣亮,张小旭,陈东萍,邱于洋,陈伟琴,覃映雪
(集美大学水产学院,福建 厦门 361021)
哈维氏弧菌(Vibrioharveyi)为嗜盐性革兰氏阴性菌,隶属于弧菌科(Vibrionaceae)、弧菌属(Vibrio),在海洋环境中广泛分布,是海水鱼虾养殖过程中常见的条件致病菌[1-4]。受该菌感染后的对虾通常会出现肌肉白灼、红腿等症状[2,5],在幼体阶段被感染的对虾死亡率高达100%[6]。哈维氏弧菌还可通过伤口或消化道感染鱼体,鱼类被感染后通常会出现眼球突出、角膜不透明、充血,肠胃炎,肌肉溃烂,鳍条基部、肛门红肿等症状,更甚者直接死亡[7]。目前已报道哈维氏弧菌可感染的鱼类包括大黄鱼(Larimichthyscrocea)[8]、鲈鱼(Centrupomusundecimalis)[9]、斜带石斑鱼(Epinepheluscoioides)[10]、大西洋牙鲆(Paralichthysdentatus)[11]、半滑舌鳎(Cynoglossussemilaevis)[12]等重要海水经济鱼类。此外,哈维氏弧菌还可作为机会病原体感染人类,导致腹泻、败血症、中耳炎等[13]。因此,对哈维氏弧菌致病机制的研究受到广泛关注。
目前关于哈维氏弧菌致病机制的研究已取得较大进展。研究发现,哈维氏弧菌的主要毒力因子包括溶血素、胞外酶(如半胱氨酸蛋白酶)、外膜蛋白、脂多糖、酯酶、铁载体及噬菌体介导的毒力基因等等,这些毒力因子相互协调,直接或间接参与致病过程,诱发对宿主有害的病理反应[14-15]。同时,研究也发现,不同来源的哈维氏弧菌菌株的毒力存在较大差别,因此有必要对不同菌株的毒力及致病机制进行更为有针对性的研究,以获得更为有效的防治方法[16]。
随着测序技术的快速发展,通过全基因组测序开展毒力因子研究已成为揭示病原菌致病机理的重要技术手段之一[17]。本文以分离自福建海域的患溃疡病青石斑鱼(Epinephelusawoara)病原哈维氏弧菌TS-628菌株为研究对象,对该菌株进行基因组测序及序列分析,预测其基因功能,挖掘毒力相关基因,以期为哈维氏弧菌致病机制以及哈维氏弧菌病防治技术的研发提供理论基础。
1 材料和方法
1.1 供试菌株
哈维氏弧菌TS-628菌株分离自福建海域患溃疡病的青石斑鱼[18],并保存于-80 ℃冰箱中。
1.2 实验材料
LB培养基(2% NaCl)、TCBS培养基,细菌基因组提取试剂盒。
1.3 菌株的纯化与扩大培养
将上述菌株从-80 ℃冰箱中取出,先在TCBS培养基上使用平板划线法分离单菌落,30 ℃恒温培养箱培养24 h。挑取单菌落后接种于LB液体培养基中,在180 r/min、28 ℃条件下振荡培养24 h。
1.4 菌株基因组DNA提取
使用细菌基因组DNA提取试剂盒(湖南艾科瑞生物工程有限公司生产),参照说明书中革兰氏阴性细菌基因组抽提的方法进行相应操作,利用1%琼脂糖凝胶电泳检测提取的基因组DNA的完整性。
1.5 基因组测序
将电泳检测合格的DNA样品送测序公司测序,主要操作步骤如下:采用超声波破碎仪随机打断基因组DNA,收集满足要求的DNA片段(400~500 bp),对DNA片段的3’端加碱基A,与3’端带有T碱基的接头进行连接,以构建合格的基因文库,完成整个文库制备。基因组测序采用第二代Illumina Hiseq平台与第三代 PacBio平台,每个样品需同时提供不低于基因组100×的Illumina PE150测序数据和100×的PacBio测序数据,以避免单纯插入10~20 kb片段的PacBio文库造成小质粒(<15 kb)信息的丢失,保证更完整精确的组装。测序原始数据上传至NCBI数据库,登录号PRJNA874896 (https://www.ncbi.nlm.nih.gov/bioproject/PRJNA874896)。
1.6 生物信息分析流程
1.6.1 基因组组分分析
使用GennMarkS软件[19]对基因组进行编码基因的预测,使用CGView Server[20]绘制基因组图解。
1.6.2 致病性和耐药性分析
为从基因组水平了解菌株的毒力因子和耐药性情况,分别使用致病菌毒力因子数据库(Virulence factors database,VFDB)对TS-628菌株基因组的毒力因子基因进行鉴定;使用耐药基因数据库(Comprehensive antibiotic resistance database,CARD)对耐药基因进行分析;使用Effective T3软件及在线工具[21]对其分泌系统进行预测分析。
2 结果与分析
2.1 哈维氏弧菌TS-628菌株的基因组组装分析
统计组装序列的GC含量及reads覆盖平均深度以分析基因组的GC偏向性及重复序列,分析结果如图1所示,菌株TS-628的基因组含量分布趋向一致,GC含量集中在40%~50%的范围内,且图中没有明显横向区块分布,说明此数据样本不存在其他物种污染。测序结果显示,TS-628菌株的基因组含2条染色体和2个质粒,长度为6.15 Mb,GC含量为44.68%(表1),共编码基因5 658个。2个主染色体的基因组长度为5.87 Mb,平均GC含量为44.86%。在2个质粒中,1个质粒为已知,1个质粒为新发现质粒(表2)。已知质粒序列较大,长度为217 kb;新发现质粒长度为66 kb,其碱基分布为:组装结果的GC含量占总基因含量的44.56%,与染色体GC%较为接近。

表1 TS-628菌株基因组组装结果Tab.1 The genome assembly results of strains TS-628
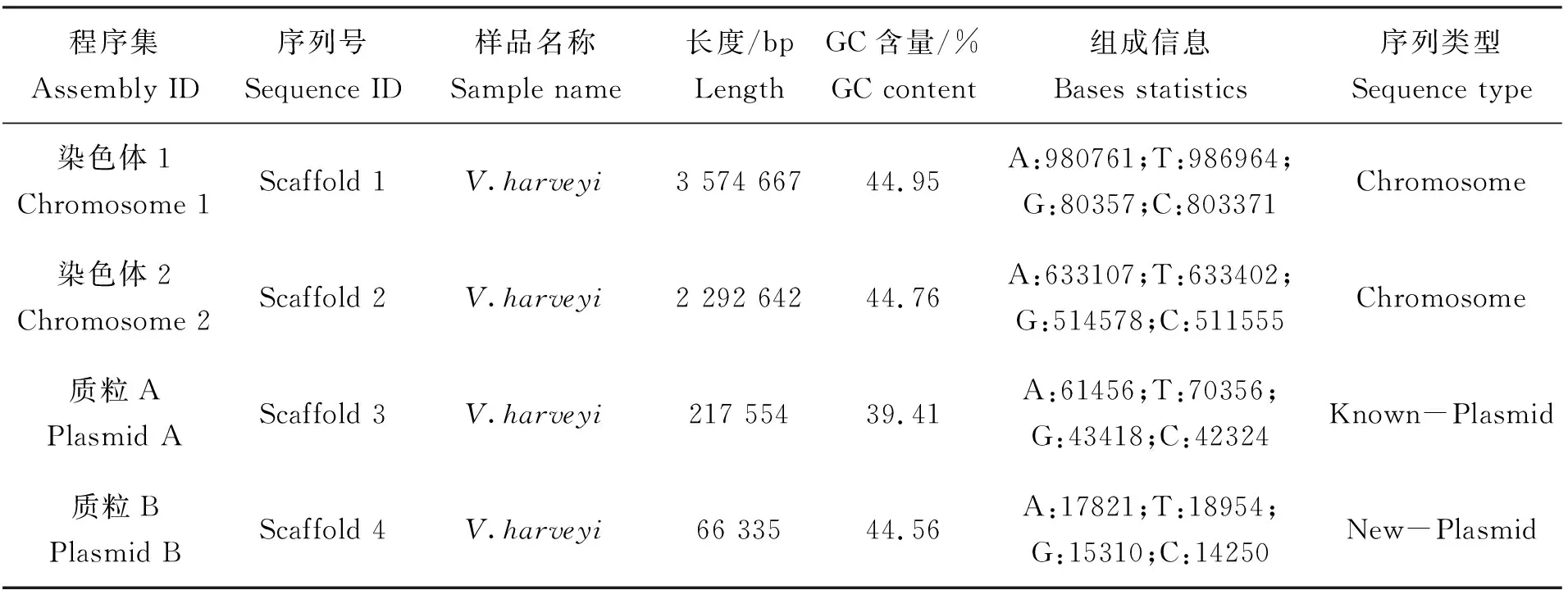
表2 TS-628菌株基因组组装详情Tab.2 The genome assembly details of strains TS-628

注:散点图的横坐标为GC含量,纵坐标为reads覆盖深度;右、上两侧直方图,分别为GC含量、测序深度的滑窗频数分布。Notes:The abscissa of the scatter chart is the GC content,and the ordinate is the read coverage depth;the histogram at right and up sides shows the frequency distribution of sliding window of GC content and sequencing depth,respectively.
2.2 基因组圈图
通过CGView软件绘制菌株TS-628组装架构的基因组圈图(图2),发现新发现质粒总长度为66 335 bp,不存在tRNA基因。根据CG View图的COG注释结果,发现质粒B(图2D)含功能基因种类较少,在二十六类基因功能中,关于胞内转运、分泌和小泡运输的相关基因最多,此类蛋白基因与菌株介导生物膜内外化学的化学物质及信号交换有密切联系;其次是关于复制、重组和修复的相关基因,此类蛋白基因会影响菌株稳定遗传和基因的损伤修复。此外,新发现质粒还含有少量关于转录、细胞周期控制、细胞分裂和染色体分裂以及碳水化合物转运代谢的相关基因,并且占其基因组比例约10%的蛋白基因具有的潜在的生物学功能未被注释到数据库中。
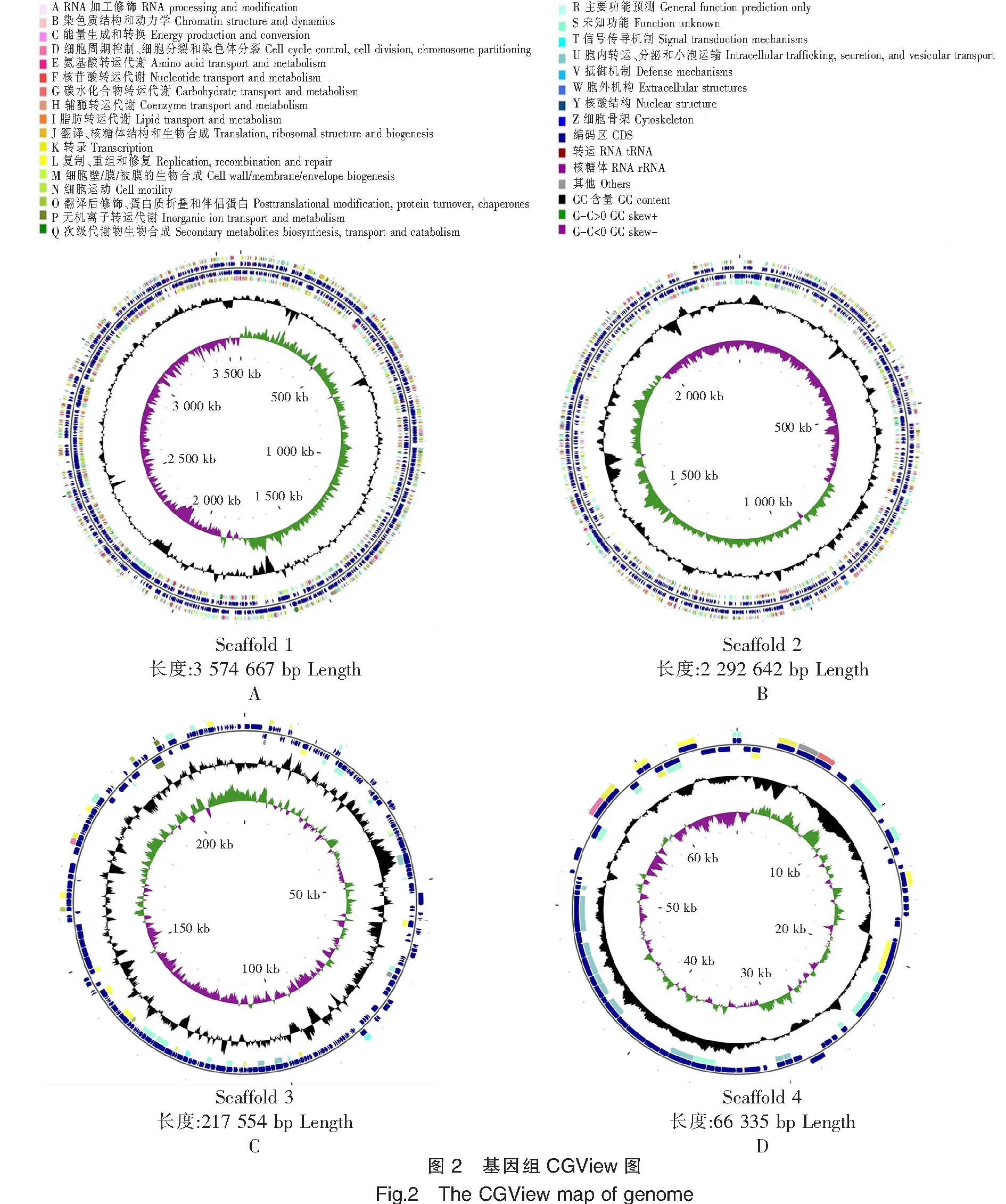
注:图2A为染色体1;图2B为染色体2;图2C为质粒A;图2D为质粒B。从内圈到外圈,第1圈代表基因组全长;第2圈表示GC skew,其中绿色表示G-C>0,紫色则相反,绿色和紫色的交接处分别表示复制起始点和终止点;第3圈黑色表示GC含量,峰值向外突出的部分表示该区域GC含量高于全基因组平均GC含量,向内则相反;第4圈和第5圈分别表示正、负链上的基因,其中包括tRNA、rRNA、CDS和其他基因。Notes:Figture 2A is chromosome 1;Figture 2B is chromosome 2;Figture 2C is plasmid A;Figture 2D is plasmid B.From the inner circle to the outer circle,the first circle represents the total length of the genome;the second circle represents the GC skeleton,where green represents G-C>0,while purple represents the opposite,the intersection of green and purple represents the start point and end point of replication,respectively;the black in the third circle indicates the GC content,and the protruding part of the peak indicates that the GC content in this region is higher than the average GC content of the whole genome,while the opposite is true inward;the fourth and fifth circles respectively represent genes on the positive and negative chains,including tRNA,rRNA,CDS and other genes.
质粒A(图2C)含有的功能基因种类也较少,主要有关于复制、重组和修复,以及无机离子转运代谢和胞内转运、分泌和小泡运输的相关基因。2个染色体(图2A、图2B)含有的功能基因种类较为丰富,主要包括能量生成和转换,氨基酸转运代谢,碳水化合物转运代谢,翻译、核糖体结构和生物合成以及细胞运动相关基因,这些种类的蛋白基因与菌株的各类生命活动密切相关,影响着细菌的能量供应、物质代谢与稳定传代。
2.3 基因组组分分析
根据基因长度分布图结果进行分析,发现长度低于200 bp的基因数量较少,共有200个;在200 bp至1 000 bp的长度区间内,各长度基因数量差距较小,平均基因数量为432个,基因长度分布集中在大于1 000 bp,有2 003个(图3),约占编码基因的35.4%。

2.4 毒力基因预测
将哈维氏弧菌TS-628基因组序列通过VFDB数据库进行比对,预测TS-628菌株共有毒力基因747个,其中附着类相关的毒力基因数量最多为132个,铁摄取相关的毒力基因数量为130个,分泌系统相关的毒力基因数量为107个,抗吞噬类的毒力相关基因数量为36个,调控相关的毒力相关基因数量为35个,细菌毒素的毒力相关基因数量为30个,其他类型的毒力相关基因数量均小于30个(表3、图4)。

表3 毒力基因预测分类表Tab.3 The prediction and classification of virulence genes

2.5 分泌系统及调控系统分析
根据预测结果,TS-628存在多种分泌系统,按照分泌系统相关基因的数量进行分析,TS-628的分泌系统应以Ⅱ型、Ⅲ型、Ⅵ型和Sec-SRP型为主(表4、图5)。同时该菌株具有多个双组份调控系统(表5),其中包含149个调控子、63个感应子以及30个具有混合功能的双组分调控基因。

表4 分泌系统分类统计表Tab.4 The classification and statistics of secretory system

表5 双组分调控分类统计表Tab.5 The statistics of two-component regulation classification
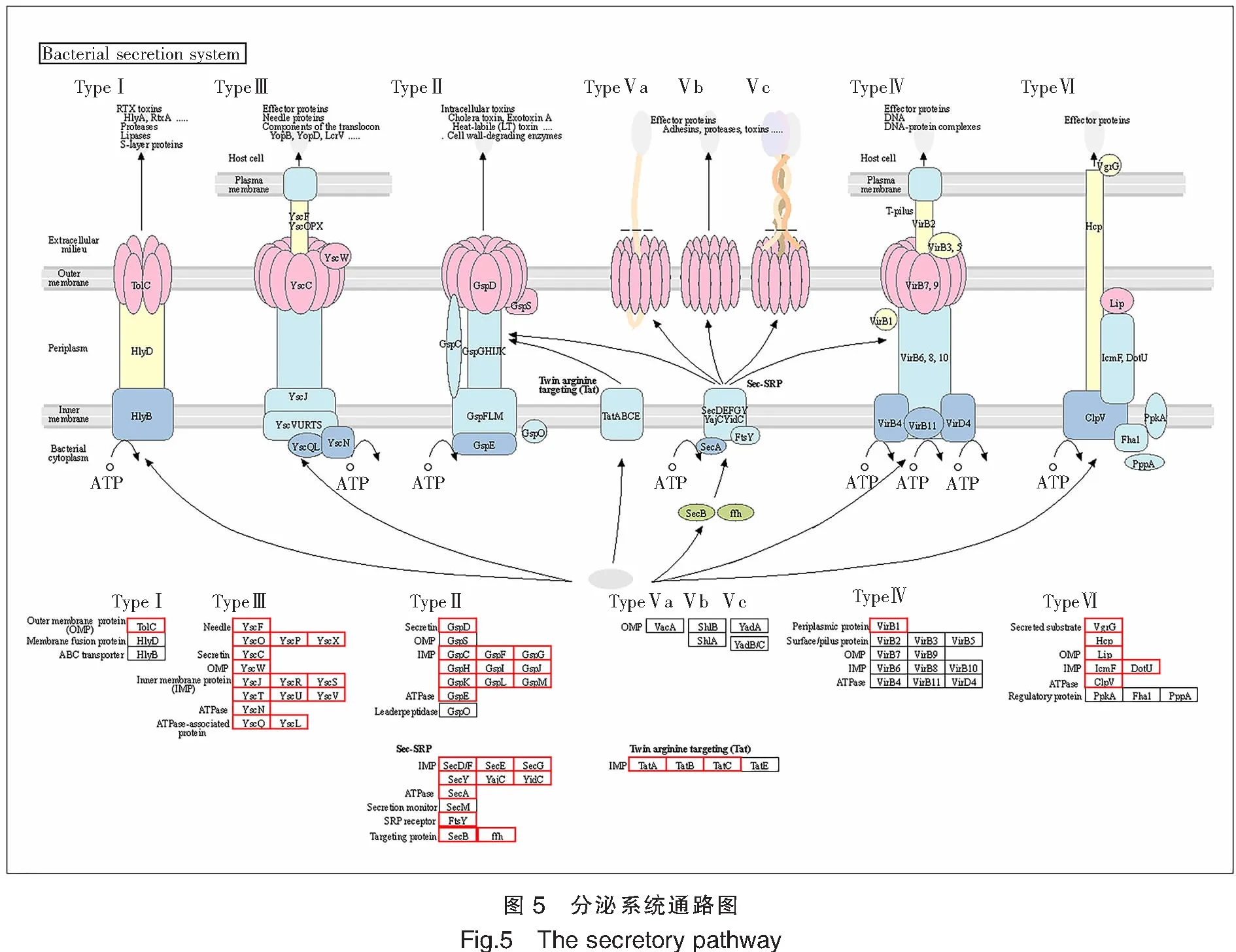
注:分泌系统通路图对每个样本中所有分泌系统的图示化展示。其中红色框标记的是在该样本中预测到的分泌系统相关基因。Notes:The secretory system pathway diagram is a graphical representation of all secretory systems in each sample.The red box marks the genes related to the secretion system predicted in the sample.
2.6 耐药基因预测
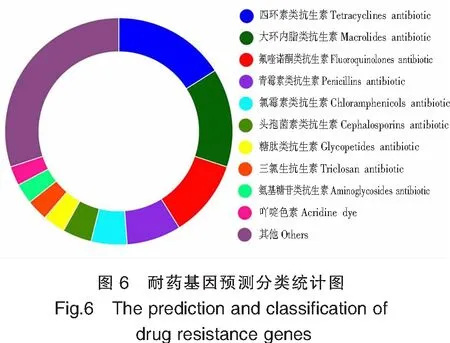
通过CARD进行比较、注释,将读数直接映射至参考数据库来预测耐药性决定因素。结果预测TS-628共携带耐药基因294个,至少对11种抗生素耐药。耐药基因预测分类统计见图6。TS-628对四环素、大环内酯、氟喹诺酮和青霉素4类抗生素具有较多抗性基因。此外,对氯霉素类、头孢菌素类、糖肽类、三氯生类、氨基糖苷类等其他至少7类抗生素有抗性基因。
3 讨论
3.1 基因组概况分析
在基因高效、快速的探索中,基因组测序技术已被广泛运用。如徐先栋[22]利用全基因组测序技术对华南海水养殖地区哈维氏弧菌GDH11385进行了测定,并对该菌株基因组序列进行了基因组注释、基因功能分类、细菌关键毒力基因等分析,为哈维氏弧菌病的防治研究提供了有力的数据支撑。
一般对于原核生物细菌而言,基因组的序列越长,表达的蛋白越多。目前在NCBI的数据库中已公布24株哈维氏弧菌的全基因组序列,如Wang Z等[23]于2015年公布的V.harveyiATCC 33843的基因组全长5.881 490 Mb,表达基因数量为5 332个。Moriyama E N等[24]的研究显示原核生物较长的基因长度会表现出对高效翻译密码子的偏好,可改善翻译效率,进而影响细菌对环境的适应性。本研究TS-628的序列大小比多数公布序列大,为6.151 198 Mb,编码基因数量为5 658个,且基因长度分布集中在大于1 000 bp,约占编码基因的35.4%,综合分析认为TS-628菌株的长基因序列增强了该菌株对环境变化的适应性,可能具有更为复杂的生理生化等功能。
质粒是附加到细胞中的非细胞的染色体或核区DNA原有的能够自主复制的较小的DNA分子。质粒天然存在于某些生物体细胞中,一个细胞里可同时有一种或多种质粒存在。Tao Y等[25]的实验表明大肠杆菌的质粒上含有抗四环素基因;Levinson B L等[26]和Singer J T等[27]均发现质粒携带的基因可以赋予细胞额外的生理代谢能力,提高细菌的致病力。本研究的哈维氏弧菌TS-628菌株的基因组共含2条染色体和2个质粒,其中1个质粒为新发现质粒,长度为66 kb。经分析发现,菌株TS-628的2个主染色体携带关于各类物质合成和代谢相关的基因数量较多,推测2个染色体影响着细菌的自身生长和繁殖,新发现质粒携带接合转运与复制重组的相关基因较多,推测该质粒对细菌生物膜形成、驱动细菌菌株适应环境和快速进化[28]以及在细菌感染宿主的过程中的快速繁殖稳定传代方面有重要意义。
3.2 毒力因子分析
哈维氏弧菌的致病性与多种毒力因子有关,包括铁摄取系统、群体感应机制、类细菌素物质、对噬菌体感染的敏感性等毒力因子[29-30],研究哈维氏弧菌的致病及耐药基因将为确定其致病及耐药机制提供重要的基础。VFDB数据库整合了病原细菌毒力因子信息,通过数据库注释可获得每个样本中含有的毒力基因信息。
已有研究发现,在哈维氏弧菌的致病机制中,宿主物种不同会影响菌株毒力菌株的变异性[31]。在NCBI数据库已公布全基因组序列的24株菌株中,只有少数菌株进行了关于毒力机制的分析,其中菌株QT-520[32]分离自中国桥头深水网箱养殖的患病卵形鲳鲹(Trachinotusovatus),其含有附着类、抗吞噬类、毒素相关的毒力基因数量较多;菌株345[33]分离自中国深圳养殖污染区的感染石斑鱼,含有附着类、免疫逃避、运动性相关、分泌类毒力基因数量较多;本研究表明哈维氏弧菌TS-628基因组携带多种与调节细菌黏附作用、细菌持续感染和营养代谢等相关的毒力因子。通过对比分析发现(表6),分离自不同鱼类的哈维氏弧菌均具有2条染色体,而具有的质粒数量有所不同。3株不同种类宿主的哈维氏弧菌的主要毒力因子类型也有一定差别,其中与附着相关的毒力基因在3株菌株中均存在,说明附着相关毒力因子在哈维氏弧菌致病过程普遍发挥作用,铁摄取和分泌相关基因也是病原性哈维氏弧菌较常见的毒力因子。

表6 3株哈维氏弧菌主要毒力因子比较分析表Tab.6 Comparative analysis table of the main virulence factors of 3 strains of V.harveyi
病原菌入侵宿主是引起疾病感染的首要条件,其首先黏附于宿主体表,才能达到增殖、感染的目的。研究已发现菌毛[34]、外膜蛋白[35]、血凝素[36]等物质参与黏附作用。本研究TS-628的附着类相关毒力基因有132个(17.7%),为预测毒力基因数量最多的一类,推测该菌可通过增强黏附能力而在感染早期便对宿主进行黏附。
铁元素是大多数微生物生存所必需的重要营养元素。铁离子不仅是许多基础代谢通路的辅助因子,还是合成许多关键代谢酶类所必需的金属元素辅基,可与蛋白质特异性结合,参与DNA合成、基因调控、电子传递等生命活动过程[37-38]。细菌在宿主体内的生存和毒力发挥都离不开铁,但环境中游离的铁离子浓度远低于其生长代谢所需的浓度[39],故铁元素是病原菌和宿主必须争夺的一种重要营养元素。铁载体(Siderophores)是由细菌和真菌在低铁环境中所产生的一类可与铁离子特异性结合的螯合剂,与致病微生物的毒力机制有关。Naka H等[40]发现铁载体能够帮助病原菌从宿主的铁结合蛋白上摄取铁,并可通过特异性的细胞膜受体和内膜上的转运系统将铁运输至细胞中并协助细菌生长代谢。本研究TS-628的铁摄取相关毒力基因有130个(17.4%),推测其在感染过程中会通过增强各类铁摄取系统的表达,掠夺宿主体内的铁元素及含铁蛋白,一方面有利于自身存活与繁殖,另一方面对宿主产生毒害,影响宿主体内的需铁代谢,从而导致感染的发生。
细菌分泌系统是一种重要的毒力因子,可将各种细菌毒力因子转运到细菌细胞外,并具有与所处环境交流的功能。目前已发现8大细菌分泌系统[41],如T1SS主要分泌黏附素、蛋白酶以及毒力因子等各种蛋白质到宿主细胞中[42]。哈维氏弧菌一般含有4至5种分泌系统,如Huang W等[43]测得的分离自石斑鱼体内的致病哈维氏弧菌菌株ZJ0603基因草图数据中显示其含有Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅵ型分泌系统,Espinoza-Valles I等[44]等对分离自对虾体内的致病哈维氏弧菌菌株CAIM1792进行测序,也得到相同结果。本研究对哈维氏弧菌TS-628基因组进行毒力基因筛查,发现其和哈维氏弧菌菌株ZJ0603、CAIM1792一样,均含有Ⅰ、Ⅱ、Ⅲ、Ⅳ和Ⅵ型5种细菌分泌系统,同时,按照分泌系统相关基因的数量分析,TS-628的分泌系统应以Ⅱ型、Ⅲ型、Ⅵ型和Sec-SRP型为主。
Ⅱ型分泌系统(T2SS)的分泌底物主要是蛋白酶、纤维素酶、几丁质酶和各种外毒素等[45-50]。T2SS将分泌底物从细胞质转移到周质空间后,再将底物分泌到细胞表面或细胞外环境中,参与生物膜的形成、养分的摄取、定植和入侵,继而赋予细菌生存优势、参与细菌的致病机制[51]。Ⅲ型分泌系统(T3SS)是许多致病性革兰氏阴性菌的关键毒力因子[52-54],可在转移效应分子以及改变宿主免疫反应方面发挥作用。T3SS通过直接将细菌胞浆毒力蛋白注入宿主细胞而参与到细菌的致病机制中[55]。Ⅵ型分泌系统(T6SS)的尾部复合物中具有一尾鞘结构,可刺穿宿主细胞壁和细胞膜,直接进入宿主细胞质中释放毒性效应蛋白,在细菌竟争和侵染寄主发病方面有重要作用[56-58]。鉴于此,推测哈维氏弧菌TS-628菌株可通过破坏宿主黏膜、免疫系统以及影响宿主生物膜形成等方式产生致病作用。
4 结论
本文通过基因组测序和基因组功能注释分析获得了哈维氏弧菌TS-628的基因组组成、毒力因子和耐药因子等信息。该菌株基因组长度约为6.15 Mb,GC含量为44.68%,共编码5 658个基因,共有2条主染色体、2个质粒(其中1个为新发现质粒)。预测共有毒力基因747个,耐药基因294个,存在多个分泌系统,具有多个双组份调控系统。根据基因组信息推测,哈维氏弧菌TS-628菌株可能首先通过强大的黏附能力黏附到宿主上,再通过分泌系统等毒力因子对宿主细胞和组织造成损伤,进而突破宿主皮肤、黏膜生理屏障,之后通过各类铁摄取系统与宿主竞争铁等重要元素,在宿主机体内定植、繁殖和扩散,最终引发疾病的产生。
猜你喜欢
当代水产(2022年8期)2022-09-20
云南化工(2021年6期)2021-12-21
食品安全导刊(2021年20期)2021-08-30
今日农业(2021年11期)2021-08-13
农药科学与管理(2019年6期)2019-11-23
当代水产(2019年5期)2019-07-25
广东海洋大学学报(2015年3期)2015-12-22
华南农业大学学报(2015年5期)2015-12-04
遗传(2014年3期)2014-02-28
世界科学(2014年8期)2014-02-28
