
外源电子与天然矿物对产甲烷菌生长代谢的影响
2023-10-09 01:03王一杰贾铭宇
化学与生物工程 2023年9期
郑 懿,王一杰,马 蕾,杨 悦,贾铭宇,赵 敏
(东北林业大学生命科学学院,黑龙江 哈尔滨 150040)
随着社会经济的快速发展,人们对能源的需求越来越大。为减轻对传统化石能源的依赖,新能源的开发和利用越来越受到重视。甲烷作为一种清洁、可再生新能源,具有不可替代的作用。产甲烷菌是一类能将无机或有机化合物通过厌氧发酵最终转化为甲烷和CO2的古细菌,主要存在于沼泽、湖泊、海洋沉积物和动物瘤胃液等自然生态系统中,也存在于废水、堆肥和消化污泥等非自然生态系统中,是全球碳循环的重要组成部分[1-2]。其产甲烷特性是可被利用的有效生物能源策略,对解决当前环境问题和能源危机具有重大意义[3-4]。产甲烷菌生长的常见底物有H2/CO2、乙酸、甲酸和甲醇等,但在现有培养条件下底物转化效率极低,产甲烷菌的生长速率非常缓慢,极大制约了产甲烷菌的实际工业化应用。近年来,研究人员开发了提升产甲烷菌产甲烷速率的有效方法,如微电流驱动法[5]、导电聚苯胺法[6]、活性炭法[7]及添加导电纳米材料或改性材料法[8-9]等。上述方法的基本原理是通过提高培养体系的胞内外电子传递效率,进而促进产甲烷菌生长代谢。不难发现,电子传递是微生物存活和代谢的基础,是产甲烷过程的重要条件。外源电子的介入可以有效加快产甲烷速率,为改良现有产甲烷菌培养方案提供了有效途径[10]。基于此,作者对产甲烷菌对外源电子的利用能力进行综述,探讨天然矿物调控产甲烷菌生长代谢的方式,以期为产甲烷菌的应用奠定理论基础。
1 产甲烷菌对外源电子的利用能力
1.1 互营细菌提供外源电子
微生物“互营”是互惠共生的一种互作方式。在缺乏产甲烷菌可利用底物的厌氧环境中,厌氧互营细菌与产甲烷菌的互营代谢合作关系更为紧密。在该过程中,通常需要互营细菌降解脂肪酸、芳香族化合物、醇类及含氮有机物等物质,为产甲烷菌提供氢、甲酸、乙酸等底物,形成产甲烷互营代谢[11]。互营细菌与产甲烷菌的生长代谢均通过种间电子传递载体相互转化而偶联。
除了这种有机质厌氧代谢的经典偶联方式,种间电子传递更是自然界中广泛存在的现象,在互营产甲烷过程中起到关键作用[12-13]。即细菌利用细胞外不溶性的电子受体完成氧化还原反应,进行呼吸电子传递,并获得能量。目前普遍认为这种机制是通过细菌外膜C型细胞色素、细胞附属物或胞外电子穿梭体等方式介导[14-15]。地杆菌与产甲烷菌可通过电子传递形成互养共生关系[16](图1)。地杆菌通过氧化有机物产生电子,释放到胞外并传递给产甲烷菌,进而将底物还原为甲烷。Rotaru等[17]通过研究Methanosaetaharundinacea和Geobactermetallireducens共培养体系发现,若将地杆菌中编码导电菌毛的pliA基因敲除,该体系则无法利用乙醇产甲烷,说明地杆菌通过导电菌毛将电子传递给产甲烷菌,使其完成生长代谢。随后,Holmes等[18]研究发现,稻田土壤中的甲烷鬃毛菌和甲烷八叠球菌只能通过微生物种间电子传递的方式将有机物转化为甲烷。Rotaru等[19]还发现在海底沉积物中也存在该现象。这些研究表明,微生物种间电子传递过程对自然界中产甲烷菌生长代谢起到了极为重要的作用,在长期生命进化过程中可能普遍存在。

图1 地杆菌与产甲烷菌间依赖电子传递的互养共生关系Fig.1 Electron-transfer dependent symbiotic relationship between Geobacter and methanogen
1.2 电极输入外源电子
外界的电压和电流能够调控微生物体系中的菌群结构,影响电子传递和一系列氧化还原反应[20]。Ding等[21]研究发现,给予厌氧消化体系合适的电压会影响微生物活性,甲烷产量增幅超过4倍。Alqahtani等[22]研究发现,在富集碳固定微生物群落时引入外加电压,可以提高体系甲烷产量,这可能与微生物群落变化、乙酸产量提高有关。刘洪周等[23]研究发现,外源添加0.6 V的电势差时,系统甲烷产量最高可提升77%;结合代谢通量分析发现,所产甲烷仍主要来源于体系中的乙酸,但用于产甲烷的氢气通量受电压影响显著,随电压增大明显升高。电化学技术是研究微生物亲电或产电作用的直接手段,在外界电流输入的条件下观察微生物的代谢作用,能够直观体现微生物利用外源电子的能力。由此可见,产甲烷菌一般可以利用外源电子作为还原力提供自身产能,进而促进生长代谢,提升甲烷产量。
1.3 元素价电子
元素价电子能量是自然界中一种重要能量形式,是自然界中变价元素的核外电子在受到氧化还原反应等化学作用时所体现的一种化学能。变价元素(主要有铁、铬和锰等元素[24])通过与微生物胞外聚合物、环境介体或细胞表面膜蛋白发生氧化还原反应,经电子传递路径向菌株输送价电子,这是微生物利用价电子的主要来源。Karri等[25]研究发现,零价铁可在底物污泥中作为微生物产甲烷的缓释电子供体,甲烷生成速率与零价铁的比表面积呈正相关。Hu等[26]研究发现,在Trichoccus参与下低浓度Cr(Ⅵ)可促进乙酸营养型产甲烷菌产甲烷。Fu等[27]在乙酸营养型产甲烷菌培养体系中添加纳米Fe3O4,发现纳米Fe3O4可促进乙酸营养型产甲烷菌产甲烷,结合光谱分析发现,产甲烷菌细胞膜中纳米Fe3O4作为电子穿梭器提高了细胞内电子传递效率,进而使甲烷产量提高。Tang等[28]将水铁矿二次矿化产生的磁铁矿作为培养物,研究其对产甲烷菌生长代谢的影响,发现加入磁铁矿后产甲烷菌对乙酸盐转化为甲烷的效率提高了30.2%。可见元素价电子是产甲烷菌利用外源电子的主要来源,能够显著提高多种培养条件下的菌株生长量和甲烷产量。
2 天然矿物调控产甲烷菌生长代谢的方式
微生物是自然界物质循环的重要组成部分,矿物与微生物在能量和物质之间一直有着复杂的作用关系,这推进了微生物的代谢进化,同时也促进了地球生物化学演化的多个过程,如碳、氮元素循环等[29]。
2.1 提高电子传递效率
天然矿物具有缩短产甲烷菌代谢滞后期、提高甲烷产量和产甲烷速率的作用,且与矿物加量和培养周期呈正相关[30]。此外,一些天然矿物的加入还可以促进具有种间电子传递功能的微生物与产甲烷菌互营生长,间接提高产甲烷菌的电子传递效率,提升产甲烷效率。Kato等[31]研究发现,含铁半导体矿物可以加速厌氧体系中甲烷的生成。Xu等[32]在厌氧反应器中加入Fe2O3纳米颗粒,通过促进铁还原细菌与产甲烷菌的种间直接电子传递,使甲烷产量提高了15.4%。高雪濛等[33]在反应器中加入底物并接种厌氧污泥,比较了在无Fe2O3、48 h后加入Fe2O3、开始时加入Fe2O3等3种条件下构建系统的产甲烷能力,结果表明,Fe2O3可解除酸抑制,加快厌氧菌的产甲烷速率。Ambuchi等[34]利用厌氧菌降解甜菜制糖工业废水,并探究了赤铁矿对甲烷产量的影响,结果表明,在赤铁矿存在下,甲烷产量提高了35%。Wang等[35]研究发现,培养体系中含铁矿物颗粒的介入可以显著刺激微生物合成蛋白质和腐殖酸,提升电子传递能力,进而提高产甲烷效率(图2)。这表明,矿物颗粒还可以通过促进胞内物质合成,增加细胞色素,进而刺激微生物进行更高效的电子传递。随后研究人员通过深入研究,进一步揭示了磁铁矿主要是通过替代微生物导电物质或诱导微生物导电物质的合成和表达来发挥作用;赤铁矿则可以通过铁离子作用促进细胞聚集体的形成和稳定,有利于微生物进行种间电子或电子载体的传递[36-37]。
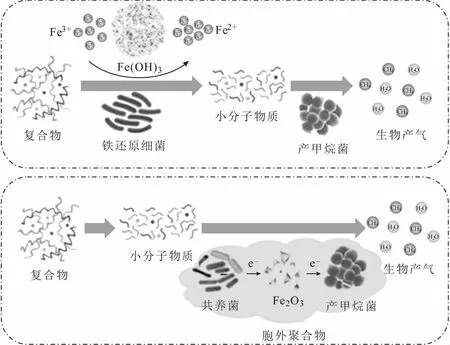
图2 Fe(OH)3和Fe2O3促进产甲烷菌代谢产甲烷的作用机制Fig.2 Mechanism of methane production by methanogens that promoted by Fe(OH)3 or Fe2O3
除了含铁矿物具有相应功能外,Ma等[38]通过在湿地系统中添加TiO2NPs,发现电活性菌和产甲烷菌同时得到富集,系统产甲烷效率可提高48.6%,表明TiO2NPs可以在自然环境中作为电活性菌进行电子传递的导体,促进种间电子传递,进而提高厌氧产甲烷能力。此外,Cr2O3和ZnO等矿物材料也对厌氧产甲烷过程起到促进作用,可见多种矿物均能促进产甲烷菌的电子传递及生长代谢。
2.2 参与氧化还原反应
在厌氧反应体系中,重金属离子、体系pH值及气体分压等条件会影响微生物生长代谢,而天然矿物材料会通过参与氧化还原反应调控环境因子,使其适宜产甲烷过程。Zhang等[39]研究发现,纳米半导体矿物材料可作为还原剂去除反应系统中的重金属离子,有利于微生物进行正常的生长代谢,进而提高甲烷产量。Lombi等[40]研究发现,在厌氧消化体系中引入的ZnO可与H2S反应,形成ZnS沉淀,减少H2S产生,降低其对厌氧消化微生物产甲烷的影响。Uchiyama等[41]研究发现,在N2和CO2环境下,零价铁可作为唯一电子供体,产甲烷菌可以通过氧化作用在其表面直接获得电子而生长。Wei等[42]研究发现,在培养体系中引入的含铁矿物极可能被微生物直接利用,矿物中的铁离子能够参与代谢关键酶的合成,加快氧化还原反应速率,进而提高甲烷产量。这种基于氧化还原反应的化学作用,可能是自然环境中矿物与微生物共存的基础。矿物介入微生物生长代谢,同时在微生物作用下完成组分演化,微生物从中获取能量,进而提高代谢产物产量。
2.3 提供光生电子
地球上矿产资源丰富,矿物种类多样,其中具有半导体性质的天然矿物能够被太阳光激发完成光电转化,即当光子能量高于矿物禁带宽度时,价带电子被激发并跃迁至导带,产生空穴和光生电子。2012年,Lu等[43]首次发现微生物能够通过闪锌矿等天然矿物光生电子促进其生长代谢,形成非光合微生物的“矿物-微生物”光合作用。此后,该研究团队利用多学科交叉研究手段,揭示了地表矿物与太阳光响应之间的关系(图3)[44],并认为这种光电子能量是继元素价电子能量与太阳光子能量后的第三种地表能量形式,意义重大[45]。目前,研究已发现多种微生物具有利用光生电子的能力,并参与多种代谢途径,这为“矿物-微生物”交互作用研究提供了新方向[46-47]。

图3 地表天然矿物膜对太阳光的响应Fig.3 Response of earth mineral membrane to sunlight
Anjum等[48]研究发现,半导体矿物纳米颗粒经可见光激发后,可使甲烷产量提高1.6倍。Ye等[49]将CdS矿物与Methanosarcinabarkeri共培养,发现光生电子能够通过氢化酶和细胞色素传递,推进菌体固定CO2产甲烷,使菌体胞内产甲烷基因表达量显著上调,提升了151.4%,产甲烷速率为0.19 μmol·h-1,体系的量子产率达0.34%。该团队还通过将双活性位点(NiCu合金)纳入CdS半导体和产甲烷菌之间的界面(图4),调控界面产氢效应,进一步提高产甲烷性能,使体系中近100%的H2被细菌利用,驱动其转化CO2产甲烷,量子产率高达12.41%[50]。由此证明,矿物光生电子对产甲烷菌生长代谢的促进作用十分明显,极有可能成为改良产甲烷菌培养方式的新方向。

图4 太阳光驱动的Methanosarcina barkeri-NiCu@CdS微生物光电化学产甲烷系统Fig.4 Solar-driven methanogenesis system with Methanosarcina barkeri-NiCu@CdS by microbial photochemistry
3 展望
产甲烷菌的生长代谢对地球环境碳循环具有十分重要的作用。产甲烷菌可以利用多种来源的外源电子,通过电子传递作用进入胞内,加快产甲烷菌代谢速率,提升甲烷产量。与水电、火电及核电等电力能源相比,通过半导体矿物将太阳光转化为电能具有成本低、效率高的特点。在胞内还原力提高的条件下,利用产甲烷菌将环境小分子碳化合物进一步转化为更高效的生物能,可以解决目前能源领域的关键问题。但天然矿物光生电子对产甲烷菌的作用机理仍有待深入研究,将矿物光生电子与现有产甲烷菌培养技术整合的高效策略极可能成为未来的研究热点,在生物燃料合成领域具有极为广阔的应用前景。
猜你喜欢
舰船科学技术(2022年11期)2022-07-15
西藏农业科技(2019年3期)2019-11-04
智富时代(2019年5期)2019-07-05
智富时代(2019年5期)2019-07-05
现代园艺(2018年3期)2018-02-10
黑龙江八一农垦大学学报(2017年5期)2017-10-31
上海农业学报(2017年3期)2017-04-10
湖南农业科学(2017年7期)2017-03-04
河北建筑工程学院学报(2015年2期)2015-04-29
河北建筑工程学院学报(2015年4期)2015-03-24
