
三江平原湿地小叶章功能性状对氮添加的响应
2023-10-10 09:37曹宏杰柴春荣付晓玲王继丰董海鹏刘赢男穆立蔷
草业科学 2023年9期
李 旭,曹宏杰,柴春荣,付晓玲,王继丰,董海鹏,孙 杰,刘赢男,穆立蔷
(1.东北林业大学林学院, 黑龙江 哈尔滨 150040;2.黑龙江省科学院自然与生态研究所, 黑龙江 哈尔滨 150040)
近年来,全球氮沉降显著增加,化石燃料、化肥的使用以及畜牧生产等人类活动使氮沉降量急剧增加,20 世纪90 年代中期大气氮沉降总量已达103 Tg·a-1,预计到2050 年全球活化氮的沉降量将达到195 Tg·a-1[1]。从1980 年 到2020 年,我国氮沉降通量呈现递增趋势[2],氮沉降的年平均值可达20.4 kg·hm-2[3]。氮是植物生长不可缺少的营养元素,是植物细胞内多种重要化合物的成分,例如核酸、叶绿素、光合酶等[4]。不同植物吸收不同形态氮素作为其主要氮源的能力存在差异[5]。
植物功能性状是指对植物生长、繁殖和存活等有一定影响的植物性状[6],反映植物对环境变化的适应策略[7]。湿地植物可通过改变形态特征和生物量分配模式等方式,提高植物获取限制性资源的能力或降低对该资源的需求,从而适应环境变化[8]。在湿地生态系统中,氮是影响湿地植物生长和繁殖的关键因素[9-11]。氮对植物的株高有促进作用[12-13]。陈慧敏等[14]在三江平原沼泽湿地的研究发现,施加硝酸铵6 g·(m2·a)-1显著增加禾本科植物的株高。张耀鸿等[15]研究表明,氮沉降还会通过提高叶面积指数、叶长和叶宽等功能性状来促进盐沼湿地互花米草(Spartina alterniflora)的生长。Yu 等[16]对狭叶甜茅(Glyceria spiculosa)以及Frevola 等[17]对芦苇(Phragmites australis)的研究表明,在氮限制的湿地生态系统中,一定量的氮沉降能显著促进湿地植物生物量的积累,而胡星云等[18]对碱蓬(Suaeda glauca)和张骞等[19]对海三棱藨草(Bolboschoenoplectus mariqueter)的研究表明高氮沉降量会抑制生物量的积累。水稻农田常施用尿素[20],三江平原湿地周围多有稻田,湿地承接农田退水后营养含量骤增,影响湿地植物生长[21]。我国自20 世纪70 年代以来,氮沉降中 NH4+-N/NO3--N 已由5 降到2,氮素沉降形态发生了变化[22]。不同形态氮素对植物的影响存在差异[23]。目前关于氮对湿地植物功能性状影响的研究多聚焦于氮沉降量的改变,以及单一形态的氮添加(尿素或硝酸铵),关于湿地植物功能性状对氮素形态响应的研究较少。因此,综合探讨湿地植物功能性状对不同氮素沉降量的响应规律和对不同形态氮素的偏好具有重要的研究意义。
三江平原位于黑龙江省东部,是我国淡水沼泽湿地分布面积最大的地区之一[24],也是对全球变化较为敏感的地区之一[25]。小叶章(Deyeuxia angustifolia)为禾本科(Gramineae)野青茅属(Deyeuxia)多年生根茎型草本植物,适应范围较广,在三江平原分布最为集中,是三江平原湿地重要的建群种和优势种,是维持三江平原湿地生态系统稳定的重要物种。本研究选取小叶章为材料,研究氮素形态和氮添加水平对小叶章茎、叶生长特征和生物量分配的影响,拟探究小叶章对氮添加水平和氮素形态的适应性及响应规律,为预测大气氮沉降通量增加及氮素形态变化后湿地植被的种群变化提供科学依据。
1 材料与方法
1.1 研究区概况
本研究地点选择在洪河国家级自然保护区黑龙江省科学院自然与生态研究所三江平原湿地生态定位研究站。洪河国家级自然保护区位于黑龙江省三江平原的东北部(47°42′~47°52′ N,133°34′38″~133°46′29″ E)。研究区为温带季风气候,年均温1.9 ℃,最冷月平均气温-23.4 ℃,最热月平均气温22.4 ℃,年均降水量为585 mm,降水多集中在7 月-9 月,年均蒸发量为1 166 mm。土壤类型主要为草甸土、白浆土和沼泽土。草地类型主要为草甸和沼泽,优势植物有小叶章、狭叶甜茅、毛果苔草(Carex lasiocarpa)、漂筏苔草(C.pseudocuraica)[26]。
1.2 试验设计
在洪河国家级自然保护区内选择群落组成、结构、密度相对一致的小叶章沼泽化草甸开展野外原位控制试验,设置3 种氮添加水平[低氮:N1,4 g·(m2·a)-1;中氮:N2,8 g·(m2·a)-1;高氮:N3,12 g·(m2·a)-1],3 种氮素形态[硝态氮,KNO3;铵态氮,NH4Cl;有机态氮,CO (NH2)2],以及1 个对照(CK,不添加氮素),其中4 g·(m2·a)-1为当地农田氮施入量,其余添加水平为此基础上的倍增。采用单因素控制试验,共计10 个试验处理,每个处理3 个重复,共计30 个试验小区,每个小区面积5 m × 5 m,相邻小区间隔3 m。为防止氮素流失对周围环境的影响,试验小区使用高度为0.6 m 不锈钢板进行围挡,地下埋深0.4 m,地上部分保留0.2 m。自2017 年开始,每年5 月中旬和6 月中旬分2 次施肥,将肥料融入到25 L水中,利用喷雾器均匀喷洒在植物叶面,再喷入25 L无肥清水,对照处理直接喷入50 L 清水。
1.3 取样和测定方法
2021 年8 月在小叶章地上生物量达到最大时,随机采集各小区中健康挺拔、无病虫害的小叶章有性株30 株[27],贴地面剪下植株装入有少量去离子水的封口袋中迅速带回野外试验站。采用直尺测量株高、节间长、穗长、叶长,用游标卡尺测量基径、叶宽,选取植株顶端向下数第2 片完全展开的叶片测量叶长、叶宽[28],并记录分枝数、叶片数、节间数。测量完成后,将小叶章植株拆分为叶片、茎、穗分别装入干净的纸袋中,放入75 ℃烘箱中恒温烘干至恒重后称量,记录并计算生物量参数(茎生物量、叶生物量、穗生物量、地上生物量)。
1.4 数据分析
采用双因素方差分析法(two-way ANOVA),分析氮素形态和氮添加水平及其交互作用对小叶章株高、基径、节间数、节间长、分枝数、叶长、叶宽、叶片数和各器官生物量的影响。采用单因素方差分析(one-way ANOVA)和Duncan’s 法多重比较,分析相同形态氮素下不同氮添加水平处理及相同氮添加水平下不同形态氮素处理之间小叶章功能性状和生物量的差异,统计水平为0.05。性状间的相关性采用Pearson 相关分析(双尾检验,α = 0.05)。统计分析使用SPSS 19.0,图表绘制使用Origin 2021 和Excel 2021。
2 结果
2.1 小叶章株高和茎特征对不同氮素形态与氮添加水平的响应
小叶章株高在不同添加水平的硝态氮和铵态氮处理下均显著高于CK (P< 0.05),其中硝态氮处理下,N2株高显著高于N1,与N3无显著差异(P>0.05);铵态氮处理下,N3株高显著高于N1和N2,N1和N2无显著差异。有机态氮处理下,N3株高显著高于其他水平。小叶章株高,在N1水平3 种氮素形态处理无显著差异;在N2水平有机态氮处理显著低于硝态氮和铵态氮;在N3水平铵态氮处理显著高于有机态氮和硝态氮(图1a)。
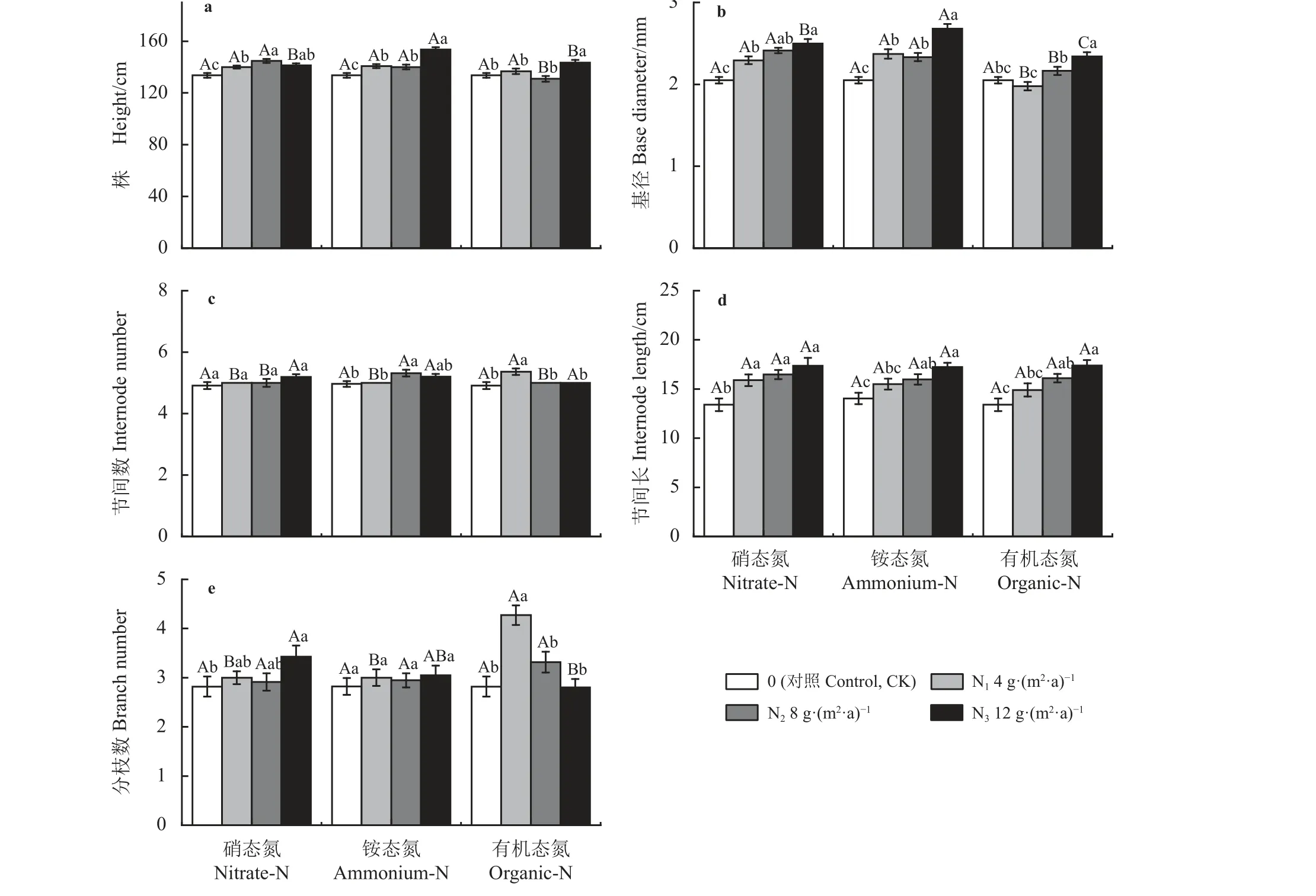
图1 氮素形态和氮添加水平对小叶章株高和茎特征的影响Figure 1 Effects of nitrogen forms and addition levels on plant height and stem characteristics of Deyeuxia angustifolia
小叶章基径在3 种氮素形态处理下均呈增加趋势,其中硝态氮处理下,N3基径显著高于N1和CK(P< 0.05),N2和N1基径显著高于CK;铵态氮处理下,N1和N2无显著差异(P> 0.05),但显著低于N3。有机态氮处理下,不同氮添加水平差异显著,但N1和N2与CK 无显著差异。小叶章基径,在N1和N2水平铵态氮和硝态氮处理显著高于有机态氮处理,但铵态氮和硝态氮无显著差异;在N3水平,铵态氮处理 > 硝态氮处理 > 有机态氮处理,且差异显著(图1b)。
小叶章节间数在硝态氮处理下随添加水平的升高逐渐增多,但CK、N1、N2、N3无显著差异(P>0.05);在铵态氮处理下,N2水平节间数显著大于N1和CK (P< 0.05),但与N3无显著差异;在有机态氮处理下,节间数在N1水平下达到最大,且显著大于其他水平。小叶章节间数,在N1水平有机态氮显著高于硝态氮和铵态氮;在N2水平铵态氮显著高于另外两种氮素形态;N3水平下3 种氮素的作用无显著性差异(图1c)。
小叶章节间长在3 种形态氮素处理下均随着氮添加水平的升高持续增长,其中硝态氮处理下,N1、N2、N3节间长显著高于CK (P< 0.05),N1、N2、N3间差异不显著;铵态氮和有机态氮处理对节间长无显著影响。相同氮添加水平下的小叶章节间长硝态氮 >铵态氮 > 有机态氮,但无显著差异(图1d)。
小叶章分枝数在硝态氮和铵态氮处理下氮添加处理高于CK (硝态氮N1水平除外),其中硝态氮N3水平显著高于其他水平(P< 0.05),铵态氮各水平间无显著差异(P> 0.05);有机态氮处理下,小叶章分枝数先增加后降低,在N1水平达到最高,且显著高于CK 和其他两个水平。小叶章节间数,在N1水平下,有机态氮显著高于硝态氮和铵态氮;在N3水平下,硝态氮处理显著高于有机态氮(图1e)。
双因素方差分析表明,氮素形态和氮添加水平及二者的交互作用均对小叶章的株高、分枝数具有显著影响(P< 0.05);氮添加水平对节间长具有显著影响;氮素形态和氮添加水平对基径影响显著,但交互作用不显著(P> 0.05);氮素形态和氮添加水平间的交互作用对节间数影响极显著(P< 0.01) (表1)。
2.2 小叶章叶特征对不同氮素形态与氮添加水平的响应
小叶章叶长随氮添加水平的增加表现出不规则变化,其中硝态氮处理在N1水平达到最大,且显著大于N2(P< 0.05);铵态氮处理在N3达到最大,且与其他水平差异显著;有机态氮处理在N2达到最大,且与CK 和N3处理差异显著。小叶章叶长,在N1、N2水平,有机态氮处理显著长于硝态氮、铵态氮,两种无机态氮处理间无显著差异(P< 0.05);在N3水平下,铵态氮对叶长的作用显著强于硝态氮、有机态氮(图2a)。
小叶章叶宽,硝态氮和有机态氮处理在N2水平达到最大;铵态氮处理在N3水平达到最大,N3叶宽显著大于CK、N1、N2(P< 0.05)。相同氮添加水平下的小叶章叶宽,除在N2水平铵态氮显著低于硝态氮、有机态氮处理,其他水平3 种氮素处理下的叶宽无显著差异 (P> 0.05) (图2b)。
小叶章叶片数随氮添加水平的增加均较CK 有不同程度的增加,其中硝态氮和铵态氮处理的叶片数在N3水平达到最大且与CK 差异显著(P<0.05);有机态氮处理下,叶片数在N1水平达到最大随后逐渐降低。小叶章叶片数,在N1水平,有机态氮处理显著高于硝态氮、铵态氮处理;在N3水平,硝态氮处理显著高于铵态氮和有机态氮(图2c)。
氮添加水平、氮素形态的极显著影响小叶章叶长(P< 0.01),交互作用的影响不显著(P> 0.05)。叶宽则仅受氮添加水平与氮素形态交互作用的极显著影响(表2)。受氮素形态和氮添加水平及二者交互作用均极显著影响小叶章的叶片数(P< 0.01)。
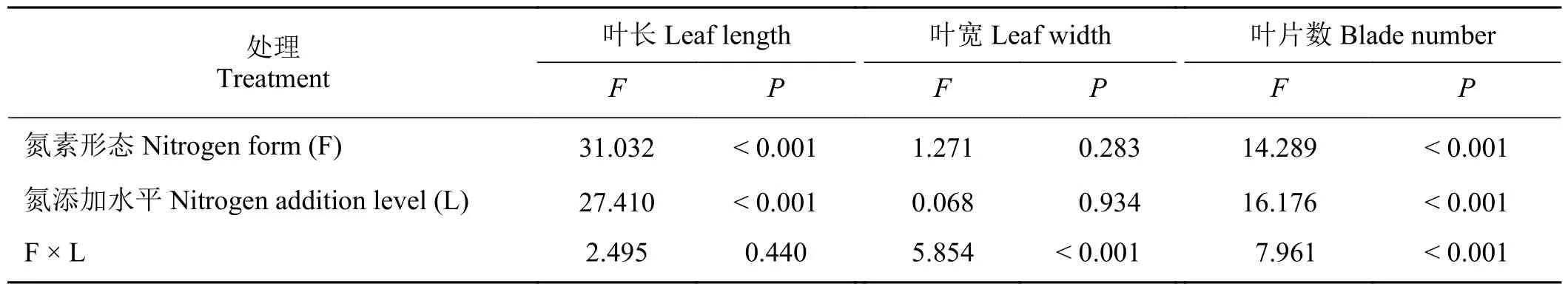
表2 不同氮素形态和氮添加水平下小叶章叶特征的双因素方差分析Table 2 Two-factor analysis of variance of leaf characteristics of Deyeuxia angustifolia under different nitrogen forms and addition levels
小叶章的功能性状间存在着不同程度的相关性。株高与节间数、节间长、基径间呈极显著正相关关系(P< 0.01),株高与分枝数则呈显著负相关关系(P< 0.05),分枝数与叶长、基径与叶长、叶宽与叶片数呈显著正相关关系(P< 0.05),叶长与叶宽呈极显著正相关关系(P< 0.01),基径与节间长、节间数呈极显著正相关关系(P< 0.01),叶片数与节间数、分枝数、基径、叶长呈极显著正相关关系(P<0.01) (表3)。
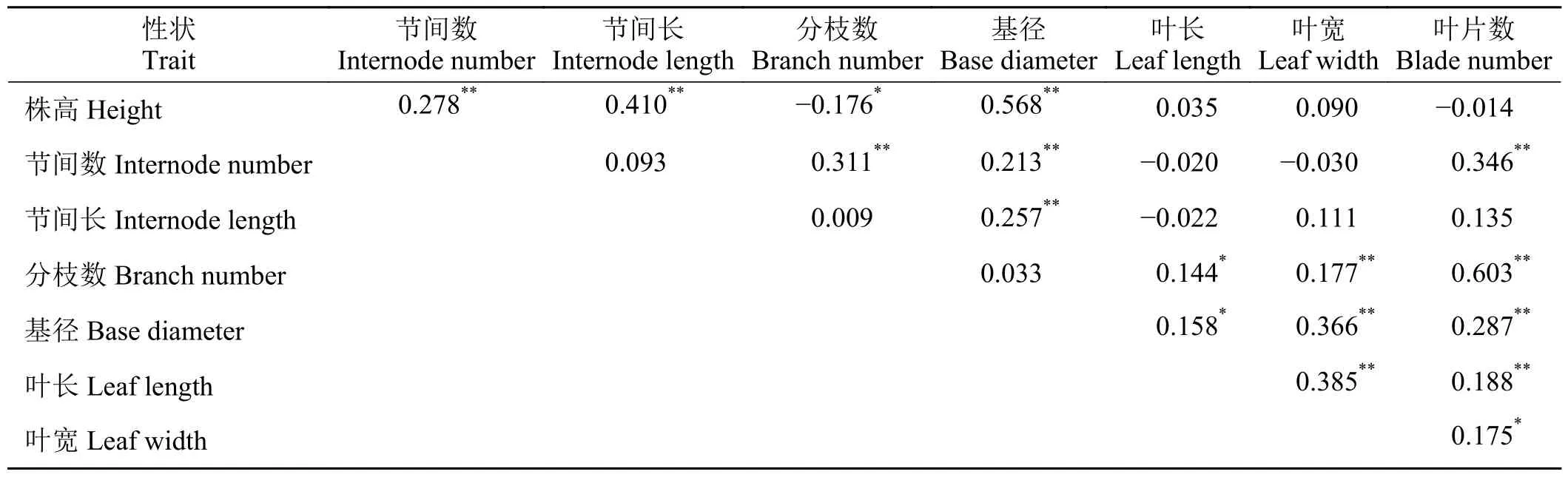
表3 小叶章功能性状间的相关性分析Table 3 Correlation analysis of functional characteristics of Deyeuxia angustifolia
2.3 小叶章各器官生物量及其分配对不同氮素形态与氮添加水平的响应
氮添加显著促进小叶章叶、茎、穗和地上生物量的增加。硝态氮与铵态氮处理下,小叶章叶、茎、穗和地上生物量均在N3水平达到最大(图3),铵态氮和硝态氮处理N3水平显著高于N1、N2和CK (P<0.05);有机态氮处理下,小叶章叶、茎和地上生物量呈先增加后降低趋势,在N1水平达到最大且与CK 差异显著。小叶章穗生物量在硝态氮与铵态氮N3水平处理下,与CK 差异显著。相同氮添加水平下,硝态氮和铵态氮处理小叶章叶生物量在N1水平与有机态氮处理差异显著,地上生物量铵态氮与有机态氮处理差异显著,茎和穗生物量无显著差异(P> 0.05);3 种氮素形态在N2水平各生物量均无显著差异茎生物量硝态氮与有机态氮除外;硝态氮与铵态氮处理小叶章叶、茎和地上生物量在N3水平显著高于有机态氮处理,铵态氮处理穗生物量显著高于硝态氮和有机态氮处理。
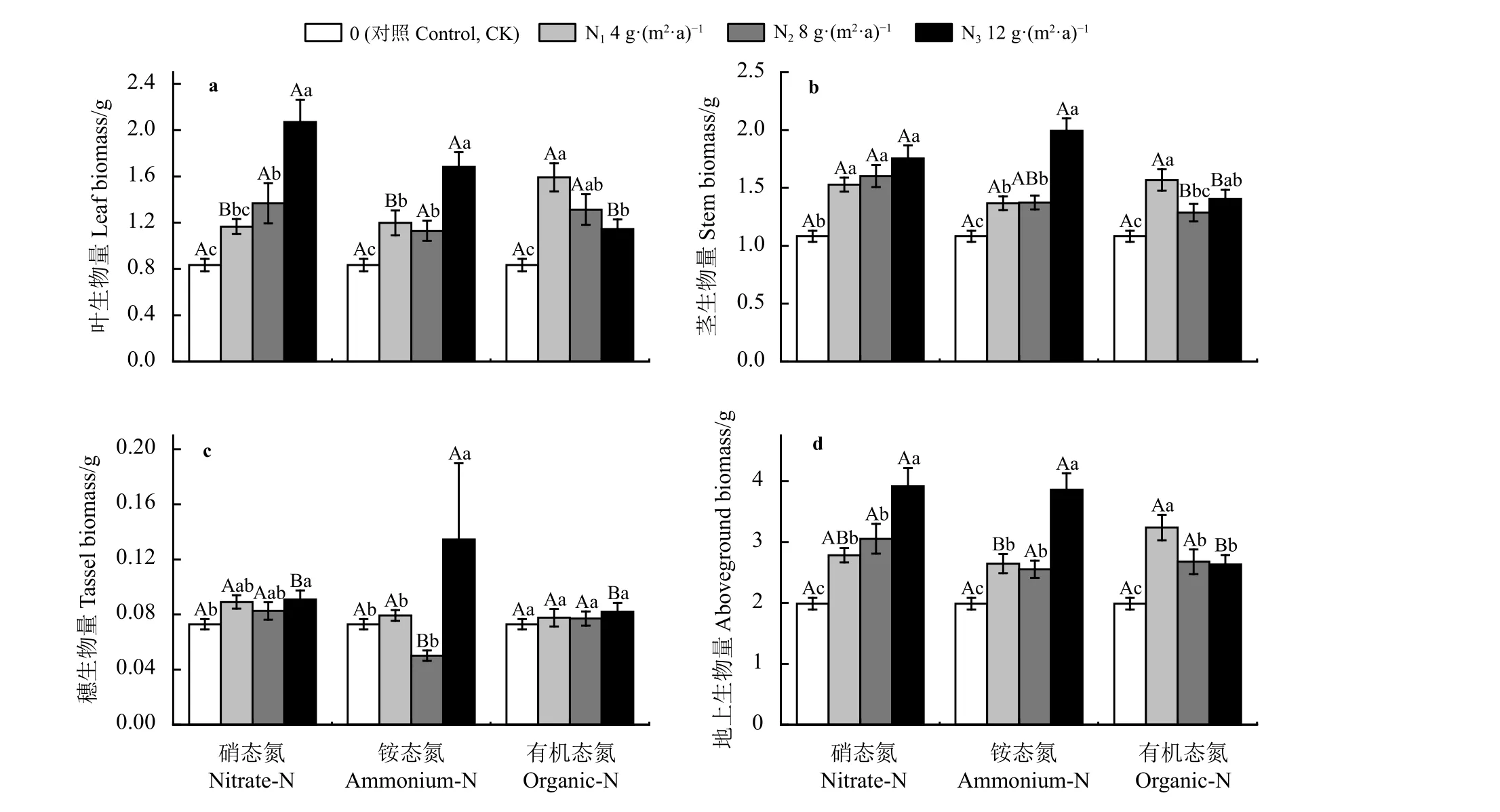
图3 氮素形态和氮添加水平对小叶章生物量的影响Figure 3 Effects of nitrogen forms and addition levels on biomass of Deyeuxia angustifolia
小叶章叶、茎、穗和地上生物量受氮添加水平及其与氮素形态的交互作用的影响显著(P< 0.05),地上生物量和茎生物量还受氮素形态的显著影响(P< 0.05) (表4)。
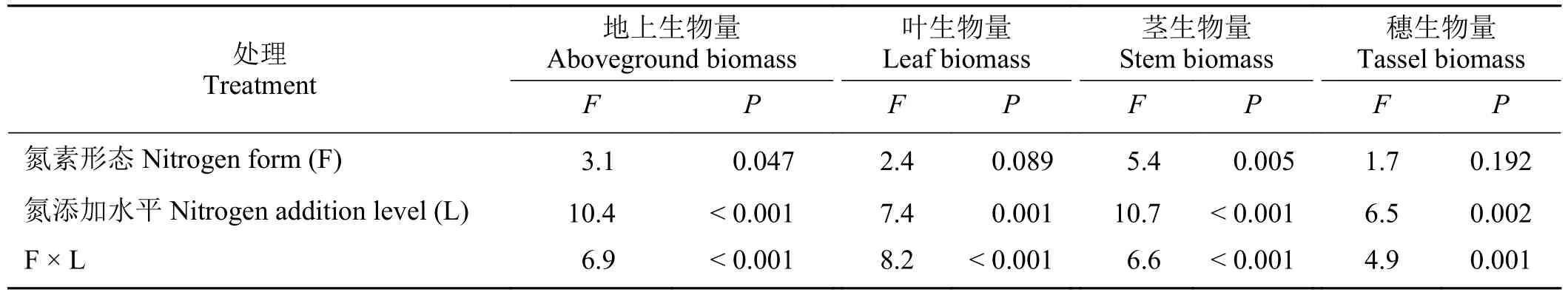
表4 不同氮素形态和氮添加水平下小叶章各器官生物量的双因素方差分析Table 4 Two-factor analysis of variance of biomass of different organs of Deyeuxia angustifolia under different nitrogen forms and addition levels
随着氮添加水平的增加,小叶章叶生物量比重逐渐增加,硝态氮处理下叶生物量在N3水平达到最大值,铵态氮和有机态氮处理下叶生物量占比在N1水平达到最大值,茎和穗生物量比重相应降低。在N1和N2水平,叶片生物量比重表现为有机态氮处理 > 铵态氮处理 > 硝态氮处理,在N3水平,叶片生物量比重表现为硝态氮处理 > 铵态氮处理 > 有机态氮处理(表5)。
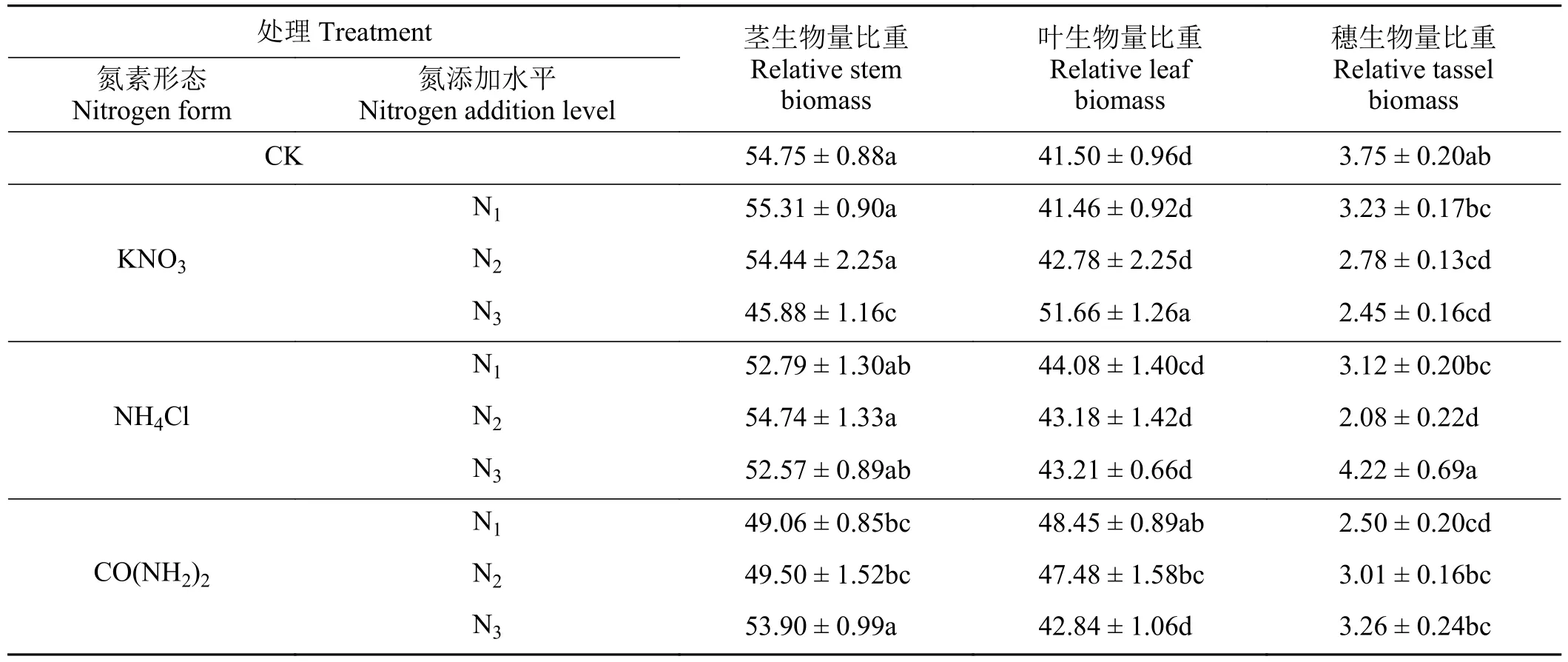
表5 不同氮素形态和氮添加水平下小叶章各器官生物量比重Table 5 Proportion of biomass in organs of Deyeuxia angustifolia under different nitrogen forms and addition levels%
3 讨论
3.1 氮添加对小叶章生长特征的影响
株高、节间长是衡量植物长势的重要性状,可以反映湿地植物竞争生长资源的能力[29]。在本研究中,3 种形态氮素处理下的小叶章株高、节间长整体随着氮添加水平的增加呈增长趋势,该结果与刘德燕等[30]和窦晶鑫等[31]研究结果一致。古勇波等[32]的研究表明,随氮添加水平增加,三江藨草(Scirpus nipponicus)株高呈递增趋势。但不同氮素形态对小叶章株高的影响程度不同,在低氮水平下3 种氮素形态对株高影响无显著差异,在中氮水平,无机态氮处理株高显著高于有机态氮,在高氮水平下铵态氮处理株高显著高于其他两种氮素形态。从节间数量的变化特征来看,不同氮素形态对节间数量的影响较小,节间数仅在中氮水平的铵态氮处理和低氮水平的有机态氮处理下显著增加,但这两个水平下小叶章株高并没有因节间数的增加而显著高于其他处理,且节间长与株高的相关性相较于节间数更强,可见施氮后小叶章株高的增加更倾向依靠节间长度的增加而非节间数的增多。
形态特征的改变是植物适应不同环境和资源水平的重要策略,叶片作为植物进行光合作用的主要器官,其性状特征如叶片数、叶长、叶宽直接影响植物的光合作用[33-34]。本研究中,氮添加增加了小叶章叶片的宽度,并在中高氮水平增加明显。不同氮素形态对叶片宽度的影响主要体现在中氮水平,在该水平有机态氮对叶宽的影响最大,其次是硝态氮,且与铵态氮差异显著。氮添加对叶片长度的影响在不同氮素形态处理下表现不一致,随硝态氮添加水平的增加,叶长先增大后减小,然后再增大,叶宽先减小后增大再减小。铵态氮处理的叶长和叶宽变化在低氮和高氮水平变化一致,并在高氮水平与对照差异显著。有机态氮处理的叶长和叶宽均表现为随氮添加水平的增加先增加后降低,在中氮水平达到最高。叶长和叶宽的变化侧面体现了叶面积的变化,经上述分析可知,硝态氮处理可能对叶面积的影响不显著,而铵态氮和有机态氮则分别在高氮水平和中氮水平显著增加了叶面积。这与杨浩等[35]对糙隐子草(Cleistogenes squarrosa)的施氮研究结果一致,在两种水分条件下,中氮水平的有机态氮均显著促进糙隐子草叶面积的增长。氮添加还增加了小叶章的分枝数和叶片数,硝态氮和铵态氮的分枝数和叶片数在高氮水平最大,有机态氮则在低氮水平最大。可见氮添加通过增加小叶章的分枝数和叶片数来促进植株生长,有机态氮在低浓度作用强度大,而无机态氮在高浓度作用强度大。该结果与张耀鸿等[15]的研究结果一致,施氮后,互花米草植株叶面积的增加主要通过叶片数的增加实现。
本研究中,硝态氮处理和铵态氮处理的小叶章节间长、分枝数在各浓度水平下均无显著差异,株高、基径、叶片数、叶长仅在高浓度水平出现显著差异,节间数和叶宽仅在中浓度水平出现显著差异,铵态氮处理下的小叶章株高、基径、节间数显著大于硝态氮,叶片数和叶宽则相反。可见,低浓度的硝态氮和铵态氮在对小叶章生长的影响上表现出了较强的相似性,中高浓度硝态氮和铵态氮对小叶章生长产生了不同的影响,其中铵态氮更侧重于影响小叶章的茎,而硝态氮则侧重于影响小叶章的叶。有机态氮在节间数、分枝数、叶片数上表现出了与两种无机态氮相反的趋势,在低氮水平达到最大值,对叶长和叶宽的影响在中氮水平达到最大值,在高氮水平对小叶章株高、基茎等的生长指标的影响效果小于两种无机态氮。这体现了植物对不同形态氮素的吸收能力和耐受范围不同[23,36-38],本研究中施氮后的土壤氮素浓度未超过小叶章的耐受范围,氮元素仍是主要限制性资源,小叶章可能会吸收各种形态的氮素满足自身生长对氮的需求。小叶章对有机态氮的耐受范围相比无机态氮小,可能是因为尿素水解会生成大量NH3,使土壤pH 升高,灼伤植物根系,使植株萎靡甚至枯死[39]。随着尿素添加量的增大,毒害作用增强,因此对小叶章生长的促进作用显著减弱。
3.2 氮添加对小叶章生物量分配的影响
已有研究表明模拟氮沉降会增加植物可利用的氮促进植株生长[40],一定量的氮沉降可以提高植物的生产力,然而过高氮量会导致土壤酸化、营养元素不平衡,反而不利于植物的正常生长[41-42]。本研究中,硝态氮和铵态氮处理下的小叶章,随着氮添加水平的增加地上生物量及营养和繁殖器官生物量(茎、叶生物量)均在N3水平达到峰值,说明该氮浓度水平未超出小叶章生长需求,这与小叶章在输入0~24 g·(m2·a)-1硝铵态氮和0~5 g·(m2·a)-1铵态氮的处理下地上生物量增长趋势的研究结果一致[30-31]。与无机态氮添加的趋势相反,随着有机态氮添加水平的增加,小叶章的地上生物量及营养器官生物量先增加后减少,在N1处理达到最大后出现显著下降,表明小叶章对不同氮素形态的耐受能力不同。赵连军等[43]在黄河三角洲高潮滩的研究中施用有机态氮,研究表明芦苇对施氮量有一定的耐受范围,添加50 g·(m2·a)-1有机态氮对芦苇生长繁殖无显著影响。潘艳文等[44]2017 年的研究中施用浓度范围为0~90 g·(m2·a)-1的硝铵态氮,随施氮量升高芦苇幼苗的生物量持续增加。
施氮能显著改变禾本科植物的生物量分配格局[45]。本研究中,小叶章叶生物量占比在施氮后升高,茎、穗生物量占比相应降低,这一结果是由于施氮增加了小叶章茎分枝数量和叶片数量。与对照相比,氮添加后更多的生物量投入到叶片这一碳同化器官,提高了植株的竞争能力[46]。在付晓玲等[47]的研究中,小叶章表现出喜硝性,各器官短期内(72 h)偏好硝态氮的优先顺序为叶 > 茎 > 根。从不同氮素形态比较来看,高浓度硝态氮处理的叶生物量占比最大,中低浓度有机态氮处理的叶生物量占比次之,铵态氮处理的叶生物量变化最小。这可能还是由于小叶章对不同氮素形态喜好及吸收程度决定的。比如不同形态氮素均可提升宽叶绿绒蒿(Meconopsis rudis)幼苗的光合作用能力,但提升效果上,铵态氮明显优于硝态氮[48]。
4 结论
施氮能促进三江平原小叶章的生长。小叶章对不同氮素形态的耐受范围存在差异。低氮水平下,两种无机态氮对小叶章生长的影响表现出较强的相似性,中氮、高氮水平下,铵态氮更侧重于影响小叶章的株高、基径、茎生物量,而硝态氮更侧重于增加小叶章的叶片数。有机态氮在节间数、分枝数、叶片数表现出了与两种无机态氮相反的趋势,在低浓度作用强度大,而无机态氮在高浓度作用强度大。
施氮对小叶章地上生物量积累有明显的促进作用,显著增加了小叶章茎分枝数量和叶片数量,导致叶生物量比重增大,茎和穗生物量比重降低。
猜你喜欢
桂林理工大学学报(2021年3期)2021-12-06
华北水利水电大学学报(自然科学版)(2020年6期)2021-01-29
农业环境科学学报(2020年10期)2020-11-11
水科学与工程技术(2016年2期)2016-07-10
环境污染与防治(2016年12期)2016-03-13
湖泊科学(2015年1期)2015-05-06
中国酿造(2014年9期)2014-03-11
植物营养与肥料学报(2011年5期)2011-11-06
植物营养与肥料学报(2011年4期)2011-10-26
植物营养与肥料学报(2010年3期)2010-11-16
